
Isolation of Aquatic Plant Growth-Promoting Bacteria for the Floating Plant Duckweed (Lemna minor)

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Cultivation of Rhizobacteria from Aquatic Plants
2.2. Screening of Aquatic PGPB for Duckweeds
2.3. Evaluation of Symbiotic Effects in the Duckweed/PGPB Co-Culture
2.4. Phylogenetic Identification of PGPB Strains
2.5. Assays on Plant Growth-Promoting Properties and Motility of PGPB Strains
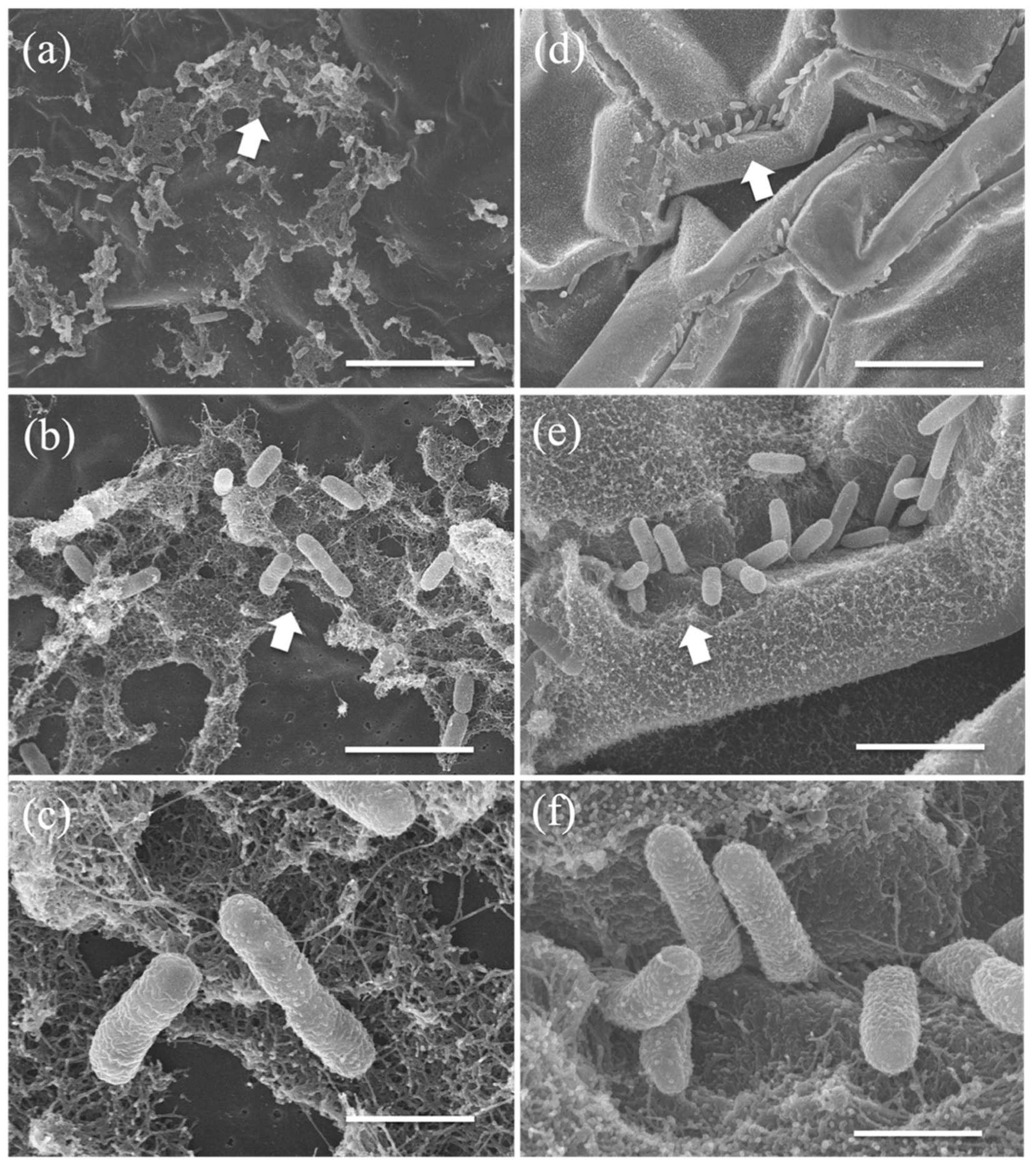
2.6. Microscopic Observations of PGPB Strains on Duckweed Surface
2.7. Statistical Analysis
3. Results and Discussion
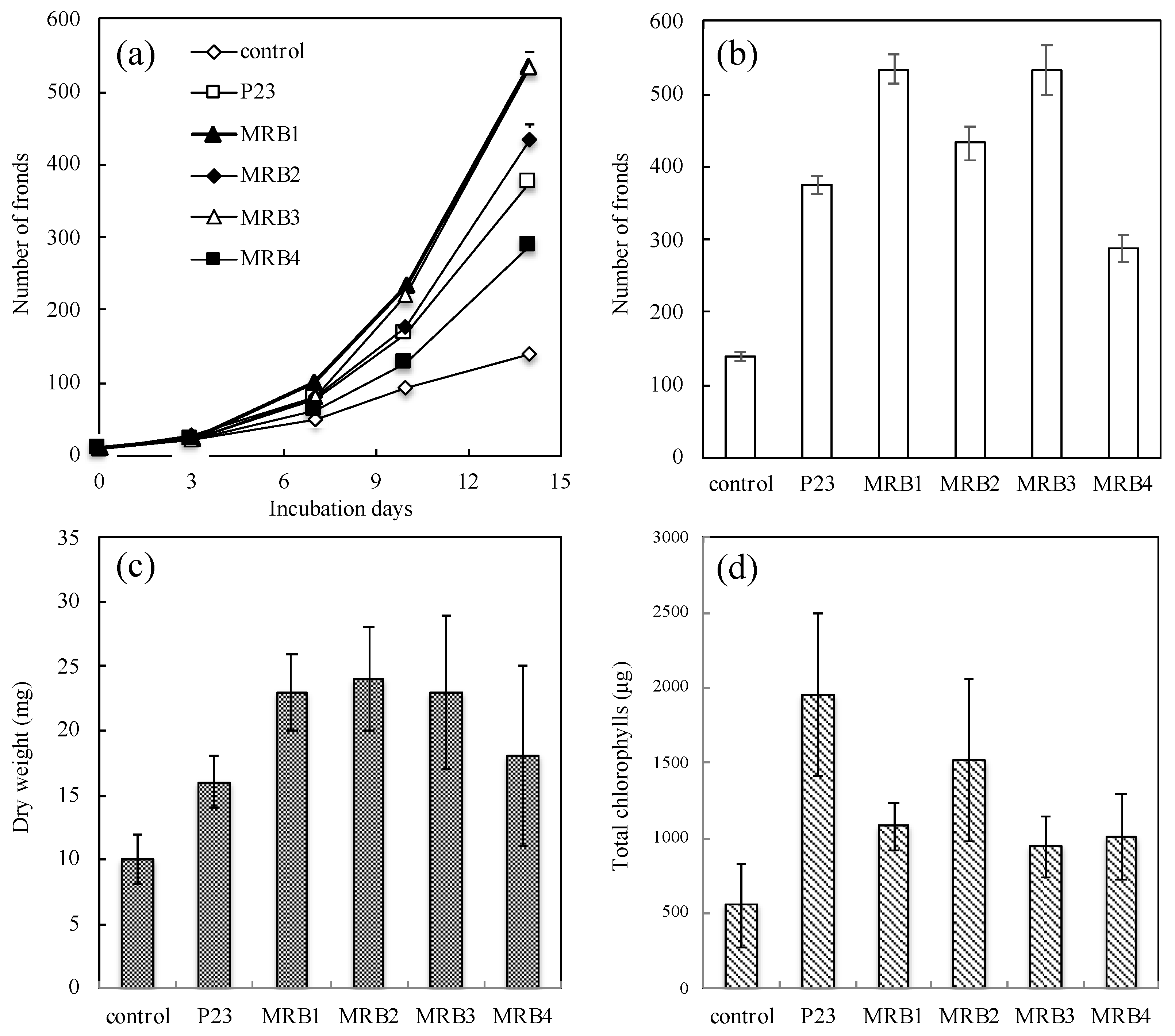
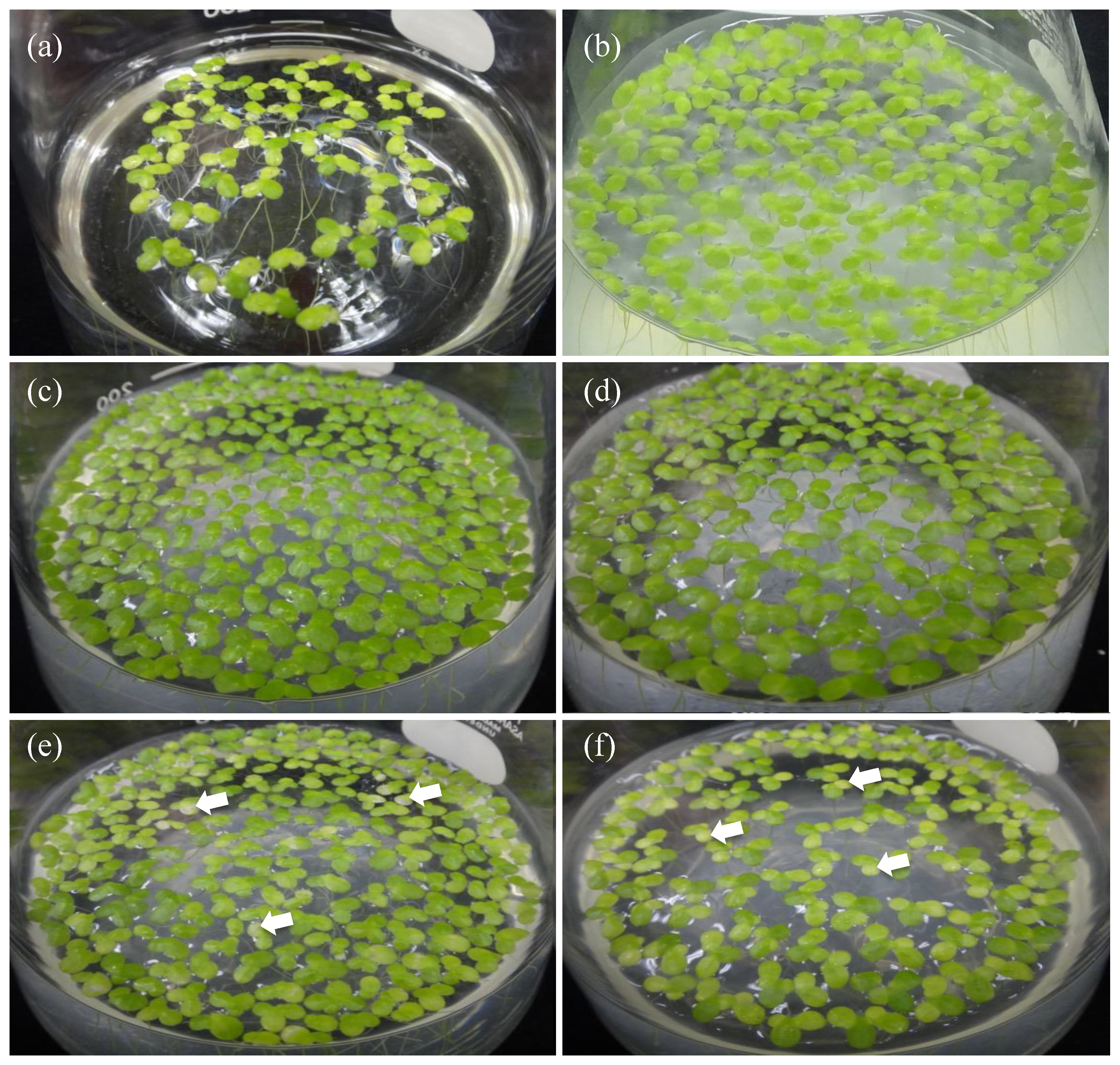
3.1. Duckweed Growth Promotion by Aquatic PGPB
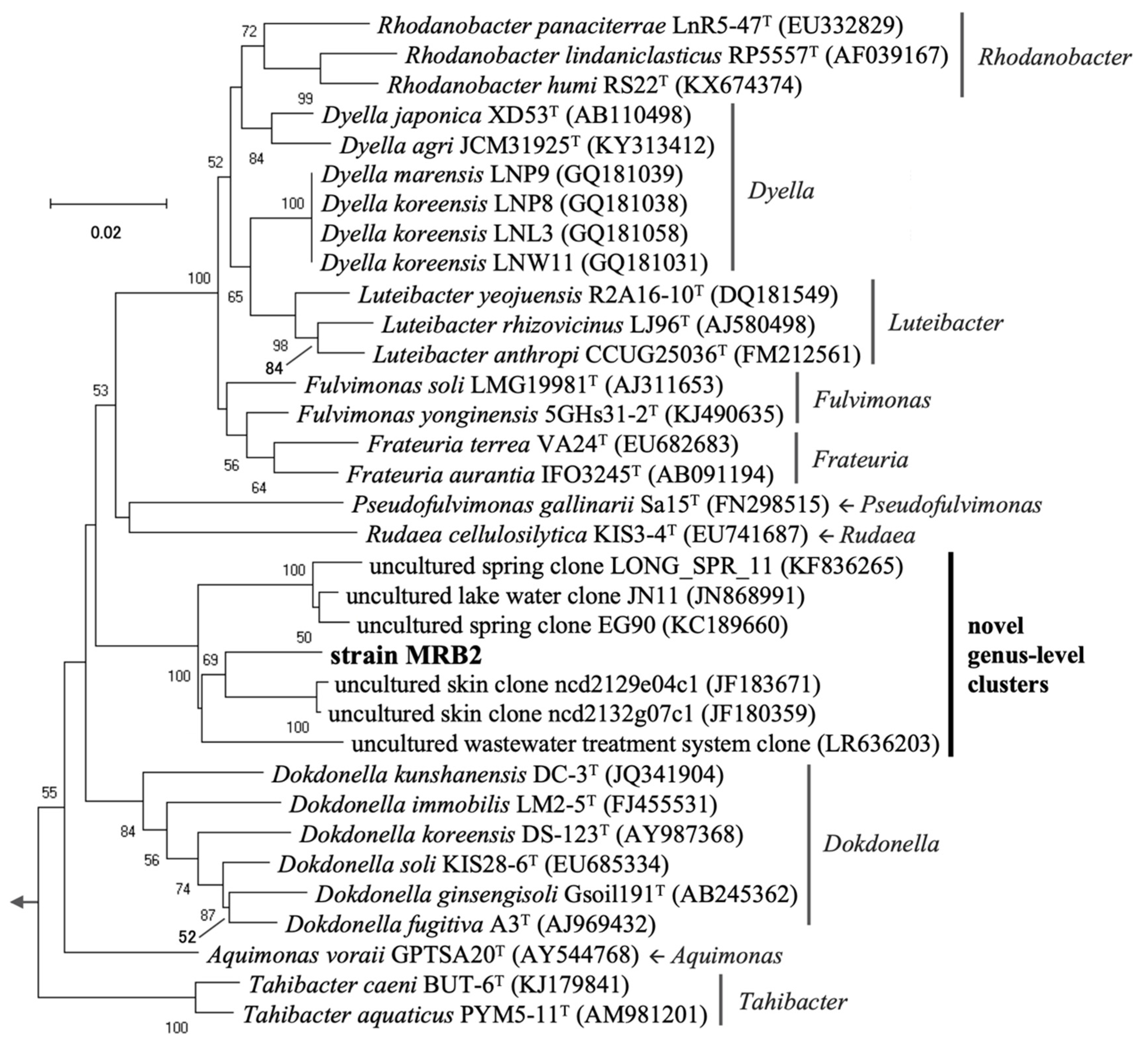
3.2. Phylogenetic Identification of Aquatic PGPB
3.3. Characterization of Plant Growth-Promoting Properties

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon (Phylum or Class) | Aquatic PGPB Strains | IAA | Siderophore | Phosphate Solubilization | Nitrogen Fixation | Motility | Reference |
|---|---|---|---|---|---|---|---|
| Acidobacteria | Paludibaculum sp. F-183 | + | − | − | nd | nd | [24,82,83] |
| Luteitalea sp. TBR-22 | − | − | − | nd | nd | ||
| Alphaproteobacteria | Bradyrhizobium sp. MRB4 | + | + | − | − * | + | this study |
| Ensifer sp. SP4 | − | + | − | − | nd | [23] | |
| Betaproteobacteria | Pelomonas sp. MRB1 | + | − | − | + | + | this study |
| Pelomonas sp. MRB3 | + | + | − | + | + | this study | |
| Aquitalea magnusonii H3 | + | + | + | nd | + | [22,84] | |
| Gammaproteobacteria | Rhodanobacteraceae sp. MRB2 | + | + | − | − | − | this study |
| Acinetobacter calcoaceticus P23 | − | + | + | nd | + † | [18,85] | |
| Pseudomonas sp. Ps6 | + | − | +/− | nd | + † | [18] | |
| Pseudomonas otitidis M12 | + | + | + | nd | nd | [22] | |
| Firmicutes | Bacillus amyloliquefaciens FZB42 | + | + | + | nd | + | [20,86,87,88] |
| Bacteroidetes | Chryseobacterium sp. 27AL | + | + | − | − | nd | [85] |
| Chryseobacterium sp. 29AL | + | + | − | − | nd |
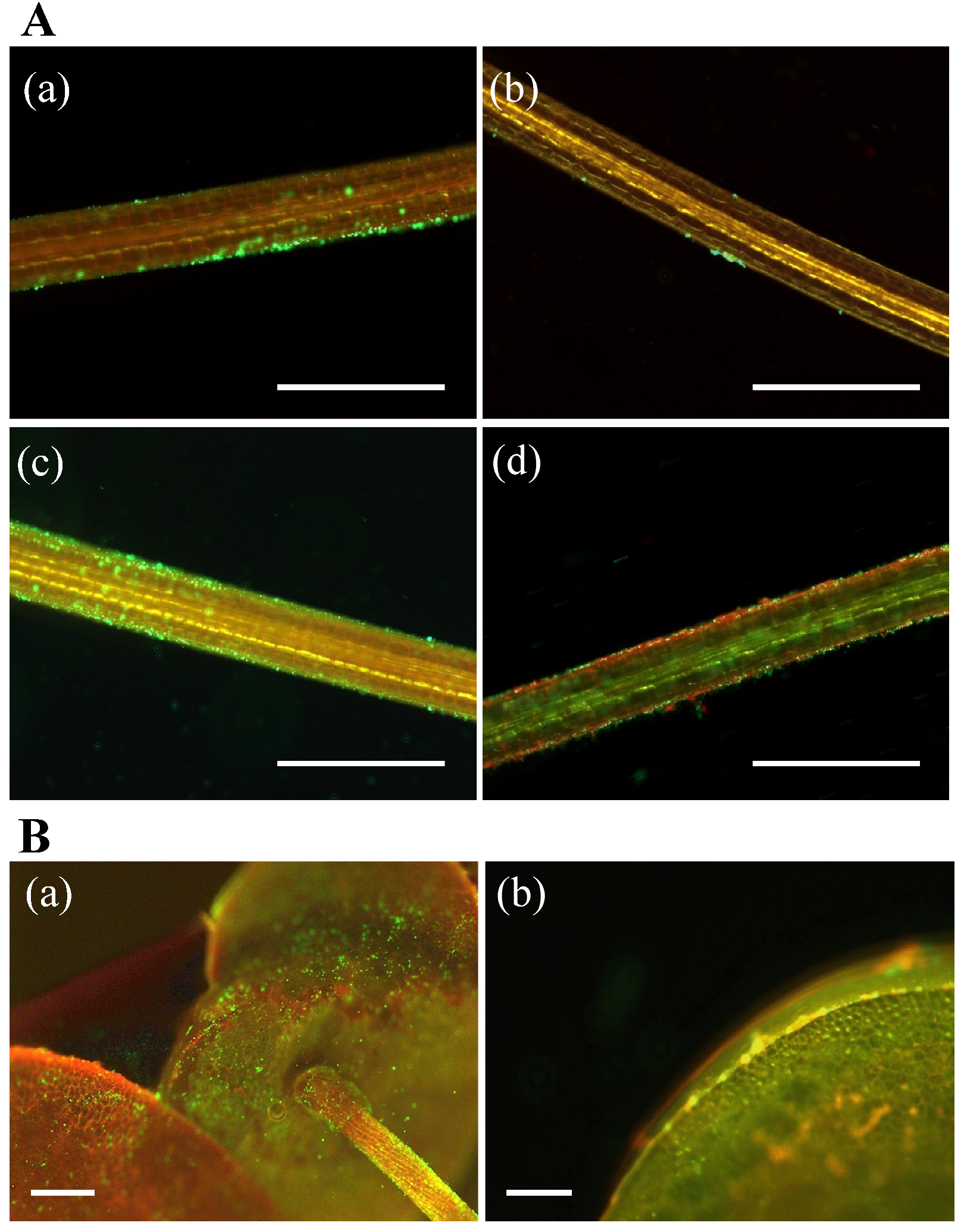
3.4. PGPB Adhesion and Colonization of Duckweed Surfaces
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.S.; Zaidi, A.; Musarrat, J. (Eds.) Microbial Strategies for Crop Improvement; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-3-642-01978-4. [Google Scholar]
- Turner, T.R.; James, E.K.; Poole, P.S. The Plant Microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y. Inoculants of Plant Growth-Promoting Bacteria for Use in Agriculture. Biotechnol. Adv. 1998, 16, 729–770. [Google Scholar] [CrossRef]
- Lucy, M.; Reed, E.; Glick, B.R. Applications of Free Living Plant Growth-Promoting Rhizobacteria. Antonie Van Leeuwenhoek 2004, 86, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.N.; Jha, D.K. Plant Growth-Promoting Rhizobacteria (PGPR): Emergence in Agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Perez-Montaño, F.; Alías-Villegas, C.; Bellogín, R.A.; del Cerro, P.; Espuny, M.R.; Jiménez-Guerrero, I.; López-Baena, F.J.; Ollero, F.J.; Cubo, T. Plant Growth Promotion in Cereal and Leguminous Agricultural Important Plants: From Microorganism Capacities to Crop Production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef]
- Leng, R.A.; Stambolie, J.H.; Bell, R. Duckweed—A Potential High-protein Feed Resource for Domestic Animals and Fish. Livest. Res. Rural Dev. 1995, 7, 36. [Google Scholar]
