
Virulence Characteristics and Distribution of the Pathogen Listeria ivanovii in the Environment and in Food

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Virulence Characters of L. ivanovii
3. L. ivanovii Invasiveness
4. L. ivanovii Persistence and Tolerance to Harsh Conditions
5. Environmental Distribution of L. ivanovii
6. L. ivanovii Infection Cases in Humans
7. L. ivanovii Infection Cases in Animals
8. Occurrence of L. ivanovii in Food
9. Methods of Isolation and Identification of L. ivanovii
9.1. Isolation Methods
9.2. Molecular Identification and Detection
10. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Carlin, C.R.; Roof, S.; Wiedmann, M. Assessment of Reference Method Selective Broth and Plating Media with 19 Listeria Species Highlights the Importance of Including Diverse Species in Listeria Method Evaluations. J. Food Prot. 2022, 85, 494–510. [Google Scholar] [CrossRef]
- Orsi, R.H.; Wiedmann, M. Characteristics and distribution of Listeria spp., including Listeria species newly described since 2009. Appl. Microbiol. Biotechnol. 2016, 100, 5273–5287. [Google Scholar] [CrossRef]
- Vázquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domínguez-Bernal, G.; Goebel, W.; González-Zorn, B.; Wehland, J.; Kreft, J. Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef]
- Schmid, M.W.; Ng, E.Y.; Lampidis, R.; Emmerth, M.; Walcher, M.; Kreft, J.; Goebel, W.; Wagner, M.; Schleifer, K.H. Evolutionary history of the genus Listeria and its virulence genes. Syst. Appl. Microbiol. 2005, 28, 1–18. [Google Scholar] [CrossRef]
- Ivanov, I. Establishment of non-motile strains of Listeria monocytogenes type 5. In Problems of Listeriosis; Woodbine, M., Ed.; Leicester University Press: Leicester, UK, 1975; pp. 18–26. [Google Scholar]
- Seeliger, H.P.R.; Welshimer, H.J. Genus Listeria. In Bergey’s Manual of Determinative Bacteriology, 8th ed.; Buchanan, R.E., Gibbons, N.E., Eds.; The Williams and Wilkins Co.: Baltimore, MD, USA, 1974; pp. 593–596. [Google Scholar]
- Seeliger, H.P.R.; Rocourt, J.; Schrettenbrunner, A.; Grimont, P.A.D.; Jones, D. Notes: Listeria ivanovii sp. nov. Int. J. Syst. Evolut. Microbiol. 1984, 34, 336–337. [Google Scholar] [CrossRef]
- Boerlin, P.; Rocourt, J.; Grimont, F.; Grimont, P.A.D.; Jaquet, C.; Piffaretti, J.C. Listeria ivanovii subsp. londoniensis subsp. nov. Int. J. Syst. Evolut. Microbiol. 1992, 42, 63–69. [Google Scholar] [CrossRef][Green Version]
- Hupfeld, M.; Trasanidou, D.; Ramazzini, L.; Klumpp, J.; Loessner, M.J.; Kilcher, S.A. functional type II-A CRISPR-Cas system from Listeria enables efficient genome editing of large non-integrating bacteriophage. Nucleic Acids Res. 2018, 46, 6920–6933. [Google Scholar] [CrossRef]
- Beye, M.; Gouriet, F.; Michelle, C.; Casalta, J.P.; Habib, G.; Raoult, D.; Fournier, P.E. Genome analysis of Listeria ivanovii strain G770 that caused a deadly aortic prosthesis infection. New Microbes New Infect. 2016, 10, 87–92. [Google Scholar] [CrossRef]
- Guillet, C.; Join-Lambert, O.; Le Monnier, A.; Leclercq, A.; Mechai, F.; Mamzer-Bruneel, M.F.; Bielecka, M.K.; Scortti, M.; Disson, O.; Berche, P.; et al. Human listeriosis caused by Listeria ivanovii. Emerg. Infect. Dis. 2010, 16, 136–138. [Google Scholar] [CrossRef]
- Blot, M.; Disson, O.; Leclercq, A.; Moura, A.; Bracq-Dieye, H.; Thouvenot, P.; Valès, G.; Burroni, B.; Lupo, A.; Lecuit, M.; et al. Listeria-Associated Lymphadenitis: A Series of 11 Consecutive Cases and Review of the Literature. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2022; Volume 9, p. ofab598. [Google Scholar]
- Gan, L.; Mao, P.; Jiang, H.; Zhang, L.; Liu, D.; Cao, X.; Wang, Y.; Wang, Y.; Sun, H.; Huang, Y.; et al. Two prevalent Listeria ivanovii subsp. ivanovii clonal strains with different virulence exist in wild rodents and pikas of China. Front. Vet. Sci. 2020, 7, 88. [Google Scholar] [CrossRef]
- Domínguez-Bernal, G.; Müller-Altrock, S.; González-Zorn, B.; Scortti, M.; Herrmann, P.; Monzó, G.H.; Lacharme, L.; Kreft, J.; Vázquez-Boland, A.J. A spontaneous genomic deletion in Listeria ivanovii identifies LIPI-2, a species-specific pathogenicity island encoding sphingomyelinase and numerous internalins. Mol. Microbiol. 2006, 59, 415–432. [Google Scholar] [CrossRef]
- Gedde, M.M.; Higgins, D.E.; Tilney, L.G.; Portnoy, D.A. Role of listeriolysin O in cell-to-cell spread of Listeria monocytogenes. Infect. Immun. 2000, 68, 999–1003. [Google Scholar] [CrossRef]
- Buchrieser, C.; Rusniok, C.; Garrido, P.; Hain, T.; Scortti, M.; Lampidis, R.; Kärst, U.; Chakraborty, T.; Cossart, P.; Kreft, J.; et al. Complete genome sequence of the animal pathogen Listeria ivanovii, which provides insights into host specificities and evolution of the genus Listeria. J. Bacteriol. 2011, 193, 6787–6788. [Google Scholar] [CrossRef]
- Disson, O.; Lecuit, M. In vitro and in vivo models to study human listeriosis: Mind the gap. Microb. Infect. 2013, 15, 971–980. [Google Scholar] [CrossRef]
- Van Ngo, H.; Bhalla, M.; Chen, D.Y.; Ireton, K. A role for host cell exocytosis in InlB-mediated internalization of Listeria monocytogenes. Cell. Microbiol. 2017, 19, e12768. [Google Scholar] [CrossRef]
- Engelbrecht, F.; Domínguez-Bernal, G.; Hess, J.; Dickneite, C.; Greiffenberg, L.; Lampidis, R.; Raffelsbauer, D.; Daniels, J.J.; Kreft, J.; Kaufmann, S.H.; et al. A novel PrfA-regulated chromosomal locus, which is specific for Listeria ivanovii, encodes two small, secreted internalins and contributes to virulence in mice. Mol. Microbiol. 1998, 30, 405–417. [Google Scholar] [CrossRef]
- González-Zorn, B.; Domínguez-Bernal, G.; Suárez, M.; Ripio, M.T.; Vega, Y.; Novella, S.; Rodríguez, A.; Chico, I.; Tierrez, A.; Vázquez-Boland, J.A. SmcL, a novel membrane-damaging virulence factor in Listeria. Int. J. Med. Microbiol. 2000, 290, 369–374. [Google Scholar] [CrossRef]
- Gouin, E.; Dehoux, P.; Mengaud, J.; Kocks, C.; Cossart, P. iactA of Listeria ivanovii, although distantly related to Listeria monocytogenes actA, restores actin tail formation in an L. monocytogenes actA mutant. Infect. Immun. 1995, 63, 2729–2737. [Google Scholar] [CrossRef]
- Karunasagar, I.; Krohne, G.; Goebel, W. Listeria ivanovii is capable of cell-to-cell spread involving actin polymerization. Infect. Immun. 1993, 61, 162–169. [Google Scholar] [CrossRef]
- Hain, T.; Chatterjee, S.S.; Ghai, R.; Kuenne, C.T.; Billion, A.; Steinweg, C.; Domann, E.; Kärst, U.; Jänsch, L.; Wehland, J.; et al. Pathogenomics of Listeria spp. Int. J. Med. Microbiol. 2007, 297, 541–557. [Google Scholar] [CrossRef]
