
Effects of Light on Growth and Metabolism of Rhodococcus erythropolis

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Culture Conditions
2.2. Cultivation of R. erythropolis under Light of Different Wavelenths
2.3. Cultivation of R. erythropolis under LED Light for Proteomic Analysis
2.4. Growth Analysis
2.5. Pigment Extraction
2.6. Fatty Acid Profile
2.7. Proteomics
2.7.1. Protein Extraction and Precipitation
2.7.2. Protein Quantification and SDS-PAGE
2.7.3. In-Gel Digestion of Protein Samples and LC-MS/MS Analysis
2.7.4. Bioinformatics Analysis
3. Results
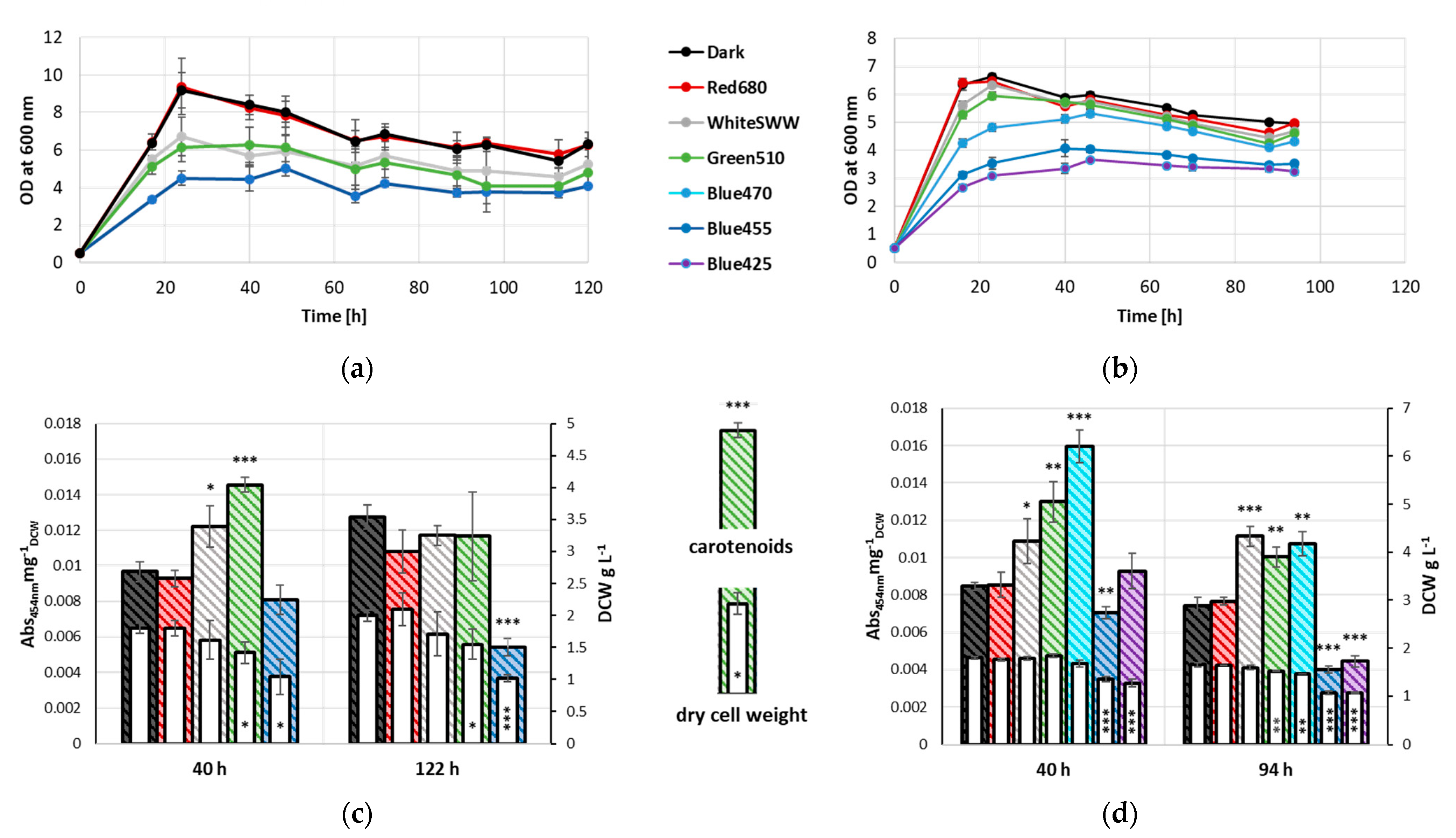
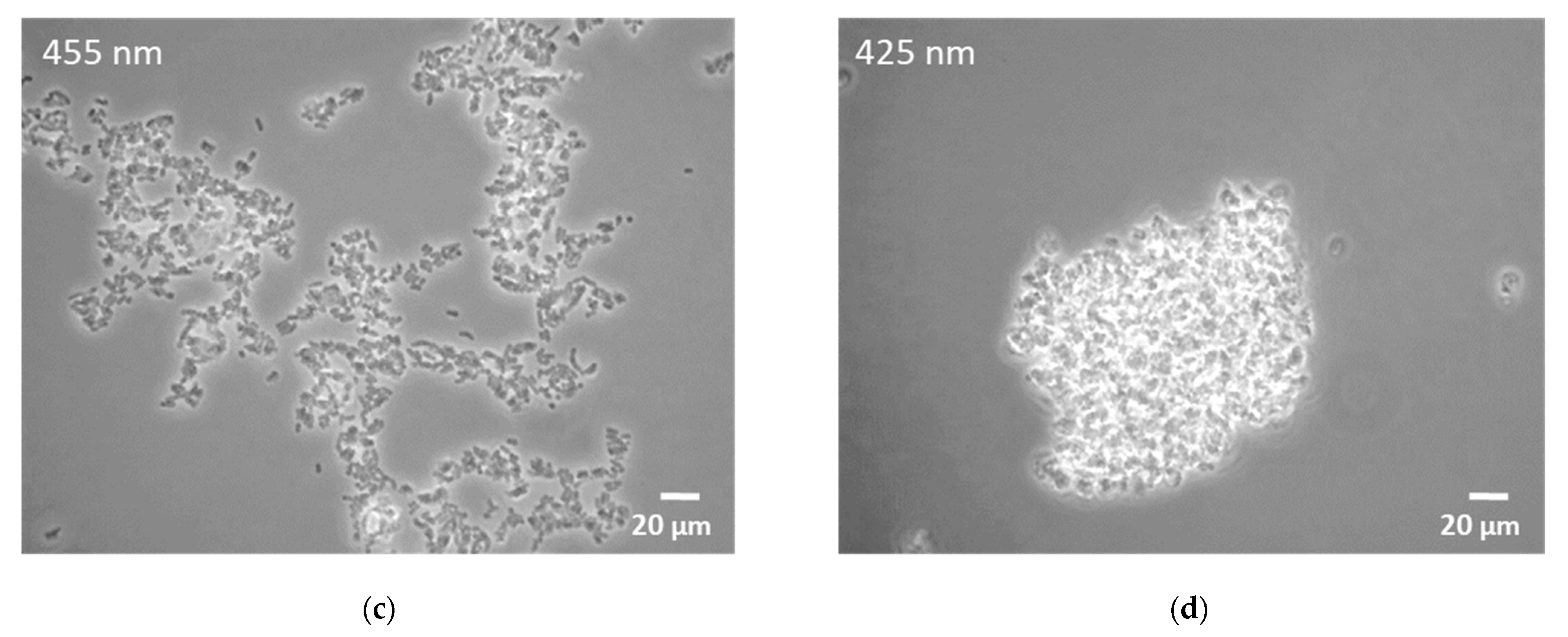
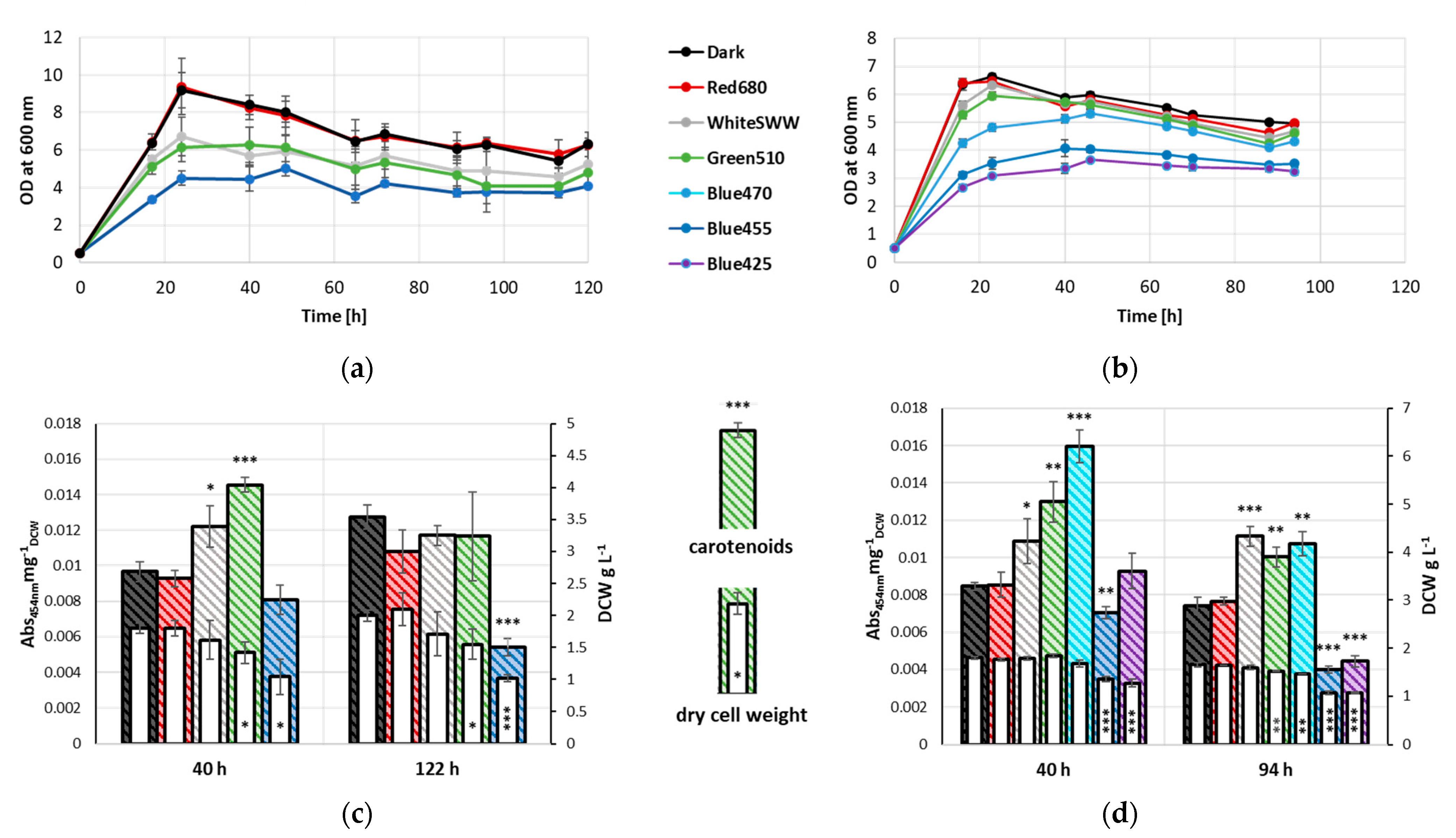
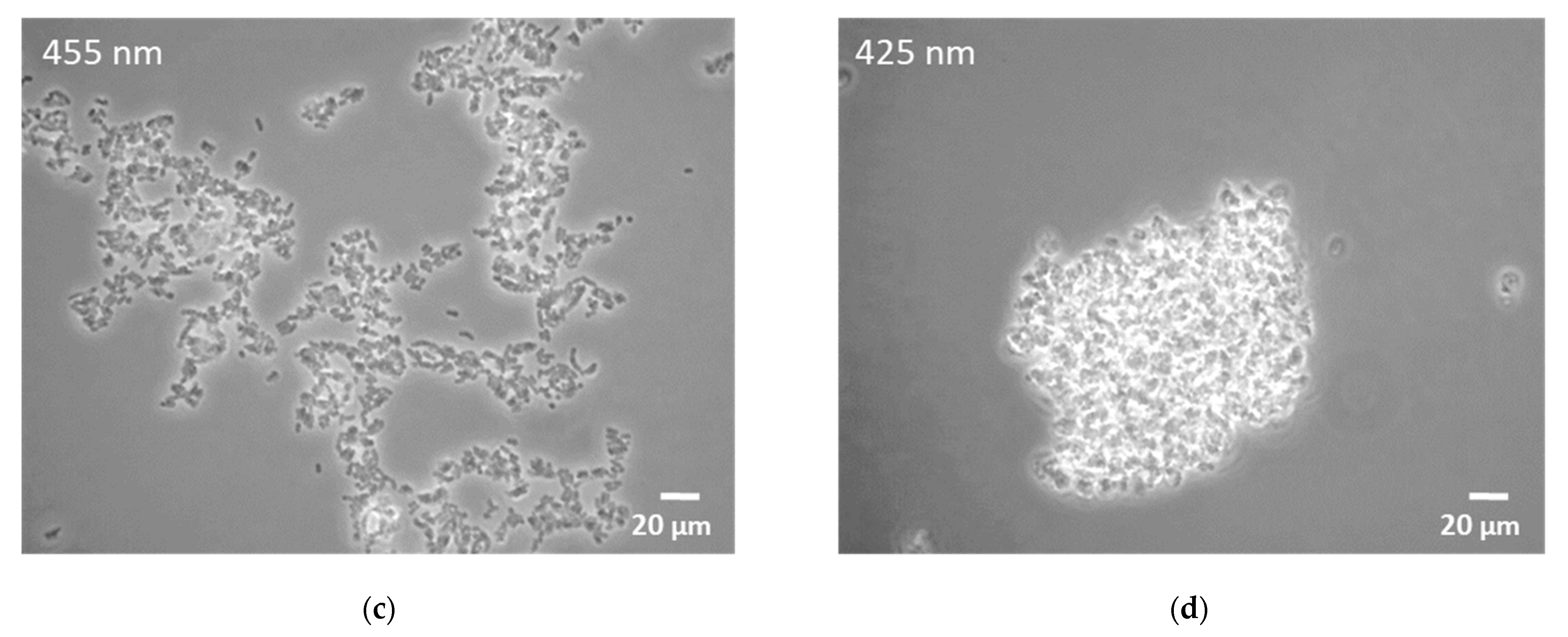
3.1. Influence of Light Quality on Growth Characteristics and Carotenoid Accumulation
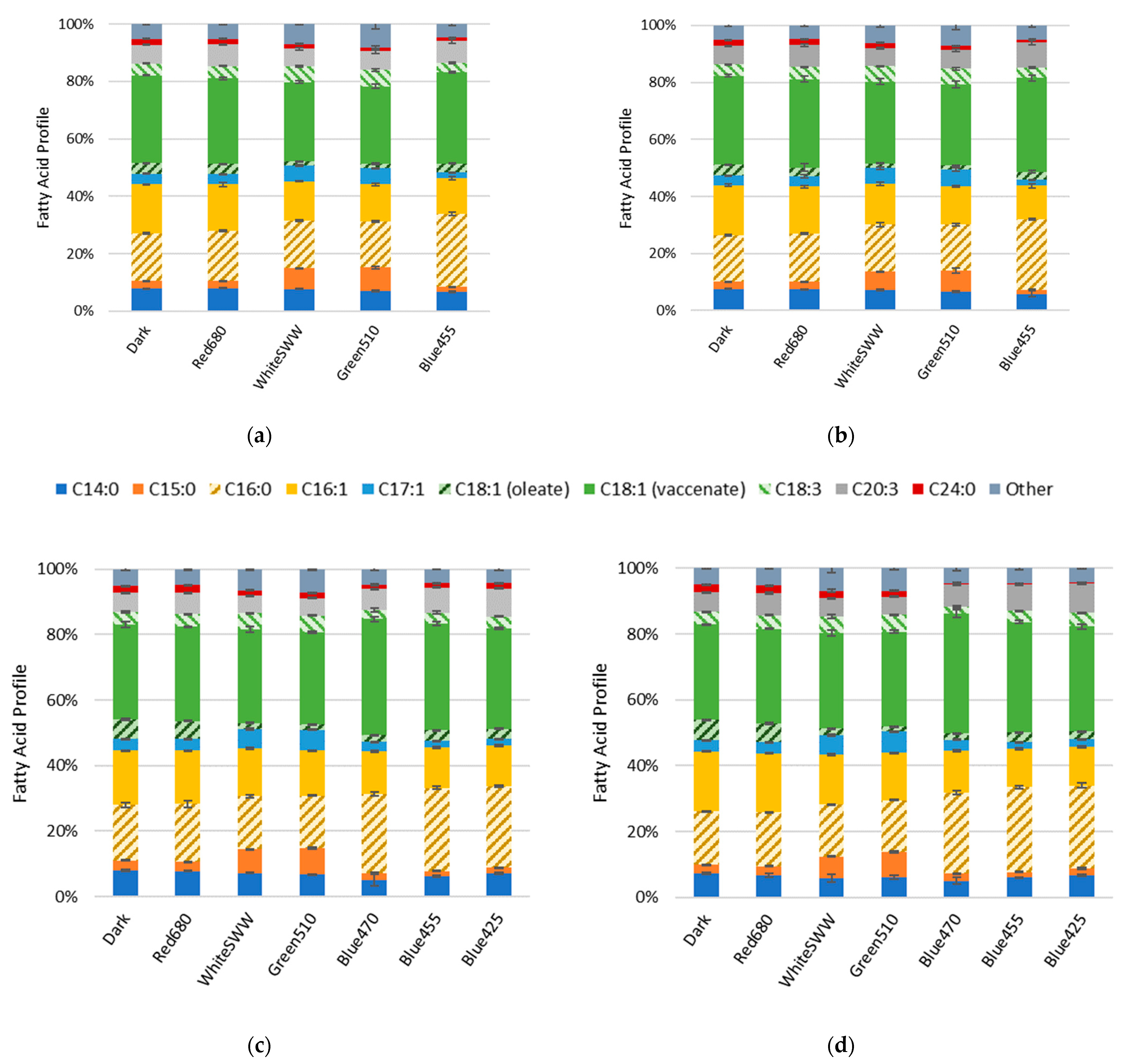
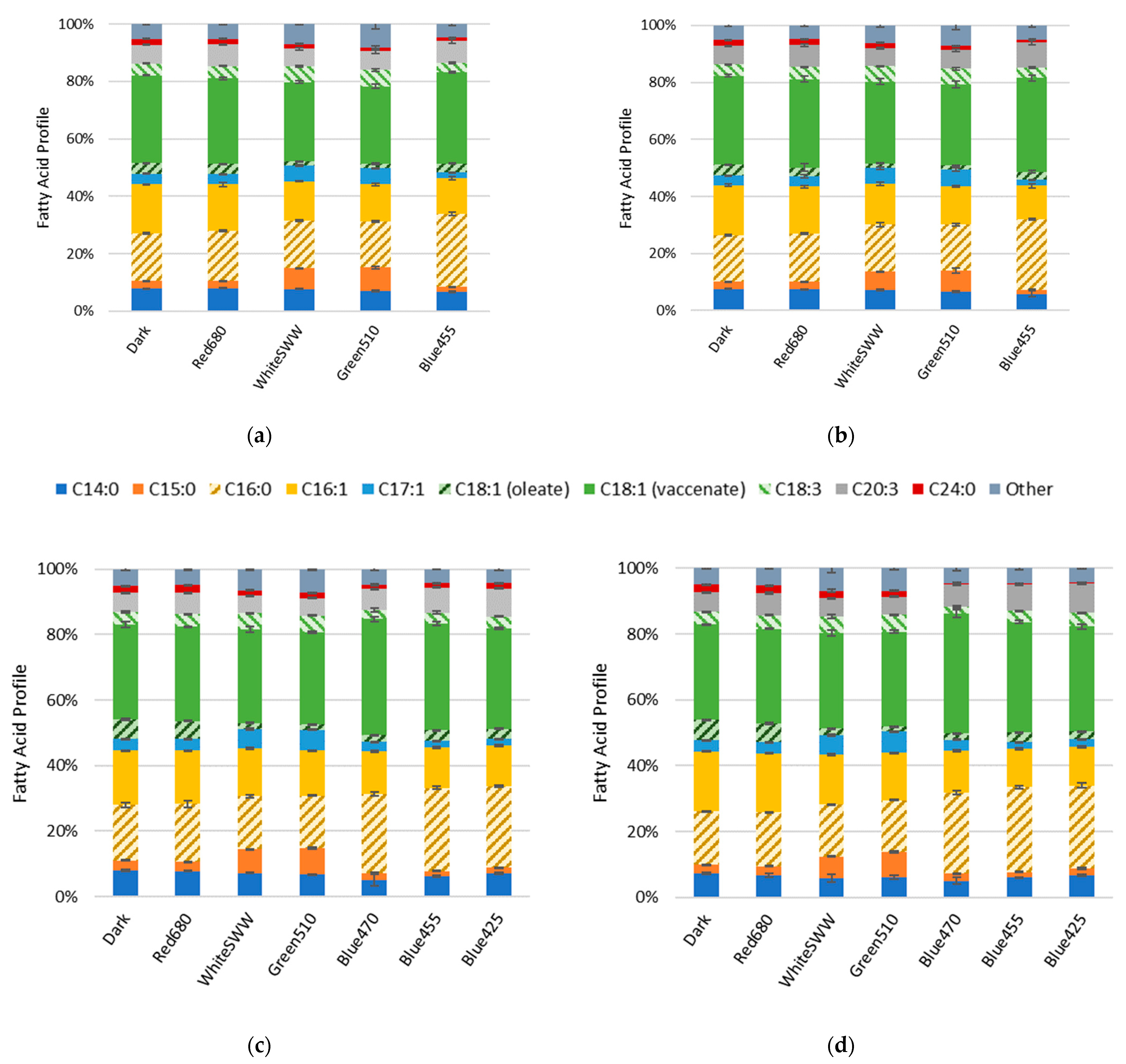
3.2. Influence of Light Quality on Fatty Acid Composition
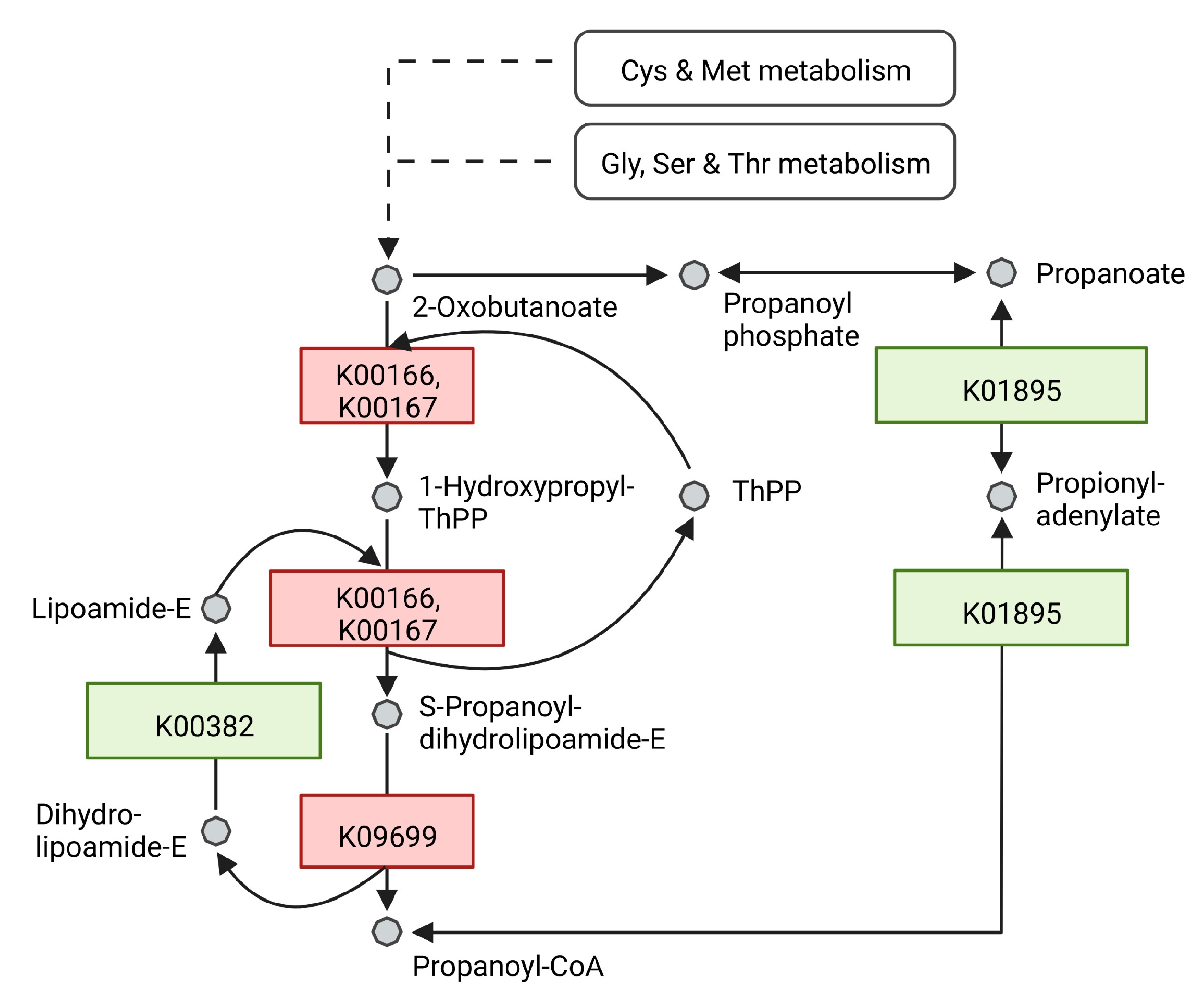
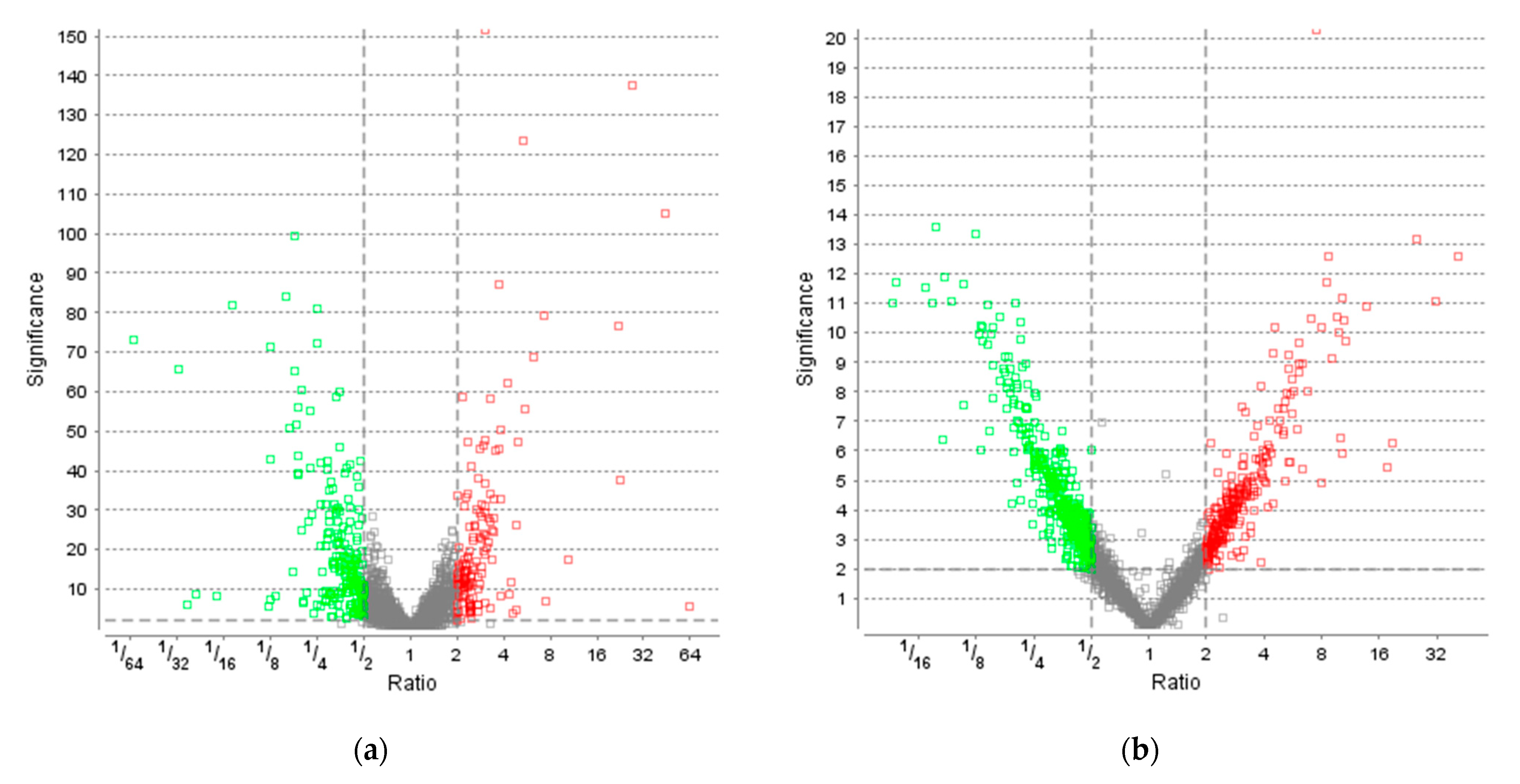
3.3. Effect of Light on Protein Levels
4. Discussion
4.1. Effects of Light Illumination on Growth
4.2. Changes in Carotenoid Content
4.3. Transcription Factors
4.4. Catalases and Peroxidases
4.5. Other Proteins Related to Stress
4.6. Influence of Light on Fatty Acid Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bell, K.S.; Philp, J.C.; Aw, D.W.; Christofi, N. The genus Rhodococcus. J. Appl. Microbiol. 1998, 85, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Gűrtler, V.; Seviour, R.J. Systematics of Members of the Genus Rhodococcus (Zopf 1891) Emend Goodfellow et al. 1998. In Biology of Rhodococcus; Microbiology Monographs; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1–28. [Google Scholar]
- Cappelletti, M.; Presentato, A.; Piacenza, E.; Firrincieli, A.; Turner, R.J.; Zannoni, D. Biotechnology of Rhodococcus for the production of valuable compounds. Appl. Microbiol. Biotechnol. 2020, 104, 8567–8594. [Google Scholar] [CrossRef] [PubMed]
- Ceniceros, A.; Dijkhuizen, L.; Petrusma, M.; Medema, M.H. Genome-based exploration of the specialized metabolic capacities of the genus Rhodococcus. BMC Genom. 2017, 18, 593. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, C.C.; da Fonseca, M.M. The remarkable Rhodococcus erythropolis. Appl. Microbiol. Biotechnol. 2005, 67, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Sandmann, G. Antioxidant Protection from UV- and Light-Stress Related to Carotenoid Structures. Antioxidants 2019, 8, 219. [Google Scholar] [CrossRef] [Green Version]
