
Phytoplankton Sources and Sinks of Dimethylsulphoniopropionate (DMSP) in Temperate Coastal Waters of Australia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Water Collection and Pre-Treatment
2.2. Incubation Set-Up for Whole Water and Size Fractionated Experiments
2.3. Quantification and Identification of Microbial Community
2.4. Sulfur Chemistry
2.5. Data Analysis
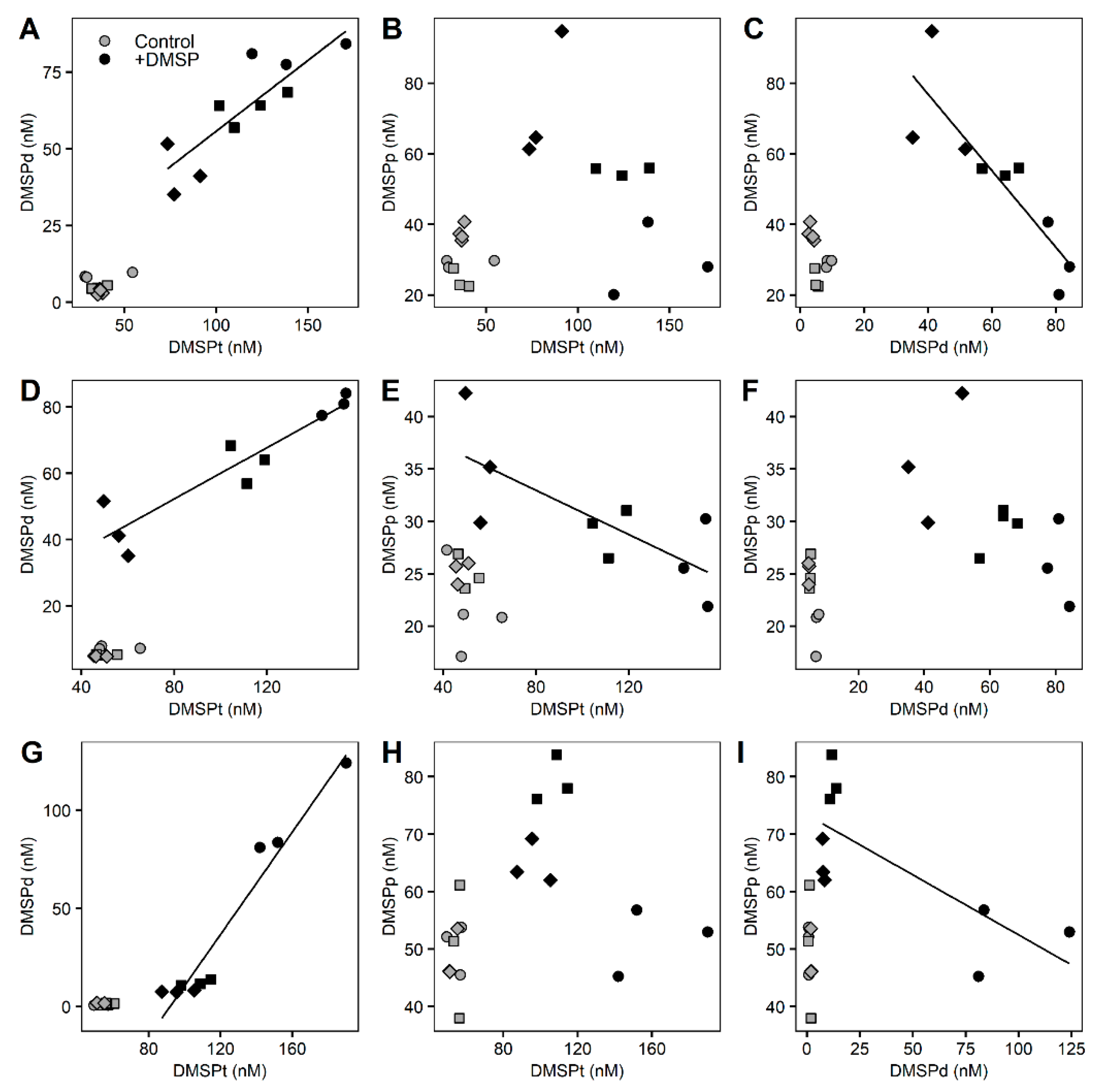
3. Results
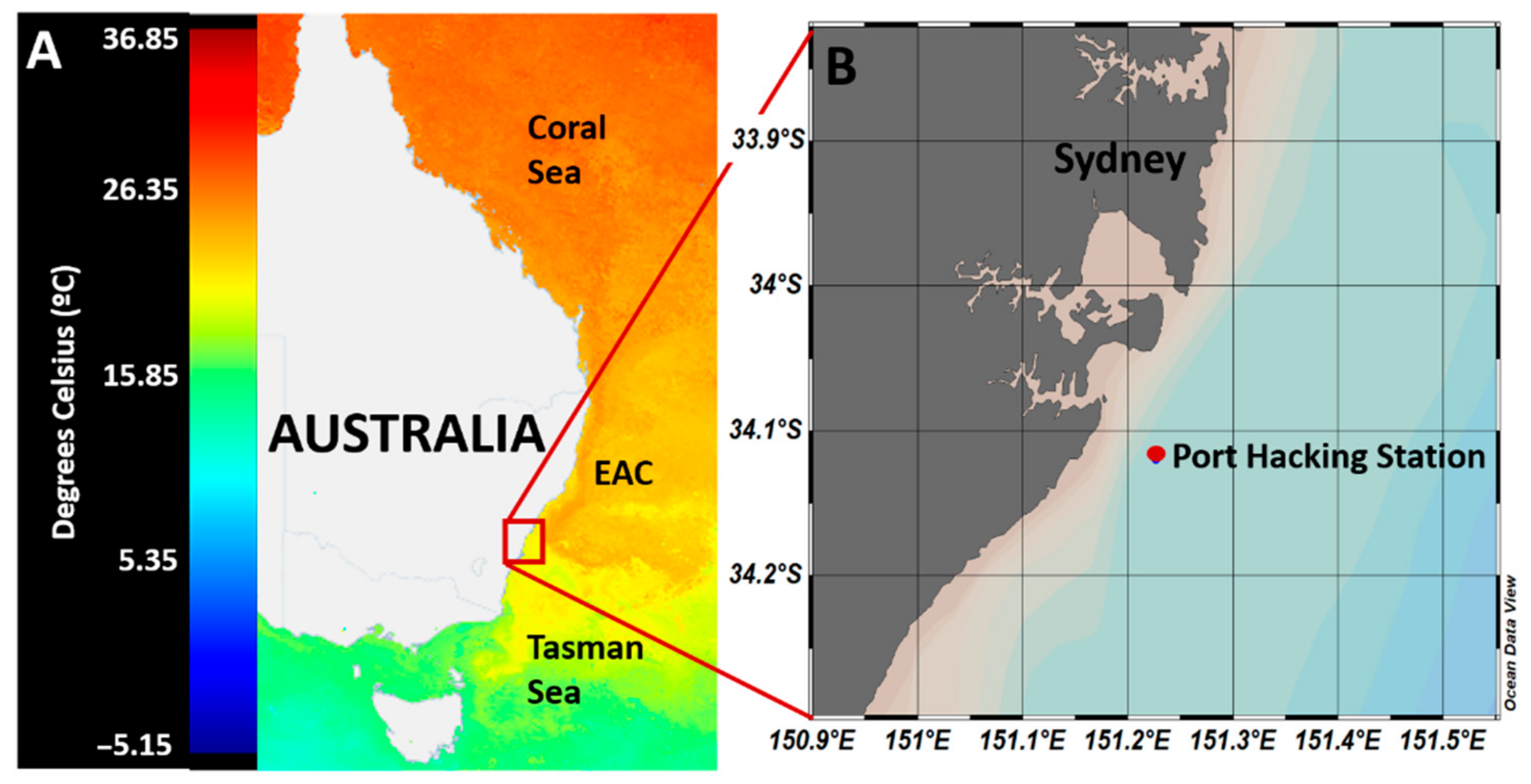
3.1. Characteristics of Initial Water Masses
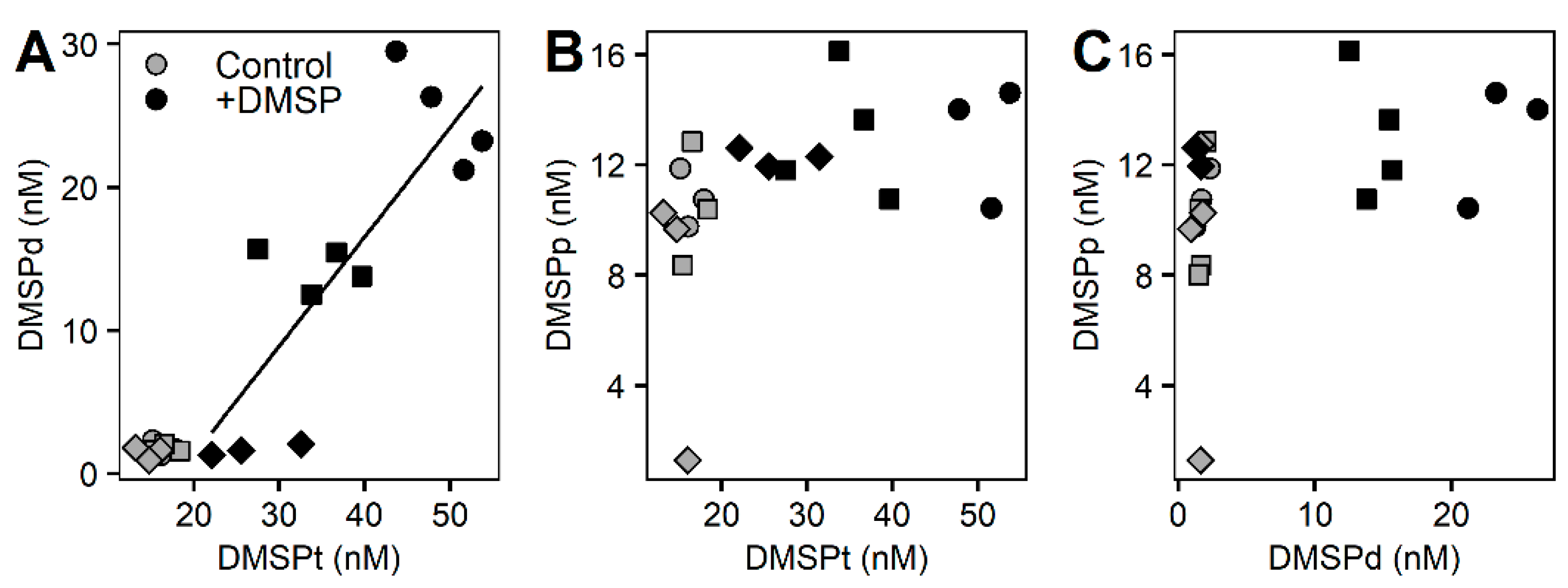
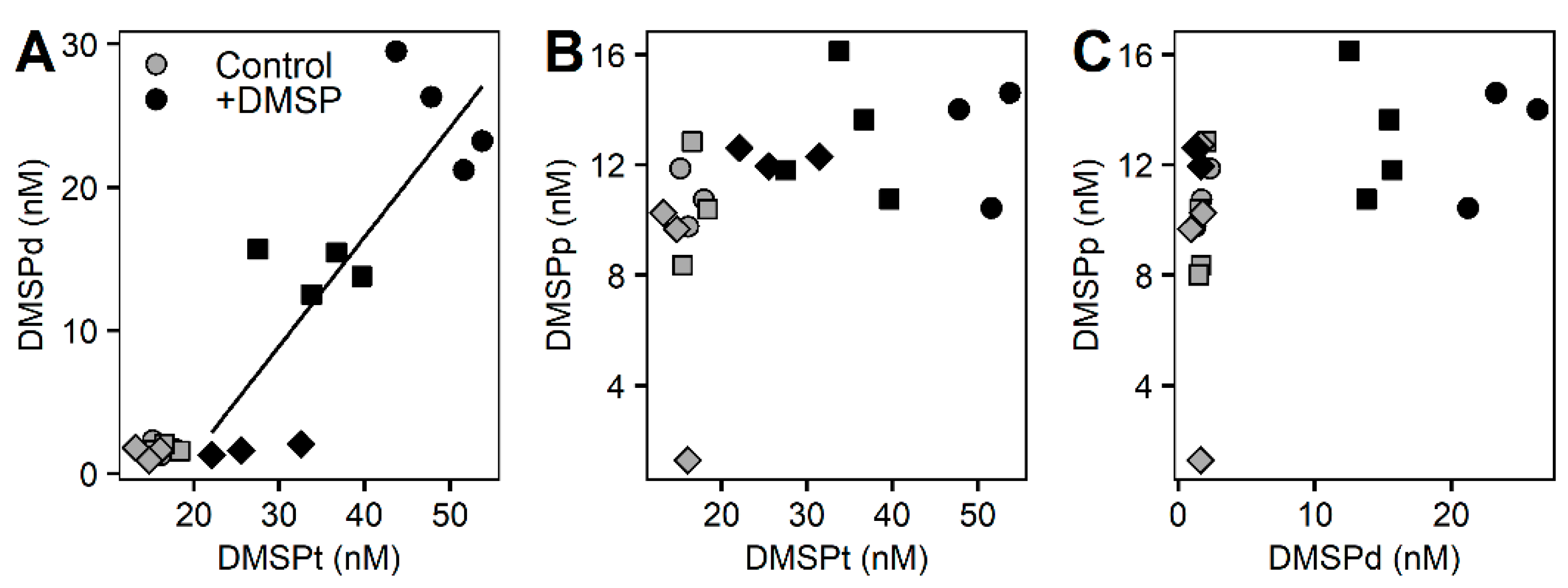
3.2. Whole Community Incubation Experiment
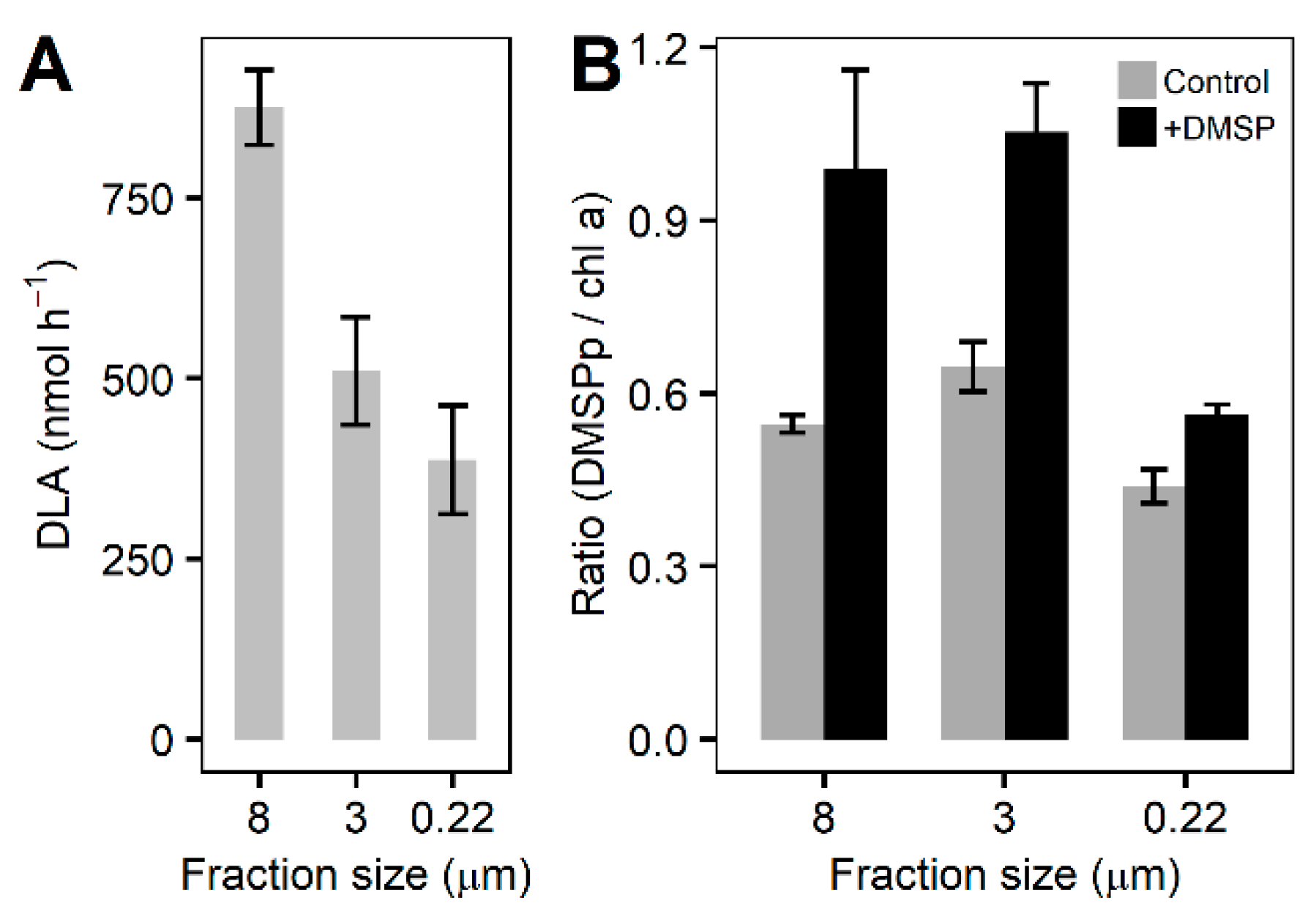
3.3. Fractioned Community Incubation Experiment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DMSP Total | DMSP Dissolved | DMSP Particulate | |||||
|---|---|---|---|---|---|---|---|
| Fraction (µm) | Treatment | Rel. Loss/Gain (%) | Loss/Gain Rate (nM h−1) | Rel. Loss/Gain (%) | Loss/Gain Rate (nM h−1) | Rel. Loss/Gain (%) | Loss/Gain Rate (nM h−1) |
| >8 | Control | - | - | −62. 9 ± 6.8 | −0.9 ± 0.02 | +31.1 ± 7.4 | +1.5 ± 7.3 |
| >8 | +DMSP | −41.8 ±16.2 | −10.4 ± 5.4 | −46.9 ± 12.6 | −6.4 ± 1.9 | +143 ± 166 | +0.3 ± 4.6 |
| >3 | Control | - | - | −40.2 ± 3.8 | −0.4 ± 0.07 | - | - |
| >3 | +DMSP | −63.3 ± 2.3 | −15.8 ± 0.3 | −46.9 ±12.6 | −6.4 ± 1.9 | +41.5 ± 30.3 * | +1.6 ± 1.5 |
| >0.22 | Control | - | - | - | - | - | - |