- Kuenne, C.; Billion, A.; Mraheil, M.A.; Strittmatter, A.; Daniel, R.; Goesmann, A.; Barbuddhe, S.; Hain, T.; Chakraborty, T. Reassessment of the Listeria monocytogenes pan-genome reveals dynamic integration hotspots and mobile genetic elements as major components of the accessory genome. BMC Genom. 2013, 14, 47. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wang, Y.; Wang, Y.; Li, H.; Luo, L.; Wang, P.; Zhang, L.; Li, H.; Liu, J.; Lu, L.; et al. Prevalence and Characteristics of Listeria ivanovii Strains in Wild Rodents in China. Vector Borne Zoonotic Dis. 2019, 19, 8–15. [Google Scholar] [CrossRef]
- Alvarez-Ordóñez, A.; Leong, D.; Morgan, C.A.; Hill, C.; Gahan, C.G.; Jordan, K. Occurrence, persistence, and virulence potential of Listeria ivanovii in foods and food processing environments in the Republic of Ireland. BioMed. Res. Int. 2015, 2015, 350526. [Google Scholar] [CrossRef]
- Ammendolia, M.G.; Superti, F.; Bertuccini, L.; Chiarini, F.; Conte, M.P.; Cipriani, D.; Seganti, L.; Longhi, C. Invasive pathway of Listeria ivanovii in human amnion-derived WISH cells. Int. J. Immunopathol. Pharmacol. 2007, 20, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Rocha, C.E.; Mol, J.P.S.; Garcia, L.N.N.; Costa, L.F.; Santos, R.L.; Paixao, T.A. Comparative experimental infection of Listeria monocytogenes and Listeria ivanovii in bovine trophoblasts. PLoS ONE 2017, 12, e0176911. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Jiang, M.; Ren, C.; Liu, S.; Pu, Q.; Goldfine, H.; Shen, H.; Wang, C. Listeria ivanovii Infection in Mice: Restricted to the Liver and Lung with Limited Replication in the Spleen. Front. Microbiol. 2016, 7, 790. [Google Scholar] [CrossRef] [PubMed]
- Borovuk, I.; Zazharska, N. Evaluation of broiler meat in experimental listeriosis. J. Adv. Vet. Anim. Res. 2022, 9, 155–165. [Google Scholar] [CrossRef]
- Nyenje, M.E.; Green, E.; Ndip, R.N. Biofilm formation and adherence characteristics of Listeria ivanovii strains isolated from ready-to-eat foods in Alice, South Africa. Scient. World J. 2012, 2012, 873909. [Google Scholar] [CrossRef]
- Vázquez-Villanueva, J.; Orgaz, B.; Ortiz, S.; López, V.; Martínez-Suárez, J.V.; Sanjose, C. Predominance and persistence of a single clone of Listeria ivanovii in a Manchego cheese factory over 6 months. Zoonoses Public Health 2010, 57, 402–410. [Google Scholar] [CrossRef]
- Chmielowska, C.; Korsa, D.; Szmulkowska, B.; Krop, A.; Lipka, K.; Krupińska, M.; Bartosik, D. Genetic Carriers and Genomic Distribution of cadA6-A Novel Variant of a Cadmium Resistance Determinant Identified in Listeria spp. Int. J. Mol. Sci. 2020, 21, 8713. [Google Scholar] [CrossRef]
- Müller, A.; Rychli, K.; Zaiser, A.; Wieser, C.; Wagner, M.; Schmitz-Esser, S. The Listeria monocytogenes transposon Tn6188 provides increased tolerance to various quaternary ammonium compounds and ethidium bromide. FEMS Microbiol. Lett. 2014, 361, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Bolten, S.; Harrand, A.S.; Skeens, J.; Wiedmann, M. Nonsynonymous Mutations in fepR Are Associated with Adaptation of Listeria monocytogenes and Other Listeria spp. to Low Concentrations of Benzalkonium Chloride but Do Not Increase Survival of L. monocytogenes and Other Listeria spp. after Exposure to Benzalkonium Chloride Concentrations Recommended for Use in Food Processing Environments. Appl. Environ. Microbiol. 2022, 88, e0048622. [Google Scholar] [PubMed]
- Longhi, C.; Ammendolia, M.G.; Conte, M.; Seganti, L.; Iosi, F.; Superti, F. Listeria ivanovii ATCC 19119 strain behaviour is modulated by iron and acid stress. Food Microbiol. 2014, 42, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Rawool, D.B.; Malik, S.V.S.; Shakuntala, I.; Sahare, A.M.; Barbuddhe, S.B. Detection of multiple virulence-associated genes in Listeria monocytogenes isolated from bovine mastitis cases. Int. J. Food Microbiol. 2007, 113, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Elezebeth, G.; Malik, S.V.S.; Chaudhari, S.P.; Barbuddhe, S.B. The occurrence of Listeria species and antibodies against listeriolysin-O in naturally infected goats. Small Rumin. Res. 2007, 67, 173–178. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, L.; Lan, R.; Salazar, J.K.; Liu, J.; Xu, J.; Ye, C. Isolation and characterization of Listeria species from rodents in natural environments in China. Emerg. Microbes Infect. 2017, 6, e44. [Google Scholar] [CrossRef]
- Akca, D.; Buyuk, F.; Celik, E.; Saglam, A.G.; Otlu, S.; Dag, S.; Celebi, O.; Coskun, M.R.; Buyuk, E.; Karakurt, E.; et al. Phylogenetic positioning of Listeria ivanovii identified in aborted sheep in Kars Region (Turkey). Thai J. Vet. Med. 2022, 52, 145–150. [Google Scholar]
- Abuhatab, E.; Naguib, D.; Abdou, A.; Gwida, M.; Elgohary, A. Genetic Characterization and Antibiogram Profiles of Listeria species Isolated from Poultry and Poultry Handlers. J. Adv. Vet. Res. 2022, 12, 205–210. [Google Scholar]
- Ahimeda, H.M.; Hikoa, A.; Abdellaha, A.; Muktarb, Y.D.; Gutema, F.D. Isolation and multidrug drug resistance profile of Listeria species in selected Dairy Farm’s Operational stages in Oromia Regional State, Ethiopia. Sci. Afr. 2022, 16, e01167. [Google Scholar] [CrossRef]
- Terentjeva, M.; Šteingolde, Z.; Meistere, I.; Elferts, D.; Avsejenko, J.; Streikiša, M.; Gradovska, S.; Alksne, L.; Ķibilds, J.; Bērziņš, A. Prevalence, Genetic Diversity and Factors Associated with Distribution of Listeria monocytogenes and Other Listeria spp. in Cattle Farms in Latvia. Pathogens. 2021, 10, 851. [Google Scholar] [CrossRef]
- Palacios-Gorba, C.; Moura, A.; Leclercq, A.; Gómez-Martín, A.; Gomis, J.; Jiménez-Trigos, E.; Mocé, M.L.; Lecuit, M.; Quereda, J.J. Listeria spp. Isolated from Tonsils of Wild Deer and Boars: Genomic Characterization. Appl. Environ. Microbiol. 2021, 87, e02651–e02720. [Google Scholar] [CrossRef] [PubMed]
- Meshref, L.; Pichon, M.; Burucoa, C.; Nusser, S.H.A.; Moura, A.; Garcia-Garcera, M.; Lecuit, M. Listeria monocytogenes faecal carriage is common and depends on the gut microbiota. Nat. Commun. 2021, 12, 6826. [Google Scholar]
- Ramage, C.P.; Low, J.C.; McLauchlin, J.; Donachie, W. Characterisation of Listeria ivanovii isolates from the UK using pulsed-field gel electrophoresis. FEMS Microbiol. Lett. 2006, 170, 349–353. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Low, J.C.; Donachie, W. A Review of Listeria monocytogenes and Listeriosis. Vet. J. 1997, 153, 9–29. [Google Scholar] [CrossRef]
- Dunnett, E.; Florea, L.; Thurston, L.; Floyd, T.; Collins, R.; Otter, A. Deaths of weaned lambs with visceral Listeria ivanovii infections. Vet. Rec. Case Rep. 2020, 8, e001254. [Google Scholar] [CrossRef]
- Della Rosa, P.; Colque Caro, L.A.; Cantón, G.J.; Morrell, E.L.; Hecker, Y.P.; Paolicchi, F.A.; Fiorentino, M.A. Aborto ovino asociado a Listeria ivanovii. In Proceedings of the XV Congreso Argentino de Microbiología, Buenos Aires, Argentina, 25–27 September 2019; Available online: https://www.researchgate.net/publication/338404312_Aborto_ovino_asociado_a_Listeria_ivanovii. (accessed on 23 July 2022).