- Dai, T.; Gupta, A.; Murray, C.K.; Vrahas, M.S.; Tegos, G.P.; Hamblin, M.R. Blue light for infectious diseases: Propionibacterium acnes, Helicobacter pylori, and beyond? Drug Resist. Updates 2012, 15, 223–236. [Google Scholar] [CrossRef] [Green Version]
- Orlandi, V.T.; Bolognese, F.; Chiodaroli, L.; Tolker-Nielsen, T.; Barbieri, P. Pigments influence the tolerance of Pseudomonas aeruginosa PAO1 to photodynamically induced oxidative stress. Microbiology 2015, 161, 2298–2309. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Halstead, F.D.; Thwaite, J.E.; Burt, R.; Laws, T.R.; Raguse, M.; Moeller, R.; Webber, M.A.; Oppenheim, B.A. Antibacterial Activity of Blue Light against Nosocomial Wound Pathogens Growing Planktonically and as Mature Biofilms. Appl. Environ. Microbiol. 2016, 82, 4006–4016. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ferrer-Espada, R.; Baglo, Y.; Gu, Y.; Dai, T. Antimicrobial Blue Light Inactivation of Neisseria gonorrhoeae: Roles of Wavelength, Endogenous Photosensitizer, Oxygen, and Reactive Oxygen Species. Lasers Surg. Med. 2019, 51, 815–823. [Google Scholar] [CrossRef]
- Guffey, J.S.; Wilborn, J. In vitro bactericidal effects of 405-nm and 470-nm blue light. Photomed. Laser Surg. 2006, 24, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, J.; Lim, W.; Jeon, S.; Kim, O.; Koh, J.T.; Kim, C.S.; Choi, H.; Kim, O. In vitro bactericidal effects of 625, 525, and 425 nm wavelength (red, green, and blue) light-emitting diode irradiation. Photomed. Laser Surg. 2013, 31, 554–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardu, M.; Bulut, S.; Kavakli, I.H. MerR and ChrR mediate blue light induced photo-oxidative stress response at the transcriptional level in Vibrio cholerae. Sci. Rep. 2017, 7, 40817. [Google Scholar] [CrossRef] [PubMed]
- Sumi, S.; Suzuki, Y.; Matsuki, T.; Yamamoto, T.; Tsuruta, Y.; Mise, K.; Kawamura, T.; Ito, Y.; Shimada, Y.; Watanabe, E.; et al. Light-inducible carotenoid production controlled by a MarR-type regulator in Corynebacterium glutamicum. Sci. Rep. 2019, 9, 13136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathews, M.M.; Sistrom, W.R. Intracellular location of carotenoid pigments and some respiratory enzymes in Sarcina lutea. J. Bacteriol. 1959, 78, 778–787. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, C.C. Adaptation of Rhodococcus erythropolis cells for growth and bioremediation under extreme conditions. Res. Microbiol. 2012, 163, 125–136. [Google Scholar] [CrossRef]