| >0.22 | +DMSP | −39.1 ± 12.9 | −10.9 ± 5.4 | −91.6 ± 1.9 | −14.7 ± 4.0 | +27.3 ± 22.8 | +2.2 ± 1.6 |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abirami, B.; Radhakrishnan, M.; Kumaran, S.; Wilson, A. Impacts of Global Warming on Marine Microbial Communities. Sci. Total Environ. 2021, 791, 147905. [Google Scholar] [CrossRef] [PubMed]
- Barton, A.D.; Irwin, A.J.; Finkel, Z.V.; Stock, C.A. Anthropogenic Climate Change Drives Shift and Shuffle in North Atlantic Phytoplankton Communities. Mar. Ecol. Prog. Ser. 2016, 113, 2964–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedford, J.; Ostle, C.; Johns, D.G.; Atkinson, A.; Best, M.; Bresnan, E.; Machairopoulou, M.; Graves, C.A.; Devlin, M.; Milligan, A.; et al. Lifeform Indicators Reveal Large-Scale Shifts in Plankton across the North-West European Shelf. Glob. Chang. Biol. 2020, 26, 3482–3497. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.; Costa, R.R.; Dotto, T.S.; Kerr, R.; Tavano, V.M.; Brito, A.C.; Brotas, V.; Secchi, E.R.; Mendes, C.R.B. Changes in Phytoplankton Communities Along the Northern Antarctic Peninsula: Causes, Impacts and Research Priorities. Front. Mar. Sci. 2020, 7, 576254. [Google Scholar] [CrossRef]
- Hjerne, O.; Hajdu, S.; Larsson, U.; Downing, A.S.; Winder, M. Climate Driven Changes in Timing, Composition and Magnitude of the Baltic Sea Phytoplankton Spring Bloom. Front. Mar. Sci. 2019, 6, 482. [Google Scholar] [CrossRef] [Green Version]