- Fasakin, E.A.; Balogun, A.M.; Fasuru, B.E. Use of Duckweed, Spirodela Polyrrhiza L. Schleiden, as a Protein Feedstuff in Practical Diets for Tilapia, Oreochromis Niloticus L. Aquac. Res. 1999, 30, 313–318. [Google Scholar] [CrossRef]
- Verma, R.; Suthar, S. Utility of Duckweeds as Source of Biomass Energy: A Review. Bioenerg. Res. 2015, 8, 1589–1597. [Google Scholar] [CrossRef]
- Yu, C.; Sun, C.; Yu, L.; Zhu, M.; Xu, H.; Zhao, J.; Ma, Y.; Zhou, G. Comparative Analysis of Duckweed Cultivation with Sewage Water and SH Media for Production of Fuel Ethanol. PLoS ONE 2014, 9, e115023. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Cheng, J.J. Growing Duckweed for Biofuel Production: A Review. Plant Biol. 2015, 17, 16–23. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, H.; Stomp, A.-M.; Cheng, J.J. The Production of Duckweed as a Source of Biofuels. Biofuels 2012, 3, 589–601. [Google Scholar] [CrossRef]
- Caicedo, J.R.; van der Steen, N.P.; Arce, O.; Gijzen, H.J. Effect of Total Ammonia Nitrogen Concentration and pH on Growth Rates of Duckweed (Spirodela polyrrhiza). Water Res. 2000, 34, 3829–3835. [Google Scholar] [CrossRef]
- Lasfar, S.; Monette, F.; Millette, L.; Azzouz, A. Intrinsic Growth Rate: A New Approach to Evaluate the Effects of Temperature, Photoperiod and Phosphorus–Nitrogen Concentrations on Duckweed Growth under Controlled Eutrophication. Water Res. 2007, 41, 2333–2340. [Google Scholar] [CrossRef]
- Yin, Y.; Yu, C.; Yu, L.; Zhao, J.; Sun, C.; Ma, Y.; Zhou, G. The Influence of Light Intensity and Photoperiod on Duckweed Biomass and Starch Accumulation for Bioethanol Production. Bioresour. Technol. 2015, 187, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Borisjuk, N.; Chu, P.; Gutierrez, R.; Zhang, H.; Acosta, K.; Friesen, N.; Sree, K.S.; Garcia, C.; Appenroth, K.J.; Lam, E. Assessment, Validation and Deployment Strategy of a Two-Barcode Protocol for Facile Genotyping of Duckweed Species. Plant Biol. 2015, 17, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Yamaga, F.; Washio, K.; Morikawa, M. Sustainable Biodegradation of Phenol by Acinetobacter calcoaceticus P23 Isolated from the Rhizosphere of Duckweed Lemna aoukikusa. Environ. Sci. Technol. 2010, 44, 6470–6474. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, Y.; Jog, R.; Morikawa, M. Effects of co-inoculation of two different plant growth-promoting bacteria on duckweed. Plant Growth Regul. 2018, 86, 287–296. [Google Scholar] [CrossRef]
- Suzuki, W.; Sugawara, M.; Miwa, K.; Morikawa, M. Plant growth-promoting bacterium Acinetobacter calcoaceticus P23 increases the chlorophyll content of the Monocot Lemna minor (Duckweed) and the Dicot Lactuca sativa (Lettuce). J. Biosci. Bioeng. 2014, 118, 41–44. [Google Scholar] [CrossRef]
- Idris, E.E.; Iglesias, D.J.; Talon, M.; Borriss, R. Tryptophan-Dependent Production of Indole-3-Acetic Acid (IAA) Affects Level of Plant Growth Promotion by Bacillus amyloliquefaciens FZB42. Mol. Plant Microbe Interact. 2007, 20, 619–626. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, Y.; Meng, H.; Xue, Z.; Ma, J. Complete Genome Sequence of Exiguobacterium sp. Strain MH3, Isolated from Rhizosphere of Lemna minor. Genome Announc. 2013, 1, e01059-13. [Google Scholar] [CrossRef]