- Choi, T.R.; Song, H.S.; Han, Y.H.; Park, Y.L.; Park, J.Y.; Yang, S.Y.; Bhatia, S.K.; Gurav, R.; Kim, H.J.; Lee, Y.K.; et al. Enhanced tolerance to inhibitors of Escherichia coli by heterologous expression of cyclopropane-fatty acid-acyl-phospholipid synthase (cfa) from Halomonas socia. Bioprocess. Biosyst. Eng. 2020, 43, 909–918. [Google Scholar] [CrossRef]
- Hari, S.B.; Grant, R.A.; Sauer, R.T. Structural and Functional Analysis of E. coli Cyclopropane Fatty Acid Synthase. Structure 2018, 26, 1251–1258. [Google Scholar] [CrossRef] [Green Version]
- Patek, M.; Grulich, M.; Nesvera, J. Stress response in Rhodococcus strains. Biotechnol. Adv. 2021, 53, 107698. [Google Scholar] [CrossRef]
- de Carvalho, C.C.; Marques, M.P.; Hachicho, N.; Heipieper, H.J. Rapid adaptation of Rhodococcus erythropolis cells to salt stress by synthesizing polyunsaturated fatty acids. Appl. Microbiol. Biotechnol. 2014, 98, 5599–5606. [Google Scholar] [CrossRef]
- Solyanikova, I.P.; Suzina, N.E.; Emelyanova, E.V.; Polivtseva, V.N.; Pshenichnikova, A.B.; Lobanok, A.G.; Golovleva, L.A. Morphological, Physiological, and Biochemical Characteristics of a Benzoate-Degrading Strain Rhodococcus opacus 1CP under Stress Conditions. Mikrobiologiia 2017, 86, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Paper, M.; Glemser, M.; Haack, M.; Lorenzen, J.; Mehlmer, N.; Fuchs, T.; Schenk, G.; Garbe, D.; Weuster-Botz, D.; Eisenreich, W.; et al. Efficient Green Light Acclimation of the Green Algae Picochlorum sp. Triggering Geranylgeranylated Chlorophylls. Front. Bioeng. Biotechnol. 2022, 10, 689. [Google Scholar] [CrossRef] [PubMed]
- Woortman, D.V.; Fuchs, T.; Striegel, L.; Fuchs, M.; Weber, N.; Bruck, T.B.; Rychlik, M. Microalgae a Superior Source of Folates: Quantification of Folates in Halophile Microalgae by Stable Isotope Dilution Assay. Front Bioeng Biotechnol 2019, 7, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaigani, P.; Awad, D.; Redai, V.; Fuchs, M.; Haack, M.; Mehlmer, N.; Brueck, T. Oleaginous yeasts- substrate preference and lipid productivity: A view on the performance of microbial lipid producers. Microb. Cell Factories 2021, 20, 220. [Google Scholar] [CrossRef]
- Awad, D.; Brueck, T. Optimization of protein isolation by proteomic qualification from Cutaneotrichosporon oleaginosus. Anal. Bioanal. Chem. 2020, 412, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, T.; Melcher, F.; Rerop, Z.S.; Lorenzen, J.; Shaigani, P.; Awad, D.; Haack, M.; Prem, S.A.; Masri, M.; Mehlmer, N.; et al. Identifying carbohydrate-active enzymes of Cutaneotrichosporon oleaginosus using systems biology. Microb. Cell Factories 2021, 20, 205. [Google Scholar] [CrossRef]