- Wasmund, N.; Nausch, G.; Gerth, M.; Busch, S.; Burmeister, C.; Hansen, R.; Sadkowiak, B. Extension of the Growing Season of Phytoplankton in the Western Baltic Sea in Response to Climate Change. Mar. Ecol. Prog. Ser. 2019, 622, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Behrenfeld, M.J.; Halsey, K.H.; Milligan, A.J. Evolved Physiological Responses of Phytoplankton to Their Integrated Growth Environment. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 2687–2703. [Google Scholar] [CrossRef] [Green Version]
- Mackey, K.R.M.; Paytan, A.; Grossman, A.R.; Bailey, S. A Photosynthetic Strategy for Coping in a High-Light, Low-Nutrient Environment. Limnol. Oceanogr. 2008, 53, 900–913. [Google Scholar] [CrossRef] [Green Version]
- Karsten, U.; Kück, K.; Vogt, C.; Kirst, G. Dimethylsulfoniopropionate Production in Phototrophic Organisms and Its Physiological Functions as a Cryoprotectant. In Biological and Environmental Chemistry of DMSP and Related Sulfonium Compounds; Kiene, R.P., Visscher, P.T., Keller, M.D., Kirst, G.o., Visscher, P.T., Eds.; Plenum Press: New York, NY, USA, 1996; pp. 143–153. [Google Scholar]
- Vance, T.R.; Davidson, A.T.; Thomson, P.G.; Levasseur, M.; Lizotte, M.; Curran, M.A.J.; Jones, G.B. Rapid DMSP Production by an Antarctic Phytoplankton Community Exposed to Natural Surface Irradiances in Late Spring. Aquat. Microb. Ecol. 2013, 71, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Darroch, L.; Lavoie, M.; Levasseur, M.; Laurion, I.; Sunda, W.; Michaud, S.; Scarratt, M.; Gosselin, M.; Caron, G. Effect of Short-Term Light- and UV-Stress on