- Ishizawa, H.; Kuroda, M.; Morikawa, M.; Ike, M. Evaluation of Environmental Bacterial Communities as a Factor Affecting the Growth of Duckweed Lemna minor. Biotechnol. Biofuels 2017, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Toyama, T.; Mori, K.; Tanaka, Y.; Ike, M.; Morikawa, M. Growth Promotion of Giant Duckweed Spirodela polyrhiza (Lemnaceae) by Ensifer sp. SP4 through Enhancement of Nitrogen Metabolism and Photosynthesis. Mol. Plant Microbe Interact. 2022, 35, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, Y.; Yamamoto, K.; Makino, A.; Tanaka, Y.; Meng, X.-Y.; Hashimoto, J.; Shin-ya, K.; Satoh, N.; Fujie, M.; Toyama, T.; et al. Novel Plant-Associated Acidobacteria Promotes Growth of Common Floating Aquatic Plants, Duckweeds. Microorganisms 2021, 9, 1133. [Google Scholar] [CrossRef] [PubMed]
- Crump, B.C.; Koch, E.W. Attached Bacterial Populations Shared by Four Species of Aquatic Angiosperms. Appl. Environ. Microbiol. 2008, 74, 5948–5957. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, H.; Tanaka, Y.; Tamaki, H.; Kamagata, Y.; Mori, K. Culture-Dependent and Independent Analyses of the Microbial Communities Inhabiting the Giant Duckweed (Spirodela polyrrhiza) Rhizoplane and Isolation of a Variety of Rarely Cultivated Organisms within the Phylum Verrucomicrobia. Microbes Environ. 2010, 25, 302–308. [Google Scholar] [CrossRef]
- Tanaka, Y.; Tamaki, H.; Matsuzawa, H.; Nigaya, M.; Mori, K.; Kamagata, Y. Microbial Community Analysis in the Roots of Aquatic Plants and Isolation of Novel Microbes Including an Organism of the Candidate Phylum OP10. Microbes Environ. 2012, 27, 149–157. [Google Scholar] [CrossRef]
- Utami, D.; Kawahata, A.; Sugawara, M.; Jog, R.N.; Miwa, K.; Morikawa, M. Effect of Exogenous General Plant Growth Regulators on the Growth of the Duckweed Lemna minor. Front. Chem. 2018, 6, 251. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.; Ambrosini, A.; Passaglia, L.M.P. Plant Growth-Promoting Bacteria as Inoculants in Agricultural Soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef] [PubMed]
- Van Hoeck, A.; Horemans, N.; Monsieurs, P.; Cao, H.X.; Vandenhove, H.; Blust, R. The First Draft Genome of the Aquatic Model Plant Lemna minor Opens the Route for Future Stress Physiology Research and Biotechnological Applications. Biotechnol. Biofuels 2015, 8, 188. [Google Scholar] [CrossRef]
- Tanaka, Y.; Matsuzawa, H.; Tamaki, H.; Tagawa, M.; Toyama, T.; Kamagata, Y.; Mori, K. Isolation of Novel Bacteria Including Rarely Cultivated Phyla, Acidobacteria and Verrucomicrobia, from the Roots of Emergent Plants by Simple Culturing Method. Microbes Environ. 2017, 32, 288–292. [Google Scholar] [CrossRef]
- Toyama, T.; Yu, N.; Kumada, H.; Sei, K.; Ike, M.; Fujita, M. Accelerated Aromatic Compounds Degradation in Aquatic Environment by Use of Interaction between Spirodela polyrrhiza and Bacteria in Its Rhizosphere. J. Biosci. Bioeng. 2006, 101, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Reasoner, D.J.; Geldreich, E.E. A New Medium for the Enumeration and Subculture of Bacteria from Potable Water. Appl. Environ. Microbiol. 1985, 49, 7. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Yamagishi, A.; Daimon, S.; Kawasaki, K.; Tamaki, H.; Kitagawa, W.; Abe, A.; Tanaka, M.; Sone, T.; Asano, K.; et al. Isolation of Previously Uncultured Slow-Growing Bacteria by Using a Simple Modification in the Preparation of Agar Media. Appl. Environ. Microbiol. 2018, 84, e00807-18. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, H.; Sekiguchi, Y.; Hanada, S.; Nakamura, K.; Nomura, N.; Matsumura, M.; Kamagata, Y. Comparative Analysis of Bacterial Diversity in Freshwater Sediment of a Shallow Eutrophic Lake by Molecular and Improved Cultivation-Based Techniques. Appl. Environ. Microbiol. 2005, 71, 2162–2169. [Google Scholar] [CrossRef]