- Meshref, A.; Zeinhom, M.; Abdel-Atty, N.S. Occurrence and distribution of Listeria species in some Egyptian foods. Alex. J. Vet. Sci. 2015, 46, 42–47. [Google Scholar] [CrossRef]
- Aksoy, A.; Sezer, Ç.; Vatansever, L.; Gulbaz, G. Presence and antibiotic resistance of Listeria monocytogenes in raw milk and dairy products. Kafkas Üniversitesi Veteriner Fakültesi Dergisi 2018, 24, 416–421. [Google Scholar]
- Guerra, M.M.; McLauchlin, J.; Bernardo, F.A. Listeria in ready-to-eat and unprocessed foods produced in Portugal. Food Microbiol. 2001, 18, 423–429. [Google Scholar] [CrossRef]
- Aygun, O.; Pehlivanlar, S. Listeria spp. in the raw milk and dairy products in Antakya, Turkey. Food Control 2006, 17, 676–679. [Google Scholar] [CrossRef]
- Gallegos, J.M.; Vanegas, M.C.; Albarracín, Y.; Máttar, S.; Poutou, R.A.; Carrascal, A.K. Frequency of isolation of Listeria species in different retail foods in Colombia. Anim. Prod. Res. Adv. 2008, 4, 9–18. [Google Scholar] [CrossRef][Green Version]
- Arslan, S.; Özdemir, F. Prevalence and antimicrobial resistance of Listeria spp. in homemade white cheese. Food Control 2008, 19, 360–363. [Google Scholar] [CrossRef]
- Latorre, L.; Fraccalvieri, R.; Parisi, S.; Santagada, G.; Normanno, G. Studio sulla contaminazione da Listeria spp. e Listeria monocytogenes in latte e prodotti lattiero-caseari ovi-caprini. Ind. Aliment. 2008, XLVII, 737–744. [Google Scholar]
- Mengesha, D.; Zewde, B.M.; Toquin, M.T.; Kleer, J.; Hildebrandt, G.; Gebreyes, W.A. Lebensmittelhygiene-Vorkommen und Verteilung von Listeria monocytogenes und anderen Listeria spp. in verzehrsfertigen Lebensmitteln und rohem Fleisch. Berliner und Munchener Tierarztliche Wochenschrift 2009, 122, 20–34. [Google Scholar] [PubMed]
- Büyükyörük, S.; Göksoy, E.Ö. Aydın ilinde satışa sunulan köy peynirlerinde Listeria varlığının araştırılması. Uludağ Üniversitesi Vet. Fakültesi Derg. 2011, 30, 9–12. [Google Scholar]
- Ramírez Mérida, L.G.; Morón de Salim, A.; Alfieri Graterol, A.Y.; Gamboa, O. Detección de Listeria monocytogenes en queso blanco criollo, mediante la reacción en cadena de la polimerasa (PCR). Archivos Latinoamericanos de Nutrición 2010, 60, 254–260. [Google Scholar] [PubMed]
- Sağun, E.; Sancak, Y.C.; İşleyici, Ö.; Ekici, K. The presence and prevalence of Listeria species in milk and herby cheese in and around Van. Turk. J. Vet. Anim. Sci. 2001, 25, 15–19. [Google Scholar]
- Cokal, Y.; Dagdelen, A.; Cenet, O.; Gunsen, U. Presence of L. monocytogenes and some bacterial pathogens in two Turkish traditional foods, Mihalic cheese and Hosmerim dessert. Food Control 2012, 26, 337–340. [Google Scholar] [CrossRef]
- Osaili, T.M.; Al-Nabulsi, A.A.; Taha, M.H.; Al-Holy, M.A.; Alaboudi, A.R.; Al-Rousan, W.M.; Shaker, R.R. Occurrence and antimicrobial susceptibility of Listeria monocytogenes isolated from brined white cheese in Jordan. J. Food Sci. 2012, 77, M528–M532. [Google Scholar] [CrossRef]
- Alzubaidy, Z.M.; Kakey, S.I.; Ali, J.F. Isolation and identification of Listeria moncytogenes by PCR from some food sources in Erbil city. Euphrates J. Agric. Sci. 2013, 5, 14–26. [Google Scholar]
- Kaptan, B. Prevalence of Listeria spp. and L. monocytogenes in homemade pottery cheese. Tekirdağ Ziraat Fakültesi Dergisi 2016, 13, 76–87. [Google Scholar]
- Kizanlik, P.K.; Göksoy, E.Ö. Microbiological quality evaluation of various types of cheese. Erciyes Üniversitesi Vet. Fakültesi Derg. 2018, 15, 86–93. [Google Scholar]
- Şanlıbaba, P.; Tezel, B.U.; Çakmak, G.A. Detection of Listeria spp. in raw milk and dairy products retailed in Ankara. Gıda 2018, 43, 273–282. [Google Scholar] [CrossRef]
- Şanlıbaba, P.; Tezel, B.U. Prevalence and characterization of Listeria species from raw milk and dairy products from çanakkale province. Turk. J. Agric. Food Sci. Technol. 2018, 6, 61–64. [Google Scholar] [CrossRef]
- Albastami, I.; Wajiej, A.H.; Aburagaegah, S. Microbiological study on Listeria species isolated from some food products of animal origin. Damanhour J. Vet. Sci. 2020, 4, 15–19. [Google Scholar] [CrossRef]
- Sobhy, M.I.; Sayed, M.; Walaa, E. Influence of essential oils on the viability of Listeria monocytogenes. Assiut Vet. Med. J. 2022, 68, 78–87. [Google Scholar] [CrossRef]
- Kamat, A.S.; Nair, P.M. Incidence of Listeria species in Indian seafoods and meat. J. Food Saf. 1994, 14, 117–130. [Google Scholar] [CrossRef]
- Dominguez, C.; Gomez, I.; Zumalacarregui, J. Prevalence and contamination levels of Listeria monocytogenes in smoked fish and pate sold in Spain. J. Food Prot. 2001, 64, 2075–2077. [Google Scholar] [CrossRef]
- Hassan, Z.; Purwati, E.; Radu, S.; Rahim, R.A.; Rahim, R.A.; Rusul, G. Prevalence of Listeria spp and Listeria monocytogenes in meat and fermented fish in Malaysia. Southeast Asian J. Trop. Med. Public Health 2001, 32, 402–407. [Google Scholar]
- Salihu, M.D.; Junaidu, U.; Manga, S.B.; Gulumbe, M.L.; Magaji, A.A.; Ahmed, A.; Adamu, A.I.; Schittu, A.; Balarabe, I. Occurrence of Listeria monocytogenes in smoked fish in Sokoto, Nigeria. Afr. J. Biotechnol. 2008, 7, 3082–3084. [Google Scholar]
- Masuda, T.; Iwaya, M.; Miura, H.; Kokubo, Y.; Maruyama, T. Occurrence of Listeria species in fresh seafood. Food Hyg. Saf. Sci. 1992, 33, 599–602. [Google Scholar] [CrossRef]
- Bianchini, M.; Arias, M.L.; Herrera, C.; Zuniga, C. Listeria monocytogenes incidence and evaluation of the sanitary quality of filleted fresh fish from the Metropolitan Area of San José. Arch. Latinoam. Nutr. 1999, 49, 358–362. [Google Scholar] [PubMed]
- Ripabelli, G.; Sammarco, M.L.; Fanelli, I.; Grasso, G.M. Ricerca di Salmonella, Listeria spp., Vibrio spp. e Yersinia enterocolitica in prodotti ittici congelati e surgelati del commercio e correlazione con gli indici di contaminazione fecale: Implicazioni in Sanità Pubblica. Ann. Ig. 2004, 16, 531–539. [Google Scholar] [PubMed]
- Akkaya, L.; Atabay, H.İ.; Gök, V.; Kara, R. Detection of Listeria species in fresh fish and fish market environment by IMS technique in Turkey. Archiv fur Lebensmittelhygiene 2011, 62, 16–19. [Google Scholar]
- Modaresi, R.; Mardani, K.; Tukmechi, A.; Ownagh, A. Prevalence of Listeria spp. in fish obtained from Urmia fish markets. Afr. J. Microbiol. Res. 2011, 5, 5398–5401. [Google Scholar] [CrossRef]