- Tran, N.H.; Qiao, R.; Xin, L.; Chen, X.; Liu, C.; Zhang, X.; Shan, B.; Ghodsi, A.; Li, M. Deep learning enables de novo peptide sequencing from data-independent-acquisition mass spectrometry. Nat. Methods 2019, 16, 63–66. [Google Scholar] [CrossRef]
- Tran, N.H.; Rahman, M.Z.; He, L.; Xin, L.; Shan, B.; Li, M. Complete De Novo Assembly of Monoclonal Antibody Sequences. Sci Rep 2016, 6, 31730. [Google Scholar] [CrossRef] [Green Version]
- Tran, N.H.; Zhang, X.; Xin, L.; Shan, B.; Li, M. De novo peptide sequencing by deep learning. Proc. Natl. Acad. Sci. USA 2017, 114, 8247–8252. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J Mol Biol 2016, 428, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Mathews, M.M.; Sistrom, W.R. The function of the carotenoid pigments of Sarcina lutea. Arch. Mikrobiol. 1960, 35, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Thanapimmetha, A.; Suwaleerat, T.; Saisriyoot, M.; Chisti, Y.; Srinophakun, P. Production of carotenoids and lipids by Rhodococcus opacus PD630 in batch and fed-batch culture. Bioprocess Biosyst. Eng. 2017, 40, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Ram, S.; Mitra, M.; Shah, F.; Tirkey, S.R.; Mishra, S. Bacteria as an alternate biofactory for carotenoid production: A review of its applications, opportunities and challenges. J. Funct. Foods 2020, 67, 103867. [Google Scholar] [CrossRef]
- Davila Costa, J.S.; Silva, R.A.; Leichert, L.; Alvarez, H.M. Proteome analysis reveals differential expression of proteins involved in triacylglycerol accumulation by Rhodococcus jostii RHA1 after addition of methyl viologen. Microbiology 2017, 163, 343–354. [Google Scholar] [CrossRef]
- Wei, X.; Mingjia, H.; Xiufeng, L.; Yang, G.; Qingyu, W. Identification and biochemical properties of Dps (starvation-induced DNA binding protein) from cyanobacterium Anabaena sp. PCC 7120. IUBMB Life 2007, 59, 675–681. [Google Scholar] [CrossRef]
- Calhoun, L.N.; Kwon, Y.M. Structure, function and regulation of the DNA-binding protein Dps and its role in acid and oxidative stress resistance in Escherichia coli: A review. J. Appl. Microbiol. 2011, 110, 375–386. [Google Scholar] [CrossRef]
- Almiron, M.; Link, A.J.; Furlong, D.; Kolter, R. A novel DNA-binding protein with regulatory and protective roles in starved Escherichia coli. Genes Dev. 1992, 6, 2646–2654. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.; Meziane, T.; Yamasaki, H. A photocarotenogenic Rhodococcus sp. isolated from the symbiotic fern Azolla. Endocytobiosis Cell Res. 2004, 15, 350–355. [Google Scholar]