- Toyama, T.; Hanaoka, T.; Tanaka, Y.; Morikawa, M.; Mori, K. Comprehensive Evaluation of Nitrogen Removal Rate and Biomass, Ethanol, and Methane Production Yields by Combination of Four Major Duckweeds and Three Types of Wastewater Effluent. Bioresour. Technol. 2018, 250, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of Accurate Extinction Coefficients and Simultaneous Equations for Assaying Chlorophylls a and b Extracted with Four Different Solvents: Verification of the Concentration of Chlorophyll Standards by Atomic Absorption Spectroscopy. Biochim. Biophys. Acta (BBA) Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S rRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Ehmann, A. The van Urk-Salkowski Reagent—a Sensitive and Specific Chromogenic Reagent for Silica Gel Thin-Layer Chromatographic Detection and Identification of Indole Derivatives. J. Chromatogr. A 1977, 132, 267–276. [Google Scholar] [CrossRef]
- Pikovskaya, R. Mobilization of Phosphorus in Soil in Connection with Vital Activity of Some Microbial Species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Alexander, D.B.; Zuberer, D.A. Use of Chrome Azurol S Reagents to Evaluate Siderophore Production by Rhizosphere Bacteria. Biol. Fert. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Ando, S.; Goto, M.; Meunchang, S.; Thongra-ar, P.; Fujiwara, T.; Hayashi, H.; Yoneyama, T. Detection of nifH Sequences in Sugarcane (Saccharum officinarum L.) and Pineapple (Ananas comosus [L.] Merr.). Soil Sci. Plant Nutr. 2005, 51, 303–308. [Google Scholar] [CrossRef]
- Gaby, J.C.; Rishishwar, L.; Valderrama-Aguirre, L.C.; Green, S.J.; Valderrama-Aguirre, A.; Jordan, I.K.; Kostka, J.E. Diazotroph Community Characterization via a High-Throughput nifH Amplicon Sequencing and Analysis Pipeline. Appl. Environ. Microbiol. 2017, 84, e01512-17. [Google Scholar] [CrossRef] [PubMed]
- Barraquio, W.L.; Padre, B.C.; Watanabe, I.; Knowles, R. Nitrogen Fixation by Pseudomonas saccharophila Doudoroff ATCC 15946. Microbiology 1986, 132, 237–241. [Google Scholar] [CrossRef]
- Ahemad, M.; Khan, M.S. Pseudomonasaeruginosa Strain PS1 Enhances Growth Parameters of Greengram [Vignaradiata (L.) Wilczek] in Insecticide-Stressed Soils. J. Pest. Sci. 2011, 84, 123–131. [Google Scholar] [CrossRef]
- Shahid, M.; Saghir Khan, M. Assessment of Glyphosate and Quizalofop Mediated Toxicity to Greengram [Vigna Radiata (L.) Wilczek], Stress Abatement and Growth Promotion by Herbicide Tolerant Bradyrhizobium and Pseudomonas Species. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 3001–3016. [Google Scholar] [CrossRef]
- Rojas-Tapias, D.; Moreno-Galván, A.; Pardo-Díaz, S.; Obando, M.; Rivera, D.; Bonilla, R. Effect of Inoculation with Plant Growth-Promoting Bacteria (PGPB) on Amelioration of Saline Stress in Maize (Zea mays). Appl. Soil Ecol. 2012, 61, 264–272. [Google Scholar] [CrossRef]
- Li, H.Q.; Jiang, X.W. Inoculation with Plant Growth-Promoting Bacteria (PGPB) Improves Salt Tolerance of Maize Seedling. Russ. J. Plant Physiol. 2017, 64, 235–241. [Google Scholar] [CrossRef]