- Voidarou, C.; Alexopoulos, A.; Plessas, S.; Noussias, H.; Stavropoulou, E.; Fotou, K.; Tzora, A.; Skoufos, I.; Bezirtzoclou, E.; Demertzi-Akrida, K. Microbiological quality of grey-mullet roe. Anaerobe 2011, 17, 273–275. [Google Scholar] [CrossRef]
- Momtaz, H.; Yadollahi, S. Molecular characterization of Listeria monocytogenes isolated from fresh seafood samples in Iran. Diagn. Pathol. 2013, 8, 149. [Google Scholar] [CrossRef]
- Abdellrazeq, G.S.; Kamar, A.M.; El-Houshy, S.M. Molecular Characterization of Listeria Species Isolated from Frozen Fish. Alex. J. Vet. Sci. 2014, 40, 1–15. [Google Scholar] [CrossRef]
- Edris, A.M.; Amany, M.S.; Michael, A.F. Incidence of Listeria monocytogenes in fresh tilapia nilotica fish. Benha Vet. Med. J. 2014, 26, 120–126. [Google Scholar]
- Mashak, Z.; Banisharif, F.; Banisharif, G.; Reza Pourian, M.; Eskandari, S.; Seif, A.; Dehkordi, F.S.; Alavi, I. Prevalence of listeria species and serotyping of Listeria monocytogenes bacteria isolated from seafood samples. Egypt. J. Vet. Sci. 2021, 52, 1–9. [Google Scholar] [CrossRef]
- Tarazi, Y.; El-Sukhon, S.; Al-Rahbi, A.; Ismail, Z.B. Molecular characterization and in vivo pathogenicity study of Listeria monocytogenes isolated from fresh and frozen local and imported fish in Jordan. Open Vet. J. 2021, 11, 517–524. [Google Scholar] [CrossRef]
- Chou, C.H.; Silva, J.L.; Wang, C. Prevalence and typing of Listeria monocytogenes in raw catfish fillets. J. Food Prot. 2006, 69, 815–819. [Google Scholar] [CrossRef] [PubMed]
- El-Malek, A.M.A.; Ali, S.F.H.; Hassanein, R.; Mohamed, M.A.; Elsayh, K.I. Occurrence of Listeria species in meat, chicken products and human stools in Assiut city, Egypt with PCR use for rapid identification of Listeria monocytogenes. Vet. World 2010, 3, 353–359. [Google Scholar] [CrossRef]
- Osaili, T.M.; Alaboudi, A.R.; Nesiar, E.A. Prevalence of Listeria spp. and antibiotic susceptibility of Listeria monocytogenes isolated from raw chicken and ready-to-eat chicken products in Jordan. Food Control 2011, 22, 586–590. [Google Scholar] [CrossRef]
- Karakolev, R. Incidence of Listeria monocytogenes in beef, pork, raw-dried and raw-smoked sausages in Bulgaria. Food Control 2009, 20, 953–955. [Google Scholar] [CrossRef]
- Sırıken, B.; Pamuk, Ş.; Özakın, C.; Gedikoglu, S.; Eyigör, M. A note on the incidences of Salmonella spp., Listeria spp. and Escherichia coli O157: H7 serotypes in Turkish sausage (Soudjouck). Meat Sci. 2006, 72, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Molla, B.; Yilma, R.; Alemayehu, D. Listeria monocytogenes and other Listeria species in retail meat and milk products in Addis Ababa, Ethiopia. Ethiop. J. Health Dev. 2004, 18, 208–212. [Google Scholar] [CrossRef]
- Gebretsadik, S.; Kassa, T.; Alemayehu, H.; Huruy, K.; Kebede, N. Isolation and characterization of Listeria monocytogenes and other Listeria species in foods of animal origin in Addis Ababa, Ethiopia. J. Infect. Public Health 2011, 4, 22–29. [Google Scholar] [CrossRef]
- Eruteya, O.C.; Odunfa, S.A.; Lahor, J. Listeria spp. in raw cow and goat meat in Port Harcourt, Nigeria. Br. Biotechnol. J. 2014, 4, 204–214. [Google Scholar] [CrossRef]
- Ismaiel, A.A.R.; Ali, A.E.S.; Enan, G. Incidence of Listeria in Egyptian meat and dairy samples. Food Sci. Biotechnol. 2014, 23, 179–185. [Google Scholar] [CrossRef]
- Ristori, C.A.; Rowlands, R.E.G.; Martins, C.G.; Barbosa, M.L.; Yoshida, J.T.; de Melo Franco, B.D. Prevalence and populations of Listeria monocytogenes in meat products retailed in Sao Paulo, Brazil. Foodborne Pathog. Dis. 2014, 11, 969–973. [Google Scholar] [CrossRef]
- Onyilokwu, S.A.; Lawan, F.A.; Hambali, I.U.; Mailafiya, S.; Adamu, N.B.; Atsanda, N.N.; Jauro, S. Phenotypic Characterisation and Distribution Pattern of Listeria Species Isolated from Food Samples Retailed In Markets and Central Abattoir in Maiduguri, Nigeria. Alex. J. Vet. Sci. 2016, 51, 122. [Google Scholar]
- Reda, W.W.; Abdel-Moein, K.; Hegazi, A.; Mohamed, Y.; Abdel-Razik, K. Listeria monocytogenes: An emerging food-borne pathogen and its public health implications. J. Infect. Dev. Ctries. 2016, 10, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Arslan, S.; Baytur, S. Prevalence and antimicrobial resistance of Listeria species and subtyping and virulence factors of Listeria monocytogenes from retail meat. J. Food Saf. 2019, 39, e12578. [Google Scholar] [CrossRef]
- Omogbai, B.A.; Esokpunwu, D.E. Molecular Characterization and Antibiotic Resistance Patterns of Listeria Species in Frozen Beef and Chicken Sold in Benin City, Nigeria. Food Appl. Biosci. J. 2019, 7, 68–86. [Google Scholar]
- Chuku, A.; Obande, G.A.; Eya, S.B. Listerial contamination of raw beef and chevon in north-central Nigeria. IMC J. Med. Sci. 2019, 13, 1–8. [Google Scholar] [CrossRef][Green Version]
- Gebremedhin, E.Z.; Hirpa, G.; Borana, B.M.; Sarba, E.J.; Marami, L.M.; Tadese, N.D.; Ambecha, H.A. Detection of Listeria species, factors associated, and antibiogram of Listeria monocytogenes in beef at abattoirs, butchers, and restaurants of Ambo and Holeta Towns, Ethiopia. Infect. Drug Resist. 2021, 14, 1493–1504. [Google Scholar] [CrossRef]
- Okorie-Kanu, O.J.; Anyanwu, M.U.; Ezenduka, E.V.; Mgbeahuruike, A.C.; Okorie-Kanu, C.O.; Ugwuijem, E.E.; Idogwu, M.N.; Anyaoha, C.O.; Majesti-Alugakberie, O.L.; Vidal, R.O.; et al. Occurrence and antibiogram of Listeria species in raw pork, beef, and chicken meats marketed in Enugu State, Southeast Nigeria. Vet. World 2020, 13, 317. [Google Scholar] [CrossRef]
- Al-Humam, N.A.; Reda, L.; Mohamed, R.E.; El-Ghareeb, W.R.; Darwish, W.S.; Ibrahim, N.A. Prevalence and Antibiogram of Listeria monocytogenes in Retailed Buffalo Raw Meat and Mince with a Protection Trial Using Nisin, and Gingerol. Buffalo Bull. 2021, 40, 47–57. [Google Scholar]
- Alonso-Hernando, A.; Prieto, M.; García-Fernández, C.; Alonso-Calleja, C.; Capita, R. Increase over time in the prevalence of multiple antibiotic resistance among isolates of Listeria monocytogenes from poultry in Spain. Food Control 2012, 23, 37–41. [Google Scholar] [CrossRef]
- Fallah, A.A.; Saei-Dehkordi, S.S.; Rahnama, M.; Tahmasby, H.; Mahzounieh, M. Prevalence and antimicrobial resistance patterns of Listeria species isolated from poultry products marketed in Iran. Food Control 2012, 28, 327–332. [Google Scholar] [CrossRef]