- Gebhard, S.; Humpel, A.; McLellan, A.D.; Cook, G.M. The alternative sigma factor SigF of Mycobacterium smegmatis is required for survival of heat shock, acidic pH and oxidative stress. Microbiology 2008, 154, 2786–2795. [Google Scholar] [CrossRef] [Green Version]
- Ondrusch, N.; Kreft, J. Blue and red light modulates SigB-dependent gene transcription, swimming motility and invasiveness in Listeria monocytogenes. PLoS ONE 2011, 6, e16151. [Google Scholar] [CrossRef] [Green Version]
- Humpel, A.; Gebhard, S.; Cook, G.M.; Berney, M. The SigF regulon in Mycobacterium smegmatis reveals roles in adaptation to stationary phase, heat, and oxidative stress. J. Bacteriol. 2010, 192, 2491–2502. [Google Scholar] [CrossRef] [Green Version]
- O’Donoghue, B.; NicAogain, K.; Bennett, C.; Conneely, A.; Tiensuu, T.; Johansson, J.; O’Byrne, C. Blue-Light Inhibition of Listeria monocytogenes Growth Is Mediated by Reactive Oxygen Species and Is Influenced by sigmaB and the Blue-Light Sensor Lmo0799. Appl. Environ. Microbiol. 2016, 82, 4017–4027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turunen, O.; Koskinen, S.; Kurkela, J.; Karhuvaara, O.; Hakkila, K.; Tyystjarvi, T. Roles of Close Homologues SigB and SigD in Heat and High Light Acclimation of the Cyanobacterium Synechocystis sp. PCC 6803. Life 2022, 12, 162. [Google Scholar] [CrossRef] [PubMed]
- Hakkila, K.; Valev, D.; Antal, T.; Tyystji Rvi, E.; Tyystji Rvi, T. Group 2 Sigma Factors are Central Regulators of Oxidative Stress Acclimation in Cyanobacteria. Plant Cell Physiol. 2019, 60, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Dorey, A.L.; Lee, B.H.; Rotter, B.; O’Byrne, C.P. Blue Light Sensing in Listeria monocytogenes Is Temperature-Dependent and the Transcriptional Response to It Is Predominantly SigB-Dependent. Front. Microbiol. 2019, 10, 2497. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Wang, Q.; Yang, M.; Chen, Z.; Li, J.; Wen, Y. Heat Shock Repressor HspR Directly Controls Avermectin Production, Morphological Development, and H2O2 Stress Response in Streptomyces avermitilis. Appl. Environ. Microbiol. 2021, 87, e00473-21. [Google Scholar] [CrossRef]
- Loewen, P.C.; Switala, J.; Triggs-Raine, B.L. Catalases HPI and HPII in Escherichia coli are induced independently. Arch Biochem Biophys 1985, 243, 144–149. [Google Scholar] [CrossRef]
- Heipieper, H.J.; Weber, F.J.; Sikkema, J.; Keweloh, H.; Debont, J.A.M. Mechanisms of Resistance of Whole Cells to Toxic Organic-Solvents. Trends Biotechnol. 1994, 12, 409–415. [Google Scholar] [CrossRef]