- Grichko, V.P.; Glick, B.R. Amelioration of Flooding Stress by ACC Deaminase-Containing Plant Growth-Promoting Bacteria. Plant Physiol. Biochem. 2001, 39, 11–17. [Google Scholar] [CrossRef]
- Kumar, K.V.; Patra, D.D. Influence of Nickel and Cadmium Resistant PGPB on Metal Accumulation and Growth Responses of Lycopersicon esculentum Plants Grown in Fly Ash Amended Soil. Water Air Soil Pollut. 2013, 224, 1357. [Google Scholar] [CrossRef]
- Figueredo, M.S.; Tonelli, M.L.; Ibáñez, F.; Morla, F.; Cerioni, G.; del Carmen Tordable, M.; Fabra, A. Induced Systemic Resistance and Symbiotic Performance of Peanut Plants Challenged with Fungal Pathogens and Co-Inoculated with the Biocontrol Agent Bacillus sp. CHEP5 and Bradyrhizobium sp. SEMIA6144. Microbiol. Res. 2017, 197, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Mallick, I.; Bhattacharyya, C.; Mukherji, S.; Dey, D.; Sarkar, S.C.; Mukhopadhyay, U.K.; Ghosh, A. Effective Rhizoinoculation and Biofilm Formation by Arsenic Immobilizing Halophilic Plant Growth Promoting Bacteria (PGPB) Isolated from Mangrove Rhizosphere: A Step towards Arsenic Rhizoremediation. Sci. Total Environ. 2018, 610–611, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.-H.; Yokota, A. Reclassification of Alcaligenes latus Strains IAM 12599T and IAM 12664 and Pseudomonas saccharophila as Azohydromonas Lata gen. nov., comb. nov., Azohydromonas australica sp. nov. and Pelomonas saccharophila gen. nov., comb. nov., Respectively. Int. J. Syst. Evol. Microbiol. 2005, 55, 2419–2425. [Google Scholar] [CrossRef] [PubMed]
- Quisehuatl-Tepexicuapan, E. Free-Living Culturable Bacteria and Protozoa from the Rhizoplanes of Three Floating Aquatic Plant Species. Plant Biosyst. 2016, 150, 855–865. [Google Scholar] [CrossRef]
- Rodrigues Coelho, M.R.; de Vos, M.; Carneiro, N.P.; Marriel, I.E.; Paiva, E.; Seldin, L. Diversity of nifH Gene Pools in the Rhizosphere of Two Cultivars of Sorghum ( Sorghum Bicolor ) Treated with Contrasting Levels of Nitrogen Fertilizer. FEMS Microbiol. Lett. 2008, 279, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Terakado-Tonooka, J.; Ohwaki, Y.; Yamakawa, H.; Tanaka, F.; Yoneyama, T.; Fujihara, S. Expressed nifH Genes of Endophytic Bacteria Detected in Field-Grown Sweet Potatoes (Ipomoea batatas L.). Microbes Environ. 2008, 23, 89–93. [Google Scholar] [CrossRef]
- Ferrando, L.; Fernández Scavino, A. Strong Shift in the Diazotrophic Endophytic Bacterial Community Inhabiting Rice (Oryza sativa) Plants after Flooding. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef]
- Gomila, M.; Bowien, B.; Falsen, E.; Moore, E.R.B.; Lalucat, J. Description of Pelomonas aquatica sp. nov. and Pelomonas puraquae sp. nov., Isolated from Industrial and Haemodialysis Water. Int. J. Syst. Evol. Microbiol. 2007, 57, 2629–2635. [Google Scholar] [CrossRef] [PubMed]
- Rivas, R. Bradyrhizobium betae sp. nov., Isolated from Roots of Beta Vulgaris Affected by Tumour-like Deformations. Int. J. Syst. Evol. Microbiol. 2004, 54, 1271–1275. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Kawasaki, H.; Muramatsu, Y.; Nakagawa, Y.; Seki, T. Bradyrhizobium iriomotense sp. nov., Isolated from a Tumor-Like Root of the Legume Entada koshunensis from Iriomote Island in Japan. Biosci. Biotechnol. Biochem. 2008, 72, 1416–1429. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kittiwongwattana, C.; Thawai, C. Rhizobium paknamense sp. nov., Isolated from Lesser Duckweeds (Lemna Aequinoctialis). Int. J. Syst. Evol. Microbiol. 2013, 63, 3823–3828. [Google Scholar] [CrossRef]