- Daniel, S.T.; Umeh, E.U.; Iheukwumere, C.C. Contamination and antibiotic susceptibility profile of Listeria species in frozen and fresh chicken sold in Makurdi, Nigeria. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 617–623. [Google Scholar]
- Dahshan, H.; Merwad, A.M.A.; Mohamed, T.S. Listeria species in broiler poultry farms: Potential public health hazards. J. Microbiol. Biotechnol. 2016, 26, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Antoniollo, P.C.; Bandeira, F.D.S.; Jantzen, M.M.; Duval, E.H.; Silva, W.P.D. Prevalence of Listeria spp. in feces and carcasses at a lamb packing plant in Brazil. J. Food Prot. 2003, 66, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Pesavento, G.; Ducci, B.; Nieri, D.; Comodo, N.; Nostro, A.L. Prevalence and antibiotic susceptibility of Listeria spp. isolated from raw meat and retail foods. Food Control 2010, 21, 708–713. [Google Scholar] [CrossRef]
- Al-Nabulsi, A.A.; Osaili, T.M.; Awad, A.A.; Olaimat, A.N.; Shaker, R.R.; Holley, R.A. Occurrence and antibiotic susceptibility of Listeria monocytogenes isolated from raw and processed meat products in Amman, Jordan. CyTA J. Food 2015, 13, 346–352. [Google Scholar] [CrossRef]
- Garedew, L.; Taddese, A.; Biru, T.; Nigatu, S.; Kebede, E.; Ejo, M.; Fikru, A.; Birhanu, T. Prevalence and antimicrobial susceptibility profile of Listeria species from ready-to-eat foods of animal origin in Gondar Town, Ethiopia. BMC Microbiol. 2015, 15, 100. [Google Scholar] [CrossRef]
- Doijad, S.P.; Vaidya, V.; Kalekar, S.; Rodrigues, J.; D’Costa, D.; Boshle, S.N.; Barbuddhe, S.B. Isolation and characterization of Listeria species from raw and processed meats. J. Vet. Pub. Health 2010, 8, 83–88. [Google Scholar]
- Soriano, J.M.; Rico, H.; Molto, J.C.; Manes, J. Listeria species in raw and ready-to-eat foods from restaurants. J. Food Prot. 2001, 64, 551–553. [Google Scholar] [CrossRef]
- Conficoni, D.; Santagiuliana, M.; Marchesan, M.; Franceschini, F.; Catellani, P.; Ferioli, M.; Giaccone, V. Distribution of Listeria spp. on Carcasses of Regularly Slaughtered Swine for Italian Dry Cured Ham. J. Food Prot. 2019, 82, 1104–1109. [Google Scholar] [CrossRef]
- Rodriguez-Calleja, J.M.; Garcia-Lopez, I.; Garcia-Lopez, M.L.; Santos, J.A.; Otero, A. Rabbit meat as a source of bacterial foodborne pathogens. J. Food Prot. 2006, 69, 1106–1112. [Google Scholar] [CrossRef]
- El-Gohary, A.H.; Mohamed, A.A.; Ramadan, H.H.; Abuhatab, E.A. Zoonotic and Molecular Aspects of Listeria Species Isolated from Some Farm Animals at Dakahlia Province in Egypt. Alex. J. Vet. Sci. 2018, 58, 208. [Google Scholar]
- Haggag, Y.N.; Nossair, M.A.; Shehab, S.A. Is Raw Milk Still Vehicle for Transmitting Listeria species To Pregnant Women? Alex. J. Vet. Sci. 2019, 61, 67–73. [Google Scholar] [CrossRef]
- Vazquez-Salinas, C.; Rodas-Suarez, O.; Quinones-Ramirez, E.I. Occurrence of Listeria species in raw milk in farms on the outskirts of Mexico City. Food Microbiol. 2001, 18, 177–181. [Google Scholar]
- Omar, S.S.; Dababneh, B.F.; Qatatsheh, A.; Abu-Romman, S.; Hawari, A.D.; Aladaileh, S. The incidence of Listeria species and other indicator bacteria in some traditional foods sold in Karak city, Jordan. J. Food Agric. Environ. 2011, 9, 79–81. [Google Scholar]
- Yakubu, Y.; Salihu, M.D.; Faleke, O.O.; Abubakar, M.B.; Junaidu, A.U.; Magaji, A.A.; Gulumbe, L.M.; Aliyu, R.M. Prevalence and antibiotic susceptibility of Listeria monocytogenes in raw milk from cattle herds within Sokoto Metropolis, Nigeria. Sokoto J. Vet. Sci. 2012, 10, 13–17. [Google Scholar] [CrossRef]
- Al-Mariri, A.; Younes, A.; Ramadan, L. Prevalence of Listeria spp. in raw milk in Syria. Bulg. J. Vet. Med. 2013, 16, 112–122. [Google Scholar]
- Rahimi, E.; Momtaz, H.; Behzadnia, A.; Baghbadorani, Z.T. Incidence of Listeria species in bovine, ovine, caprine, camel and water buffalo milk using cultural method and the PCR assay. Asian Pac. J. Trop. Dis. 2014, 4, 50–53. [Google Scholar] [CrossRef]
- Acaröz, U.; Acaröz-Arslan, D.; Recep, K.A.R.A.; Zemheri, F.; Gürler, Z. Determination of Listeria species in water buffalo and cow milk obtained from Afyonkarahisar province. Kocatepe Vet. J. 2017, 10, 264–268. [Google Scholar]
- EL-Naenaeey, E.S.; Abdelwahab, A.; Merwad, A.; Abdou, H. Prevalence of Listeria Species in Dairy Cows and Pregnant Women with Reference to Virulotyping of Listeria monocytogenes in Egypt. Zagazig Vet. J. 2019, 47, 248–258. [Google Scholar] [CrossRef]
- Babacan, O. Determination of the presence and antibiotic resistance of Listeria species and aerobic mesophilic bacteria count of cow milks. Veteriner Hekimler Derneği Dergisi 2021, 92, 16–23. [Google Scholar]
- Rodriguez, J.L.; Gava, P.; Medina, M.; Nuñez, M. Incidence of Listeria monocytogenes and other Listeria spp. in ewes’ raw milk. J. Food Prot. 1994, 57, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Vitas, A.I. Occurrence of Listeria monocytogenes in fresh and processed foods in Navarra (Spain). Int. J. Food Microbiol. 2004, 90, 349–356. [Google Scholar] [CrossRef]
- Lianou, D.T.; Skoulakis, A.; Michael, C.M.; Katsarou, E.I.; Chatzopoulos, D.C.; Solomakos, N.; Tsilipounidaki, K.; Florou, Z.; Cripps, P.J.; Katsafadou, A.I.; et al. Isolation of Listeria ivanovii from Bulk-Tank Milk of Sheep and Goat Farms-From Clinical Work to Bioinformatics Studies: Prevalence, Association with Milk Quality, Antibiotic Susceptibility, Predictors, Whole Genome Sequence and Phylogenetic Relationships. Biology 2022, 11, 871. [Google Scholar] [CrossRef]
- Gaya, P.; Saralegui, C.; Medina, M.; Nunez, M. Occurrence of Listeria monocytogenes and other Listeria spp. in raw caprine milk. J. Dairy Sci. 1996, 79, 1936–1941. [Google Scholar] [CrossRef]
- Baher, W.; Shalaby, M.; Abdelghfar, S. Prevalence of multidrug-resistant Listeria monocytogenes in retailed goat meat and offal. Damanhour J. Vet. Sci. 2021, 7, 19–22. [Google Scholar] [CrossRef]
- Hyera, E.; Msalya, G.; Karimuribo, E.D.; Kurwijila, L.R.; Alonso, S.; Roesel, K.; Grace, D. Isolation and identification of Listeria species along the milk value chain in one region of Tanzania. In Proceedings of the First Joint AITVM—STVM Conference, Berlin, Germany, 4–8 September 2016. [Google Scholar]