- Weber, F.J.; de Bont, J.A. Adaptation mechanisms of microorganisms to the toxic effects of organic solvents on membranes. Biochim. Biophys. Acta 1996, 1286, 225–245. [Google Scholar] [CrossRef]
- Sinensky, M. Homeoviscous adaptation--a homeostatic process that regulates the viscosity of membrane lipids in Escherichia coli. Proc. Natl. Acad. Sci. USA 1974, 71, 522–525. [Google Scholar] [CrossRef] [Green Version]
- Sundararaghavan, A.; Mukherjee, A.; Sahoo, S.; Suraishkumar, G.K. Mechanism of the oxidative stress-mediated increase in lipid accumulation by the bacterium, R. opacus PD630: Experimental analysis and genome-scale metabolic modeling. Biotechnol. Bioeng. 2020, 117, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Zhuang, W.Q.; Sahu, M.; Biswas, P.; Tang, Y.J. Cu-doped TiO(2) nanoparticles enhance survival of Shewanella oneidensis MR-1 under ultraviolet light (UV) exposure. Sci. Total Environ. 2011, 409, 4635–4639. [Google Scholar] [CrossRef] [PubMed]
- Monier, J.M.; Lindow, S.E. Differential survival of solitary and aggregated bacterial cells promotes aggregate formation on leaf surfaces. Proc. Natl. Acad. Sci. USA 2003, 100, 15977–15982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Chu, Z.; Ruan, Z.; Wang, X.; Dai, T.; Hu, X. Changes of Intracellular Porphyrin, Reactive Oxygen Species, and Fatty Acids Profiles During Inactivation of Methicillin-Resistant Staphylococcus aureus by Antimicrobial Blue Light. Front. Physiol. 2018, 9, 1658. [Google Scholar] [CrossRef]
- Tsitko, I.V.; Zaitsev, G.M.; Lobanok, A.G.; Salkinoja-Salonen, M.S. Effect of aromatic compounds on cellular fatty acid composition of Rhodococcus opacus. Appl. Environ. Microbiol. 1999, 65, 853–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S.K.; Gurav, R.; Choi, T.R.; Han, Y.H.; Park, Y.L.; Jung, H.R.; Yang, S.Y.; Song, H.S.; Yang, Y.H. A clean and green approach for odd chain fatty acids production in Rhodococcus sp. YHY01 by medium engineering. Bioresour. Technol. 2019, 286, 121383. [Google Scholar] [CrossRef]
- Park, Y.K.; Dulermo, T.; Ledesma-Amaro, R.; Nicaud, J.M. Optimization of odd chain fatty acid production by Yarrowia lipolytica. Biotechnol. Biofuels 2018, 11, 158. [Google Scholar] [CrossRef] [Green Version]
- Srirangan, K.; Bruder, M.; Akawi, L.; Miscevic, D.; Kilpatrick, S.; Moo-Young, M.; Chou, C.P. Recent advances in engineering propionyl-CoA metabolism for microbial production of value-added chemicals and biofuels. Crit. Rev. Biotechnol. 2017, 37, 701–722. [Google Scholar] [CrossRef]