- Kittiwongwattana, C.; Thawai, C. Rhizobium lemnae sp. nov., a Bacterial Endophyte of Lemna aequinoctialis. Int. J. Syst. Evol. Microbiol. 2014, 64, 2455–2460. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Fang, Y.; Jin, Y.; Huang, J.; Bao, S.; Fu, T.; He, Z.; Wang, F.; Zhao, H. Potential of Duckweed in the Conversion of Wastewater Nutrients to Valuable Biomass: A Pilot-Scale Comparison with Water Hyacinth. Bioresour. Technol. 2014, 163, 82–91. [Google Scholar] [CrossRef] [PubMed]
- De Lajudie, P.; Laurent-Fulele, E.; Willems, A.; Torek, U.; Coopman, R.; Collins, M.D.; Kersters, K.; Dreyfus, B.; Gillis, M. Allorhizobium undicola gen. nov., sp. nov., Nitrogen-Fixing Bacteria That Efficiently Nodulate Neptunia natans in Senegal. Int. J. Syst. Bacteriol. 1998, 48, 1277–1290. [Google Scholar] [CrossRef]
- Grandlic, C.J.; Palmer, M.W.; Maier, R.M. Optimization of Plant Growth-Promoting Bacteria-Assisted Phytostabilization of Mine Tailings. Soil Biol. Biochem. 2009, 41, 1734–1740. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, P.; Chauhan, P.S.; Saravanan, V.S.; Anandham, R.; Sa, T. Isolation and Characterization of Plant Growth Promoting Endophytic Bacterial Isolates from Root Nodule of Lespedeza sp. Biol. Fertil. Soils 2010, 46, 807–816. [Google Scholar] [CrossRef]
- Guglielmetti, S.; Basilico, R.; Taverniti, V.; Arioli, S.; Piagnani, C.; Bernacchi, A. Luteibacter rhizovicinus MIMR1 Promotes Root Development in Barley (Hordeum vulgare L.) under Laboratory Conditions. World J. Microbiol. Biotechnol. 2013, 29, 2025–2032. [Google Scholar] [CrossRef] [PubMed]
- Subhashini, D.V. Growth Promotion and Increased Potassium Uptake of Tobacco by Potassium-Mobilizing Bacterium Frateuria aurantia Grown at Different Potassium Levels in Vertisols. Commun. Soil Sci. Plant Anal. 2015, 46, 210–220. [Google Scholar] [CrossRef]
- Yang, C.; Hamel, C.; Gan, Y.; Vujanovic, V. Pyrosequencing Reveals How Pulses Influence Rhizobacterial Communities with Feedback on Wheat Growth in the Semiarid Prairie. Plant Soil 2013, 367, 493–505. [Google Scholar] [CrossRef]
- Kamilova, F.; Kravchenko, L.V.; Shaposhnikov, A.I.; Azarova, T.; Makarova, N.; Lugtenberg, B. Organic Acids, Sugars, and l-Tryptophane in Exudates of Vegetables Growing on Stonewool and Their Effects on Activities of Rhizosphere Bacteria. Mol. Plant Microbe Interact. 2006, 19, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.; Xu, J.; Acosta, K.; Poulev, A.; Lebeis, S.; Lam, E. Bacterial Production of Indole Related Compounds Reveals Their Role in Association Between Duckweeds and Endophytes. Front. Chem. 2018, 6, 265. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed]
- Schippers, B.; Bakker, A.W.; Bakker, P.a.H.M. Interactions of Deleterious and Beneficial Rhizosphere Microorganisms and the Effect of Cropping Practices. Annu. Rev. Phytopathol. 1987, 25, 339–358. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of Phosphate Solubilizing Microorganisms in Sustainable Agriculture—A Review. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 551–570. ISBN 978-90-481-2666-8. [Google Scholar]