- Usman, U.B.; Kwaga, J.K.P.; Kabir, J.; Olonitola, O.S. Isolation and antimicrobial susceptibility of Listeria monocytogenes from raw milk and milk products in Northern Kaduna State, Nigeria. J. Appl. Environ. Microbiol. 2016, 4, 46–54. [Google Scholar]
- El Hag, M.M.; El Zubeir, I.E.M.; Mustafa, N.E. Prevalence of Listeria species in dairy farms in Khartoum State (Sudan). Food Control 2021, 123, 107699. [Google Scholar] [CrossRef]
- Borena, B.M.; Dilgasa, L.; Gebremedhin, E.Z.; Sarba, E.J.; Marami, L.M.; Kelbesa, K.A.; Tadese, N.D. Listeria Species Occurrence and Associated Risk Factors and Antibiogram of Listeria monocytogenes in Milk and Milk Products in Ambo, Holeta, and Bako Towns, Oromia Regional State, Ethiopia. Vet. Med. Int. 2022, 2022, 5643478. [Google Scholar] [CrossRef]
- Eldaly, E.A.; Saleh, E.A.; Moustafa, A.H.; Atya, O. Prevalence Of Listeria Organisms In Meat And Some Meat Products. Zagazig Vet. J. 2013, 41, 57–68. [Google Scholar] [CrossRef]
- Aisha, B.M.; Kawo, A.H. Isolation of Listeria monocytogenes recovered from some ready-to-eat foods sold in Kano, North-Western Nigeria. Bayero J. Pure Appl. Sci. 2014, 7, 8–12. [Google Scholar] [CrossRef]
- Ndahi, M.D.; Kwaga, J.K.P.; Bello, M.; Kabir, J.; Umoh, V.J.; Yakubu, S.E.; Nok, A.J. Prevalence and antimicrobial susceptibility of Listeria monocytogenes and methicillin-resistant Staphylococcus aureus strains from raw meat and meat products in Zaria, Nigeria. Lett. Appl. Microbiol. 2014, 58, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, H.; Karmi, M.; Maky, M. Occurrence and Characterization of Listeria Species Isolated from Processed Meat in Qena, Egypt. Zagazig Vet. J. 2019, 47, 267–276. [Google Scholar] [CrossRef]
- Nyenje, M.E.; Odjadjare, C.E.; Tanih, N.F.; Green, E.; Ndip, R.N. Foodborne pathogens recovered from ready-to-eat foods from roadside cafeterias and retail outlets in Alice, Eastern Cape Province, South Africa: Public health implications. Int. J. Environ. Res. Public Health 2012, 9, 2608–2619. [Google Scholar] [CrossRef] [PubMed]
- Büyükyörük, S.; Beyaz, D.; Göksoy, E.Ö.; Filiz, K.Ö.K.; Kocak, P. Microbiological evaluation of ready-to-eat sandwiches served near hospitals and schools. Ank. Üniversitesi Vet. Fakültesi Derg. 2014, 61, 193–198. [Google Scholar]
- İşleyici, Ö.; Sancak, Y.C.; Tuncay, R.M.; Atlan, M. Presence of Listeria species in ready-made meatballs offered by sale under freezing or cooling preservation. Ank. Univ. Vet. Fakültesi Derg. 2019, 66, 81–288. [Google Scholar] [CrossRef]
- Sotohy, E.M.; Abd EL-Malek, A. Assessment of microbiological quality of ready to eat meat sandwiches in new valley governorate. Int. J. Food Sci. Nutr. Eng. 2019, 4, 186–192. [Google Scholar]
- Kovačević, M.; Burazin, J.; Pavlović, H.; Kopjar, M.; Piližota, V. Prevalence and level of Listeria monocytogenes and other Listeria sp. in ready-to-eat minimally processed and refrigerated vegetables. World J. Microbiol. Biotechnol. 2013, 29, 707–712. [Google Scholar] [CrossRef]
- Alsheikh, A.D.I.; Mohammed, G.E.; Abdalla, M.A. Isolation and identification of Listeria monocytogenes from retail broiler chicken ready to eat meat products in Sudan. Int. J. Anim. Vet. Adv. 2013, 5, 9–14. [Google Scholar] [CrossRef]
- Ripabelli, G.; Sammarco, M.L.; Fanelli, I.; Grasso, G.M. Prevalenza di Campylobacter, Salmonella, Vibrio, Yersinia enterocolitica, Listeria ed Escherichia coli in vegetali freschi del commercio. L’Igiene Mod. 2002, 118, 137–147. [Google Scholar]
- Francis, G.A.; O’Beirne, D.A.V.I.D. Isolation and pulsed-field gel electrophoresis typing of Listeria monocytogenes from modified atmosphere packaged fresh-cut vegetables collected in Ireland. J. Food Prot. 2006, 69, 2524–2528. [Google Scholar] [CrossRef]
- Stonsaovapak, S.; Boonyaratanakornkit, M. Prevalence and antimicrobial resistance of Listeria species in food products in Bangkok, Thailand. J. Food Saf. 2010, 30, 154–161. [Google Scholar] [CrossRef]
- Bouayad, L.; Hamdi, T.M. Prevalence of Listeria spp. in ready to eat foods (RTE) from Algiers (Algeria). Food Control 2012, 23, 397–399. [Google Scholar] [CrossRef]
- Wang, F.I.; Chern, M.K.; Li, C.W.; Yan, M.; Hsieh, Y.H. Prevalence and antibiotic resistance of Listeria species in food products in Taipei, Taiwan. Afr. J. Microbiol. Res. 2012, 6, 4702–4706. [Google Scholar]
- Ebakota, D.O.; Abiodun, O.A.; Nosa, O.O. Prevalence of antibiotics resistant Listeria monocytogenes strains in Nigerian ready-to-eat foods. Food Saf. 2018, 6, 118–125. [Google Scholar] [CrossRef]
- Camellini, S.; Iseppi, R.; Condò, C.; Messi, P. Ready-to-Eat Sandwiches as Source of Pathogens Endowed with Antibiotic Resistance and Other Virulence Factors. Appl. Sci. 2021, 11, 7177. [Google Scholar] [CrossRef]
- Ramírez Mérida, L.G.; Morón de Salim, A.; Alfieri Graterol, A.Y.; Gamboa, O. Frecuencia de Listeria monocytogenes en muestras de tomates (Lycopersicum esculentum) y cilantro (Coriandrum sativum) frescos en tres supermercados de Valencia, Venezuela. Arch. Latinoam. Nutr. 2009, 59, 318–324. [Google Scholar]
- Mawak, J.D.; Dashen, M.M.; Idolo, A.J.; Chukwu, O.O.C. Occurrence of Listeria monocytogenes in irrigation water and vegetable at Jos, Plateau State, Nigeria. Int. J. Trop. Agric. Food Syst. 2009, 3. [Google Scholar]
- Hunter, R. Observations on Listeria monocytogenes type 5 (Iwanov) isolated in New Zealand. Med. Lab. Technol. 1973, 30, 51–56. [Google Scholar]
- Dennis, S.M. Perinatal lamb mortality in western Australia. Aust. Vet. J. 1975, 51, 75–79. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Diriba, K.; Awulachew, E.; Diribsa, K. The prevalence of Listeria species in different food items of animal and plant origin in Ethiopia: A systematic review and meta-analysis. Eur. J. Med. Res. 2021, 26, 60. [Google Scholar] [CrossRef] [PubMed]
- Castrica, M.; Menchetti, L.; Panseri, S.; Cami, M.; Balzaretti, C.M. When Pet Snacks Look Like Children’s Toys! The Potential Role of Pet Snacks in Transmission of Bacterial Zoonotic Pathogens in the Household. Foodborne Pathog. Dis. 2021, 18, 56–62. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Detection of Listeria monocytogenes in foods and environmental samples, and enumeration of Listeria monocytogenes in foods. In Bacteriological Analytical Manual; 2017; Chapter 10. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-detection-and-enumeration-Listeria-monocytogenes (accessed on 27 July 2022).