- Otzen, C.; Bardl, B.; Jacobsen, I.D.; Nett, M.; Brock, M. Candida albicans utilizes a modified beta-oxidation pathway for the degradation of toxic propionyl-CoA. J. Biol. Chem. 2014, 289, 8151–8169. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcription Factors Involved in Oxidative Stress | ||||||||
|---|---|---|---|---|---|---|---|---|
| 40 h | 122 h | |||||||
| Accession Number | Description | KO-ID | Identified Unique Peptides | Significance | Fold Change | Identified Unique Peptides | Significance | Fold Change |
| WP_019747469.1 | SigB; RNA polymerase sigma-B factor | K03090 | 1 | 3.8 | 2.38 | 1 | 2.2 | 3.88 |
| WP_020906601.1 | hspR; MerR family transcriptional regulator, heat shock protein hspR | K13640 | 2 | 2.36 | 1.38 | 1 | 4.19 | 4.51 |
| WP_020906739.1 | MULTISPECIES: RNA polymerase sigma factor SigF [Rhodococcus] | K03090 | 1 | 4.35 | 2.38 | n.d. | n.d. | n.d. |
| WP_060939090.1 | hspR; MerR family transcriptional regulator, heat shock protein hspR | K13640 | 5 | 4.07 | 1.39 | 4 | 4.52 | 3.29 |
| Catalases and Peroxidases | ||||||||
| WP_019749140.1 | BCP, PRXQ, DOT5; thioredoxin-dependent peroxiredoxin [EC:1.11.1.24] | K03564 | 8 | 0.11 | 0.99 | 8 | 3.13 | 0.49 |
| WP_021346030.1 | katE, CAT, catB, srpA; catalase [EC:1.11.1.6] | K03781 | 14 | 26.31 | 2.6 | 14 | 0.19 | 1.06 |
| WP_003940303.1 | SOD; superoxide dismutase, Fe-Mn family [EC:1.15.1.1] | K04564 | 5 | 47.57 | 3.06 | 3 | 4.34 | 2.8 |
| WP_003942119.1 | ahpC; lipoyl-dependent peroxiredoxin subunit C [EC:1.11.1.28] | K24126 | 16 | 47.31 | 2.35 | 16 | 3.56 | 2.16 |
| WP_060938296.1 | katG; catalase-peroxidase [EC:1.11.1.21] | K03782 | 27 | 23.13 | 0.45 | 24 | 1.78 | 0.63 |
| Other Stress-Related Proteins | ||||||||
| WP_003942530.1 | trxA; thioredoxin | K03671 | 2 | 9.59 | 0.43 | 3 | 4.05 | 2.86 |
| WP_019747464.1 | dps; starvation-inducible DNA-binding protein | K04047 | 14 | 0.32 | 0.98 | 14 | 2.98 | 2.0 |
| WP_019749386.1 | dnaJ; molecular chaperone DnaJ | K03686 | 12 | 5.42 | 2.39 | 13 | 0.78 | 1.28 |
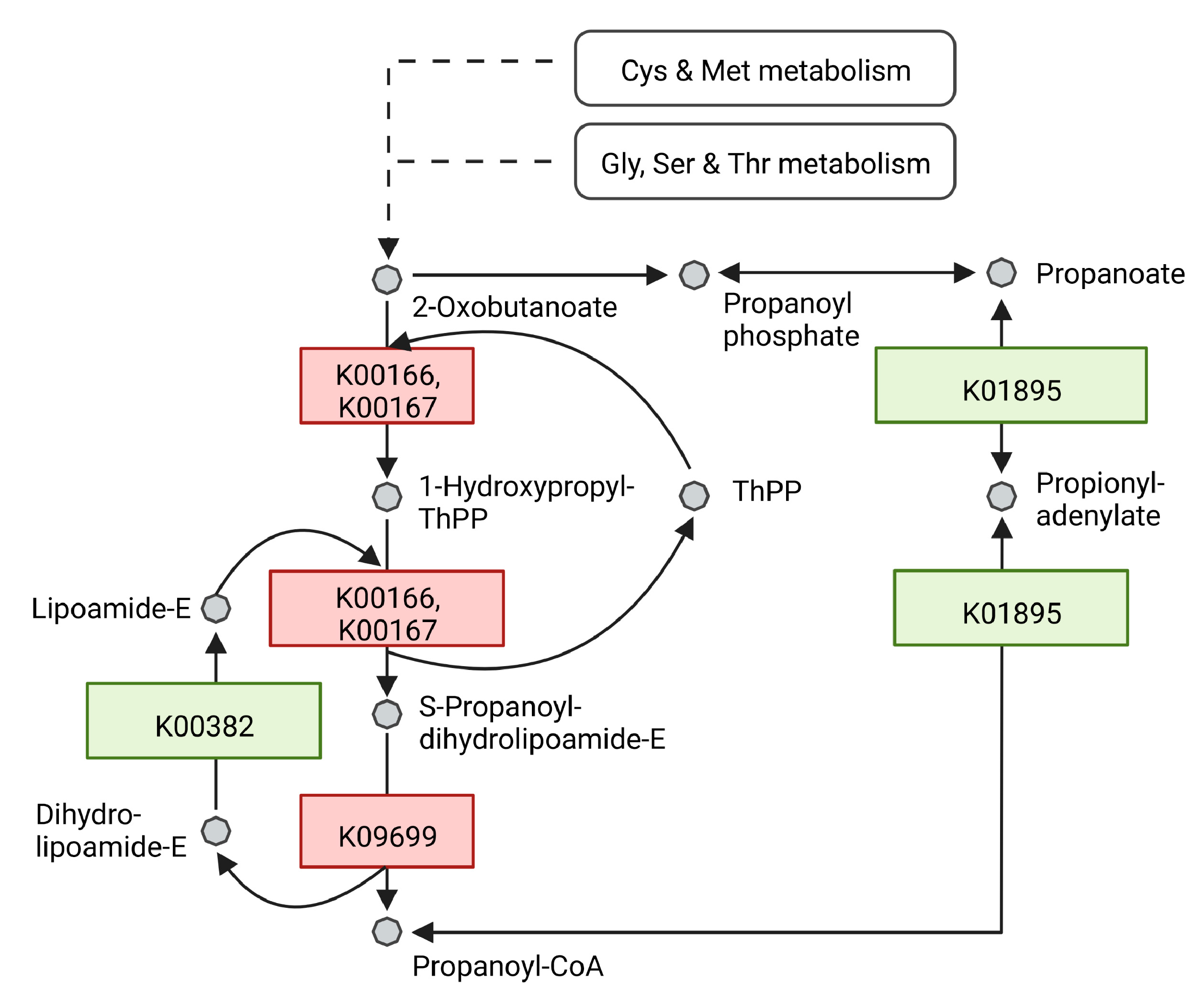
| Propanoate Pathway | ||||||||
|---|---|---|---|---|---|---|---|---|
| 40 h | 122 h | |||||||
| Accession Number | Description | KO-ID | Identified Unique Peptides | Significance | Fold Change | Identified Unique Peptides | Significance | Fold Change |
| WP_019745948.1 | acs; acetyl-CoA synthetase [EC:6.2.1.1] | K01895 | 20 | 26.37 | 0.46 | 15 | 13.6 | 0.08 |
| WP_020970089.1 | bkdB; 2-oxoisovalerate dehydrogenase E2 component (dihydrolipoyl transacylase) [EC:2.3.1.168] | K09699 | 7 | 29.17 | 3.05 | 7 | 4.78 | 3.1 |