- Wright, G.S.A.; Saeki, A.; Hikima, T.; Nishizono, Y.; Hisano, T.; Kamaya, M.; Nukina, K.; Nishitani, H.; Nakamura, H.; Yamamoto, M.; et al. Architecture of the Complete Oxygen-Sensing FixL-FixJ Two-Component Signal Transduction System. Sci. Signal. 2018, 11, eaaq0825. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Zhang, X.X.; Sui, X.H.; Li, Y.H.; Young, J.P.W.; Wang, R.; Chen, W.X. Bradyrhizobium guangdongense sp. nov. and Bradyrhizobium guangxiense sp. nov., Isolated from Effective Nodules of Peanut. Int. J. Syst. Evol. Microbiol. 2015, 65, 4655–4661. [Google Scholar] [CrossRef]
- Helene, L.C.F.; Delamuta, J.R.M.; Ribeiro, R.A.; Hungria, M. Bradyrhizobium mercantei sp. nov., a Nitrogen-Fixing Symbiont Isolated from Nodules of Deguelia costata (Syn. Lonchocarpus costatus). Int. J. Syst. Evol. Microbiol. 2017, 67, 1827–1834. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Yoneda, Y.; Makino, A.; Tanaka, Y.; Meng, X.-Y.; Hashimoto, J.; Shin-ya, K.; Satoh, N.; Fujie, M.; Toyama, T.; et al. Draft Genome Sequence of Bryobacteraceae Strain F-183. Microbiol. Resour. Announc. 2022, 11, e00453-21. [Google Scholar] [CrossRef]
- Yamamoto, K.; Yoneda, Y.; Makino, A.; Tanaka, Y.; Meng, X.-Y.; Hashimoto, J.; Shin-ya, K.; Satoh, N.; Fujie, M.; Toyama, T.; et al. Complete Genome Sequence of Luteitalea sp. Strain TBR-22. Microbiol. Resour. Announc. 2022, 11, e00455-21. [Google Scholar] [CrossRef] [PubMed]
- Ishizawa, H.; Tada, M.; Kuroda, M.; Inoue, D.; Ike, M. Performance of Plant Growth-Promoting Bacterium of Duckweed under Different Kinds of Abiotic Stress Factors. Biocatal. Agric. Biotechnol. 2019, 19, 101146. [Google Scholar] [CrossRef]
- Khairina, Y.; Jog, R.; Boonmak, C.; Toyama, T.; Oyama, T.; Morikawa, M. Indigenous Bacteria, an Excellent Reservoir of Functional Plant Growth Promoters for Enhancing Duckweed Biomass Yield on Site. Chemosphere 2021, 268, 129247. [Google Scholar] [CrossRef]
- Mpanga, I.K.; Dapaah, H.K.; Geistlinger, J.; Ludewig, U.; Neumann, G. Soil Type-Dependent Interactions of P-Solubilizing Microorganisms with Organic and Inorganic Fertilizers Mediate Plant Growth Promotion in Tomato. Agronomy 2018, 8, 213. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, R.; Shang, Q.; Hao, H.; Li, Y.; Zhang, Y.; Guo, Z.; Wang, Y.; Xie, Z. The New Flagella-Associated Collagen-like Proteins ClpB and ClpC of Bacillus amyloliquefaciens FZB42 Are Involved in Bacterial Motility. Microbiol. Res. 2016, 184, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol Mechanism by Root-Associated Bacillus amyloliquefaciens FZB42—A Review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bahadur, I.; Maurya, B.R.; Raghuwanshi, R.; Meena, V.S.; Singh, D.K.; Dixit, J. Does a Plant Growth Promoting Rhizobacteria Enhance Agricultural Sustainability? J. Pure Appl. Microbiol. 2015, 9, 715–724. [Google Scholar]
- Sharma, P.; Kumawat, K.C.; Kaur, S. Plant Growth Promoting Rhizobacteria in Nutrient Enrichment: Current Perspectives. In Biofortification of Food Crops; Singh, U., Praharaj, C.S., Singh, S.S., Singh, N.P., Eds.; Springer: New Delhi, India, 2016; pp. 263–289. ISBN 978-81-322-2716-8. [Google Scholar]
- Fan, B.; Chen, X.H.; Budiharjo, A.; Bleiss, W.; Vater, J.; Borriss, R. Efficient Colonization of Plant Roots by the Plant Growth Promoting Bacterium Bacillus amyloliquefaciens FZB42, Engineered to Express Green Fluorescent Protein. J. Biotechnol. 2011, 151, 303–311. [Google Scholar] [CrossRef]