- International Organization for Standardization. Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Detection and Enumeration of Listeria Monocytogenes and Listeria spp. ISO 11290-1:2017; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- U.S. Department of Agriculture. Food Safety and Inspection Service. Isolation and identification of Listeria monocytogenes from red meat, poultry, ready-to-eat Siluriformes (fish) and egg products, and environmental samples, method 8.11. In Microbiology Laboratory Guidebook; U.S. Department of Agriculture: Washington, DC, USA, 2019. Available online: https://www.fsis.usda.gov/news-events/publications/microbiology-laboratory-guidebook (accessed on 27 July 2022).
- Supanivatin, P.; Kosonpisit, S.; Liamkaew, R.; Saeteaw, N.; Thipayarat, A. Inhibitory effects of Listeria selective enrichment media on growth characteristics of L. ivanovii. Procedia Eng. 2012, 32, 112–118. [Google Scholar] [CrossRef][Green Version]
- Liu, D.; Ainsworth, A.J.; Austin, F.W.; Lawrence, M.L. PCR detection of a putative N-acetylmuramidase gene from Listeria ivanovii facilitates its rapid identification. Vet. Microbiol. 2004, 101, 83–89. [Google Scholar] [CrossRef]
- Liu, D.; Lawrence, M.L.; Austin, F.W.; Ainsworth, A.J. A multiplex PCR for species- and virulence-specific determination of Listeria monocytogenes. J. Microbiol. Methods 2007, 71, 133–140. [Google Scholar] [CrossRef]
- Li, F.; Ye, Q.; Chen, M.; Zhang, J.; Xue, L.; Wang, J.; Wu, S.; Zeng, H.; Gu, Q.; Zhang, Y.; et al. Multiplex PCR for the Identification of Pathogenic Listeria in Flammulina velutipes Plant Based on Novel Specific Targets Revealed by Pan-Genome Analysis. Front. Microbiol. 2021, 11, 634255. [Google Scholar] [CrossRef]
- Mao, Y.; Huang, X.; Xiong, S.; Xu, H.; Aguilar, Z.P.; Xiong, Y. Large-volume immunomagnetic separation combined with multiplex PCR assay for simultaneous detection of Listeria monocytogenes and Listeria ivanovii in lettuce. Food Control 2016, 59, 601–608. [Google Scholar] [CrossRef]
- Xiao, F.; Bai, X.; Wang, K.; Sun, Y.; Xu, H. Rapid-Response Magnetic Enrichment Strategy for Significantly Improving Sensitivity of Multiplex PCR Analysis of Pathogenic Listeria Species. Appl. Sci. 2022, 12, 6415. [Google Scholar] [CrossRef]
- Rodríguez-Lázaro, D.; López-Enríquez, L.; Hernández, M. smcL as a novel diagnostic marker for quantitative detection of Listeria ivanovii in biological samples. J. Appl. Microbiol. 2010, 109, 863–872. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Xu, H.; Dai, H.; Meng, S.; Ye, C. Rapid and sensitive detection of Listeria ivanovii by loop-mediated isothermal amplification of the smcL gene. PLoS ONE 2014, 9, e115868. [Google Scholar] [CrossRef]
- Barbuddhe, S.B.; Maier, T.; Schwarz, G.; Kostrzewa, M.; Domann, E.; Chakraborty, T.; Hain, T. Rapid identification and typing of Listeria species using matrix assisted laser desorption ionization-time of flight mass spectrometry. Appl. Environ. Microbiol. 2008, 74, 5402–5407. [Google Scholar] [CrossRef] [PubMed]


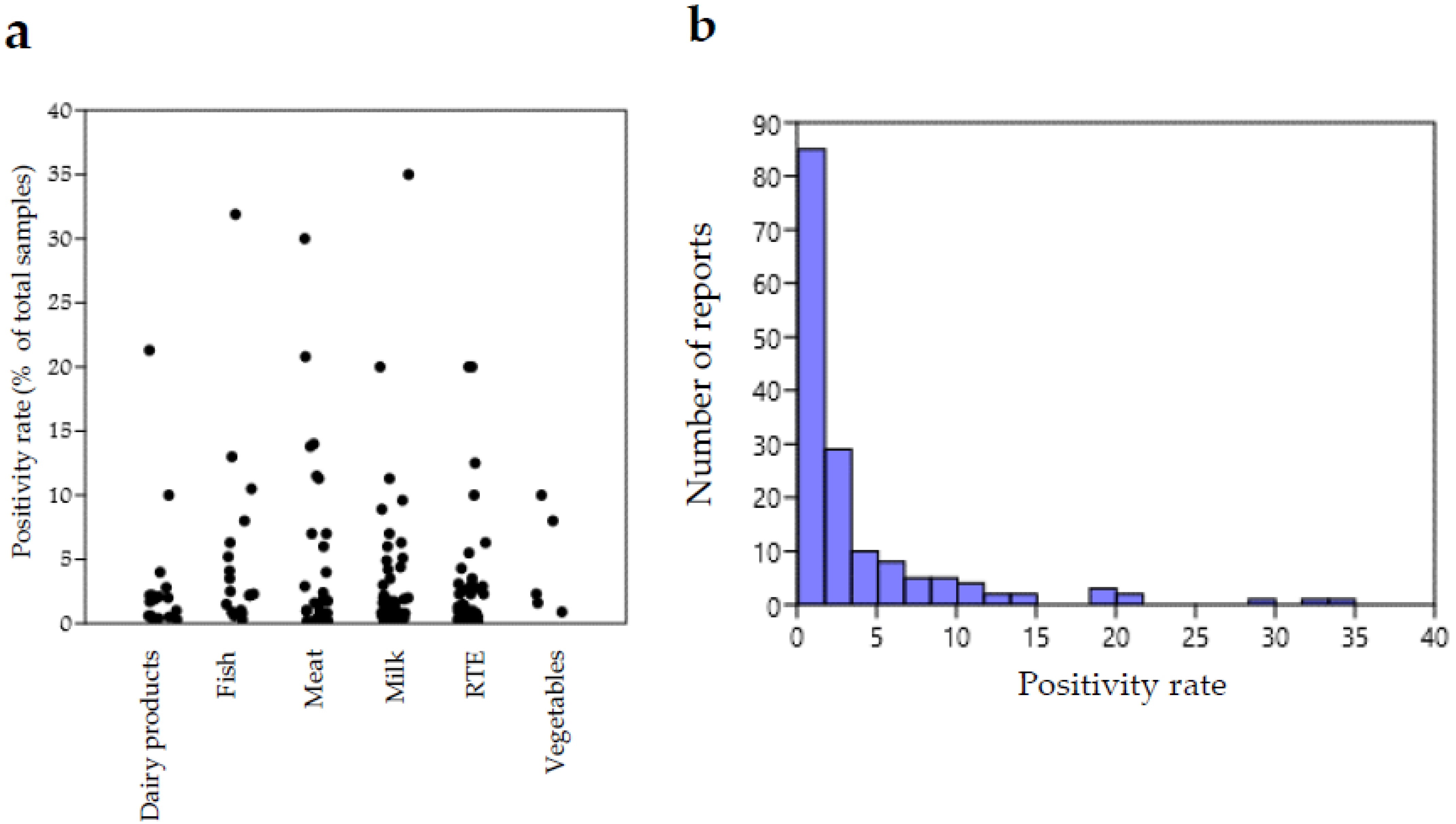{kind=link}
| Food Matrix | Number of Samples Positive for L. ivanovii, Positivity Rate (%) on All Analyzed Samples for Each Report, Country and Reference |
|---|---|
| Dairy products | |
| Butter | 1 (0.4) Egypt [50], 1 (0.3) Turkey [51] |
| Cheese | 3 (0.6) Portugal [52], 3 (1.9) Turkey [53], 32 (2.2) Colombia [54], 3 (2.1) Turkey [55], 2 (2.8) Italy [56], 2 (0.2) Ethiopia [57], 1 (1.7) Turkey [58], 3 (10) Venezuela [59], 1 (0.4) Turkey [60], 2 (2.7) Turkey [61], 14 (4) Jordan [62], 1 (0.4) Iraq [63], 1 (0.4) Egypt [50], 6 (2.2) Turkey [64], 2 (0.6) Turkey [51], 6 (0.5) Turkey [65], 5 (4.0) Turkey [66], 4 (2.0) Turkey [67], 2 (1.0) Libya [68], 48 (21.3) Egypt [69] |
| Fish | |
| Conserved fish and seafood | 2 (5.2) India [70], 2 (0.6) Spain [71], 30 (31.9) Malaysia [72], 15 (13) Nigeria [73] |
| Fresh or frozen fish and seafood, | 2 (0.3) Japan [74], 4 (10.5) India [70], 3 (2.2) Costa Rica [75], 5 (2.3) Italy [76], 1 (1) Turkey [77], 7 (3.5) Iran [78], 3 (6.3) Greece [79], 3 (1) Iran [80], 2 (0.9) Egypt [81], 8 (8) Egypt [82], 8 (2.2) Iran [83], 21 (4.1) Jordan [84], 2 (0.8) USA [85], 3 (1.5) Libya [68] |
| Meat | |
| Conserved chicken | 2 (7) India [70], 1 (1) Egypt [86], 25 (20.8) Jordan [87] |
| Conserved pork | 4 (14) India [70], 2 (0.2) Bulgaria [88], 1 (1.0) Turkey [89] |
| Conserved meat (animal species not detailed) | 1 (0.2) Ireland [26] |