| WP_060938768.1 | bkdA1; 2-oxoisovalerate dehydrogenase E1 component alpha subunit [EC:1.2.4.4] | K00166 | 22 | 20.92 | 0.59 | 19 | 4.81 | 0.42 |
| WP_060938769.1 | bkdA2; 2-oxoisovalerate dehydrogenase E1 component beta subunit [EC:1.2.4.4] | K00167 | 11 | 28.47 | 0.57 | 11 | 8.94 | 0.23 |
| WP_060938770.1 | bkdB; 2-oxoisovalerate dehydrogenase E2 component (dihydrolipoyl transacylase) [EC:2.3.1.168] | K09699 | 21 | 23.64 | 0.55 | 18 | 5.08 | 0.35 |
| WP_060938993.1 | bkdA2; 2-oxoisovalerate dehydrogenase E1 component beta subunit [EC:1.2.4.4] | K00167 | 11 | 30.2 | 2.63 | 12 | 4.09 | 2.62 |
| WP_060938994.1 | bkdA1; 2-oxoisovalerate dehydrogenase E1 component alpha subunit [EC:1.2.4.4] | K00166 | 12 | 50.3 | 3.82 | 13 | 4.41 | 2.73 |
| WP_060939079.1 | bccA, pccA; acetyl-CoA/propionyl-CoA carboxylase, biotin carboxylase, biotin carboxyl carrier protein [EC:6.4.1.2 6.4.1.3 6.3.4.14] | K11263 | 31 | 58.35 | 3.28 | 34 | 5.63 | 3.92 |
| WP_060939587.1 | acs; acetyl-CoA synthetase [EC:6.2.1.1] | K01895 | 23 | 33.08 | 2.28 | 20 | 4.29 | 2.39 |
| WP_174531767.1 | DLD, lpd, pdhD; dihydrolipoamide dehydrogenase [EC:1.8.1.4] | K00382 | 22 | 30.59 | 0.41 | 16 | 8.77 | 0.19 |
| Cyclopropane-Fatty-Acyl-Phospholipid Synthase Family | ||||||||
| WP_060938639.1 | cfa; cyclopropane-fatty-acyl-phospholipid synthase [EC:2.1.1.79] | K00574 | 2 | 76.43 | 22.02 | 1 | 6.24 | 18.92 |
| WP_060938640.1 | cfa; cyclopropane-fatty-acyl-phospholipid synthase [EC:2.1.1.79] | K00574 | 3 | 105.03 | 43.79 | 4 | 11.05 | 31.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Engelhart-Straub, S.; Cavelius, P.; Hölzl, F.; Haack, M.; Awad, D.; Brueck, T.; Mehlmer, N. Effects of Light on Growth and Metabolism of Rhodococcus erythropolis. Microorganisms 2022, 10, 1680. https://doi.org/10.3390/microorganisms10081680
Engelhart-Straub S, Cavelius P, Hölzl F, Haack M, Awad D, Brueck T, Mehlmer N. Effects of Light on Growth and Metabolism of Rhodococcus erythropolis. Microorganisms. 2022; 10(8):1680. https://doi.org/10.3390/microorganisms10081680
Chicago/Turabian StyleEngelhart-Straub, Selina, Philipp Cavelius, Fabian Hölzl, Martina Haack, Dania Awad, Thomas Brueck, and Norbert Mehlmer. 2022. "Effects of Light on Growth and Metabolism of Rhodococcus erythropolis" Microorganisms 10, no. 8: 1680. https://doi.org/10.3390/microorganisms10081680
APA StyleEngelhart-Straub, S., Cavelius, P., Hölzl, F., Haack, M., Awad, D., Brueck, T., & Mehlmer, N. (2022). Effects of Light on Growth and Metabolism of Rhodococcus erythropolis. Microorganisms, 10(8), 1680. https://doi.org/10.3390/microorganisms10081680