DMSP, DMS, and DMSP Lyase Activity in Emiliania huxleyi. Aquat. Microb. Ecol. 2015, 74, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Dickson, D.; Kirst, G. The Role of β-Dimethylsulphoniopropionate, Glycine Betaine and Homarine in the Osmoacclimation of Platymonas subcordiformis. Planta 1986, 167, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Stefels, J.; Gieskes, W.W.C.; Dijkhuizen, L. Intriguing Functionality of the Production and Conversion of DMSP in Phaeocystis SP. In Biological and Environmental Chemistry of DMSP and Related Sulfonium Compounds; Kiene, R.P., Visscher, P.T., Keller, M.D., Kirst, G.O., Eds.; Springer: Boston, MA, USA, 1996. [Google Scholar] [CrossRef]
- Sunda, W.; Kieber, D.J.; Kiene, R.P.; Huntsman, S. An Antioxidant Function for DMSP and DMS in Marine Algae. Nature 2002, 418, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.M.D.; Bellows, W.W.K.; Guillard, R.R.R.L. Dimethyl Sulfide Production in Marine Phytoplankton. In Biogenic Sulfur in the Environment; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1989; Volume 393, pp. 167–182. [Google Scholar]
- Belviso, S.; Claustre, H.; Marty, J.-C. Evaluation of the Utility of Chemotaxonomic Pigments as a Surrogate for Particulate DMSP. Limnol. Oceanogr. 2001, 46, 989–995. [Google Scholar] [CrossRef] [Green Version]
- McParland, E.L.; Levine, N.M. The Role of Differential DMSP Production and Community Composition in Predicting Variability of Global Surface DMSP Concentrations. Limnol. Oceanogr. 2019, 64, 757–773. [Google Scholar] [CrossRef] [Green Version]