| Raw/frozen beef | 14 (13.8) Malaysia [72], 1 (0.3) Ethiopia [90], 2 (0.2) Bulgaria [89], 2 (0.2) Ethiopia [57], 1 (0.25) Ethiopia [91], 4 (1.7) Nigeria [92], 3 (1.6) Egypt [93], 1 (0.2) Brazil [94], 6 (1.8) Nigeria [95], 2 (0.8) Egypt [96], 5 (4.0) Turkey [97], 7 (2.9) Nigeria [98], 1 (0.5) Libya [68], 12 (11.5) Nigeria [99], 4 (0.9) Ethiopia [100], 4 (0.9) Nigeria [101] |
| Raw buffalo meat | 1 (1) Egypt [102] |
| Raw/frozen chicken | 2 (7) India [70], 6 (6) Egypt [86], 48 (30.0) Jordan [87], 1 (1.0) Spain [103], 10 (2.4) Iran [104], 2 (1.7) Nigeria [105], 9 (11.3) Egypt [106], 5 (2.0) Nigeria [98], 3 (0.6) Ethiopia [100], 3 (0.6) Nigeria [101], 4 (27) Egypt [41] |
| Raw/frozen lamb | 1 (1.4) Brazil [107], |
| Raw goat meat | 10 (9.6) Nigeria [99] |
| Raw/frozen meat (animal species not detailed) | 9 (8.9) Malaysia [72], 10 (1.7) Italy [108], 19 (7.0) Jordan [109], 2 (0.5) Ethiopia [110], 17 (5.1) Nigeria [95], 10 (2.2) Ethiopia [100], 1 (0.9) India [111] |
| Raw/frozen pork | 2 (1.8) Spain [112], 1 (0.1) Bulgaria [88], 17 (11.3) Italy [113] |
| Raw rabbit | 1 (2.0) Spain [114] |
| Milk | |
| Raw buffalo milk | 1 (1.6) Egypt [115], 2 (0.8) Egypt [116] |
| Raw cow milk | 2 (0.4) Portugal [51], 82 (6.3) Mexico [117], 7 (35.0) Jordan [118], 8 (4.2) Nigeria [119], 7 (0.9) Syria [120], 1 (0.5) Egypt [93], 1 (0.4) Iran [121], 1 (0.5) Turkey [122], 1 (1.6) Egypt [115], 12 (6) Egypt [123], 2 (0.8) Egypt [116], 3 (4.4) Turkey [124] |
| Raw ewe milk | 2 (0.2) Spain [125], 5 (0.9) Spain [126], 5 (0.6) Syria [120], 1 (0.4) Iran [121], 1 (1.6) Egypt [115], 1 (0.4) Egypt [116], 3 (0.9) Greece [127] |
| Raw goat milk | 3 (0.2) Spain [128], 4 (20) Egypt [129] |
| Raw zebu milk | 2 (1.9) Tanzania [130] |
| Raw milk (animal species not detailed) | 1 (0.6) Turkey [53], 1 (0.4) Italy [56], 1 (0.9) India [127], 1 (0.4) Turkey [60], 2 (0.8) Egypt [50], 27 (4.9) Nigeria [131], 5 (1.6) Turkey [51], 1 (0.9) Turkey [66], 3 (1.5) Turkey [67], 7 (3.5) Libya [68], 2 (1.7) Sudan [132], 6 (1.2) Ethiopia [133] |
| Raw buffalo milk | 1 (1.6) Egypt [115], 2 (0.8) Egypt [116] |
| Ready to eat (RTE) 1 food products | |
| Meat (animal species not detailed) | 1 (0.2) Portugal [52], 3 (0.7) Iran [104], 1 (0.4) Egypt [134], 12 (3.5) Nigeria [135], 19 (6.3) Nigeria [136], 54 (20) Jordan [109], 1 (0.4) Egypt [50], 12 (10) Egypt [137], 2 (1.0) Libya [68] |
| Beef | 6 (2.3) South Africa [138], 4 (1.4) Turkey [139], 8 (2.7) Egypt [140], 1 (0.8) Egypt [141] |
| Cabbages | 1 (1.0) Croatia [142], 9 (2.6) Nigeria [135], |
| Cheese | 1 (0.3) Turkey [139], |
| Chicken | 11 (4.3) South Africa [138], 52 (20.8) Sudan [143], |
| Lettuce | 3 (2.9) Spain [112], 1 (0.5) Italy [144], 3 (3.0) Croatia [142], 10 (2.9) Nigeria [135] |
| Potatoes | 6 (2.3) South Africa [138] |
| Rice | 6 (2.3) South Africa [138] |
| Bean sprouts | 1 (0.1) Ireland [145], |
| Turkey | 1 (0.3) Egypt [140] |
| Vegetables | 8 (3.1) South Africa [138] |
| Not detailed | 3 (0.8) Thailand [146], 20 (20.0) Jordan [118], 3 (1.3) Algeria [147], 4 (1) Taiwan [148], 1 (1.0) Croatia [142], 14 (5.5) South Africa [138], 6 (1.5) Nigeria [149], 3 (12.5) Italy [150] |
| Vegetables | |
| Coriander | 32 (1.6) Venezuela [151] |
| Leafy vegetables | 1 (10) Nigeria [152] |
| Lettuce | 1 (0.9) Spain [112] |
| Tomato | 16 (8.0) Venezuela [151] |
| Not detailed | 17 (2.3) Nigeria [95] |
| Other | |
| Liquid whole egg | 1 (0.25) Ethiopia [91] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, F.; Giaccone, V.; Colavita, G.; Amadoro, C.; Pomilio, F.; Catellani, P. Virulence Characteristics and Distribution of the Pathogen Listeria ivanovii in the Environment and in Food. Microorganisms 2022, 10, 1679. https://doi.org/10.3390/microorganisms10081679
Rossi F, Giaccone V, Colavita G, Amadoro C, Pomilio F, Catellani P. Virulence Characteristics and Distribution of the Pathogen Listeria ivanovii in the Environment and in Food. Microorganisms. 2022; 10(8):1679. https://doi.org/10.3390/microorganisms10081679
Chicago/Turabian StyleRossi, Franca, Valerio Giaccone, Giampaolo Colavita, Carmela Amadoro, Francesco Pomilio, and Paolo Catellani. 2022. "Virulence Characteristics and Distribution of the Pathogen Listeria ivanovii in the Environment and in Food" Microorganisms 10, no. 8: 1679. https://doi.org/10.3390/microorganisms10081679
APA StyleRossi, F., Giaccone, V., Colavita, G., Amadoro, C., Pomilio, F., & Catellani, P. (2022). Virulence Characteristics and Distribution of the Pathogen Listeria ivanovii in the Environment and in Food. Microorganisms, 10(8), 1679. https://doi.org/10.3390/microorganisms10081679