- Seneviratne, G.; Weerasekara, M.L.M.A.W.; Seneviratne, K.A.C.N.; Zavahir, J.S.; Kecskés, M.L.; Kennedy, I.R. Importance of Biofilm Formation in Plant Growth Promoting Rhizobacterial Action. In Plant Growth and Health Promoting Bacteria; Maheshwari, D.K., Ed.; Microbiology Monographs; Springer-Verlag: Berlin/Heidelberg, Germany, 2011; pp. 81–95. ISBN 978-3-642-13612-2. [Google Scholar]
- Qurashi, A.W.; Sabri, A.N. Bacterial Exopolysaccharide and Biofilm Formation Stimulate Chickpea Growth and Soil Aggregation under Salt Stress. Braz. J. Microbiol. 2012, 43, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Tamaki, H.; Tanaka, K.; Tozawa, E.; Matsuzawa, H.; Toyama, T.; Kamagata, Y.; Mori, K. “Duckweed-Microbe Co-Cultivation Method” for Isolating a Wide Variety of Microbes Including Taxonomically Novel Microbes. Microbes Environ. 2018, 33, 402–406. [Google Scholar] [CrossRef] [PubMed]





| Strain | Taxon | Closest Type Strain (Accession Number) | Isolation Source | Identity (%) |
|---|---|---|---|---|
| MRB1 | Betaproteobacteria | Pelomonas saccharophila DSM654T (SMBU01000080) | mud | 99.22 |
| Pelomonas aquatica CCUG52575T (AM501435) | industrial water | 99.21 | ||
| Pelomonas puraquae CCUG52769T (NISI01000035) | hemodialysis water | 99.15 | ||
| MRB2 | Gammaproteobacteria | Aquimonas voraii DSM16957T (jgi.1058856) | warm spring water | 93.82 |
| Fulvimonas soli LMG19981T (AJ311653) | soil | 93.58 | ||
| Dokdonella soli KIS28-6T (EU685334) | mud | 93.51 | ||
| MRB3 | Betaproteobacteria | Pelomonas saccharophila DSM654T (SMBU01000080) | mud | 99.27 |
| Pelomonas aquatica CCUG52575T (AM501435) | industrial water | 99.25 | ||
| Pelomonas puraquae CCUG52769T (NISI01000035) | hemodialysis water | 99.12 | ||
| MRB4 | Alphaproteobacteria | Bradyrhizobium guangdongense CCBAU51649T (KC508867) | nodules of Dalbergia | 99.58 |
| Bradyrhizobium centrosematis A9T (KC247115) | nodules of Centrosema | 99.53 | ||
| Bradyrhizobium ganzhouense RITF806T (JQ796661) | nodules of Acacia | 99.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makino, A.; Nakai, R.; Yoneda, Y.; Toyama, T.; Tanaka, Y.; Meng, X.-Y.; Mori, K.; Ike, M.; Morikawa, M.; Kamagata, Y.; et al. Isolation of Aquatic Plant Growth-Promoting Bacteria for the Floating Plant Duckweed (Lemna minor). Microorganisms 2022, 10, 1564. https://doi.org/10.3390/microorganisms10081564
Makino A, Nakai R, Yoneda Y, Toyama T, Tanaka Y, Meng X-Y, Mori K, Ike M, Morikawa M, Kamagata Y, et al. Isolation of Aquatic Plant Growth-Promoting Bacteria for the Floating Plant Duckweed (Lemna minor). Microorganisms. 2022; 10(8):1564. https://doi.org/10.3390/microorganisms10081564
Chicago/Turabian StyleMakino, Ayaka, Ryosuke Nakai, Yasuko Yoneda, Tadashi Toyama, Yasuhiro Tanaka, Xian-Ying Meng, Kazuhiro Mori, Michihiko Ike, Masaaki Morikawa, Yoichi Kamagata, and et al. 2022. "Isolation of Aquatic Plant Growth-Promoting Bacteria for the Floating Plant Duckweed (Lemna minor)" Microorganisms 10, no. 8: 1564. https://doi.org/10.3390/microorganisms10081564
APA StyleMakino, A., Nakai, R., Yoneda, Y., Toyama, T., Tanaka, Y., Meng, X.-Y., Mori, K., Ike, M., Morikawa, M., Kamagata, Y., & Tamaki, H. (2022). Isolation of Aquatic Plant Growth-Promoting Bacteria for the Floating Plant Duckweed (Lemna minor). Microorganisms, 10(8), 1564. https://doi.org/10.3390/microorganisms10081564