- Bucciarelli, E.; Ridame, C.; Sunda, W.G.; Dimier-Hugueney, C.; Cheize, M.; Belviso, S. Increased Intracellular Concentrations of DMSP and DMSO in Iron-Limited Oceanic Phytoplankton Thalassiosira oceanica and Trichodesmium erythraeum. Limnol. Oceanogr. 2013, 58, 1667–1679. [Google Scholar] [CrossRef] [Green Version]
- Kiene, R.P.; Linn, L.J.; Bruton, J.A. New and Important Roles for DMSP in Marine Microbial Communities. J. Sea Res. 2000, 43, 209–224. [Google Scholar] [CrossRef]
- Simó, R. From Cells to Globe: Approaching the Dynamics of DMS(P) in the Ocean at Multiple Scales. Can. J. Fish. Aquat. Sci. 2004, 61, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Kiene, R.P.; Linn, L.J.; González, J.; Moran, M.A.; Bruton, J.A. Dimethylsulfoniopropionate and Methanethiol Are Important Precursors of Methionine and Protein-Sulfur in Marine Bacterioplankton. Appl. Environ. Microbiol. 1999, 65, 4549–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiene, R.P.; Linn, L.J. Distribution and Turnover of Dissolved DMSP and Its Relationship with Bacterial Production and Dimethylsulfide in the Gulf of Mexico. Limnol. Oceanogr. 2000, 45, 849–861. [Google Scholar] [CrossRef]
- Yoch, D.C. Dimethylsulfoniopropionate: Its Sources, Role in the Marine Food Web, and Biological Degradation to Dimethylsulfide. Appl. Environ. Microbiol. 2002, 68, 5804–5815. [Google Scholar] [CrossRef] [Green Version]
- Bullock, H.A.; Luo, H.; Whitman, W.B. Evolution of Dimethylsulfoniopropionate Metabolism in Marine Phytoplankton and Bacteria. Front. Microbiol. 2017, 8, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohapatra, B.R.; Rellinger, A.N.; Kieber, D.J.; Kiene, R.P. Kinetics of DMSP Lyases in Whole Cell Extracts of Four Phaeocystis Species: Response to Temperature and DMSP Analogs. J. Sea Res. 2014, 86, 110–115. [Google Scholar] [CrossRef]
- Steinke, M.; Malin, G.; Archer, S.; Burkill, P.; Liss, P. DMS Production in a Coccolithophorid Bloom: Evidence for the Importance of Dinoflagellate DMSP Lyases. Aquat. Microb. Ecol. 2002, 26, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Steinke, M.; Wolfe, G.; Kirst, G. Partial Characterisation of Dimethylsulfoniopropionate (DMSP) Lyase Isozymes in 6 Strains of Emiliania huxleyi. Mar. Ecol. Prog. Ser. 1998, 175, 215–225. [Google Scholar] [CrossRef]
- Wolfe, G.; Steinke, M. Grazing-activated Production of Dimethyl Sulfide (DMS) by Two Clones of Emiliania huxleyi. Limnol. Oceanogr. 1996, 41, 1151–1160. [Google Scholar] [CrossRef] [Green Version]
- Yost, D.M.; Mitchelmore, C.L. Substrate Kinetics of DMSP-Lyases in Various Cultured Symbiodinium Strains. Bull. Mar. Sci. 2012, 88, 267–282. [Google Scholar] [CrossRef]
- Van Boekel, J.; Stefels, W.; Stefels, J.; van Boekel, W. Production of DMS from Dissolved DMSP in Axenic Cultures of the Marine Phytoplankton Species Phaeocystis sp. Mar. Ecol. Prog. Ser. 1993, 97, 11–18. [Google Scholar] [CrossRef]
- Vila-Costa, M.; del Valle, D.A.; González, J.M.; Slezak, D.; Kiene, R.P.; Sánchez, O.; Simó, R. Phylogenetic Identification and Metabolism of Marine Dimethylsulfide-Consuming Bacteria. Environ. Microbiol. 2006, 8, 2189–2200. [Google Scholar] [CrossRef] [PubMed]
- Petrou, K.; Nielsen, D.A. Uptake of Dimethylsulphoniopropionate (DMSP) by the Diatom Thalassiosira weissflogii: A Model to Investigate the Cellular Function of DMSP. Biogeochemistry 2018, 141, 265–271. [Google Scholar] [CrossRef]
- Fernandez, E.; Ostrowski, M.; Siboni, N.; Seymour, J.R.; Petrou, K. Uptake of Dimethylsulfoniopropionate (DMSP) by Natural Microbial Communities of the Great Barrier Reef (GBR), Australia. Microorganisms 2021, 9, 1891. [Google Scholar] [CrossRef]
- Vila-Costa, M.; Simó, R.; Harada, H.; Gasol, J.M.; Slezak, D.; Kiene, R.P. Dimethylsulfoniopropionate Uptake by Marine Phytoplankton. Science 2006, 314, 652–654. [Google Scholar] [CrossRef] [PubMed]
- Ridgway, K.; Dunn, J. Mesoscale Structure of the Mean East Australian Current System and Its Relationship with Topography. Prog. Oceanogr. 2003, 56, 189–222. [Google Scholar] [CrossRef]
- Cetina-Heredia, P.; Roughan, M.; van Sebille, E.; Coleman, M.A. Long-Term Trends in the East Australian Current Separation Latitude and Eddy Driven Transport. J. Geophys. Res. Ocean. 2014, 119, 4351–4366. [Google Scholar] [CrossRef] [Green Version]
- Middleton, J.H.; Cox, D.; Tate, P. The Oceanography of the Sydney Region. Mar. Pollut. Bull. 1996, 33, 124–131. [Google Scholar] [CrossRef]
- IMOS. Available online: imos.org.au (accessed on 9 September 2019).
- Schlitzer, R. Data Analysis and Visualization with Ocean Data View. CMOS Bull. SCMO 2015, 43, 9–13. [Google Scholar]
- Marie, D.; Partensky, F.; Jacquet, S.; Vaulot, D. Enumeration and Cell Cycle Analysis of Natural Populations of Marine Picoplankton by Flow Cytometry Using the Nucleic Acid Stain SYBR Green I. Appl. Environ. Microbiol. 1997, 63, 186–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasol, J.M.; Del Giorgio, P.A. Using Flow Cytometry for Counting Natural Planktonic Bacteria and Understanding the Structure of Planktonic Bacterial Communities. Sci. Mar. 2000, 64, 197–224. [Google Scholar] [CrossRef] [Green Version]
- Seymour, J.R.; Seuront, L.; Mitchell, J.G. Microscale Gradients of Planktonic Microbial Communities above the Sediment Surface in a Mangrove Estuary. Estuar. Coast. Shelf Sci. 2007, 73, 651–666. [Google Scholar] [CrossRef]
- Dacey, J.W.H.; Blough, N.V. Hydroxide Decomposition of Dimethylsulfoniopropionate to Form Dimethylsulfide. Geophys. Res. Lett. 1987, 14, 1246–1249. [Google Scholar] [CrossRef]
- Harada, H.; Rouse, M.-A.; Sunda, W.; Kiene, R.P. Latitudinal and Vertical Distributions of Particle-Associated Dimethylsulfoniopropionate (DMSP) Lyase Activity in the Western North Atlantic Ocean. Can. J. Fish. Aquat. Sci. 2004, 61, 700–711. [Google Scholar] [CrossRef]
- Steinke, M.; Malin, G.; Turner, S.; Liss, P. Determinations of Dimethylsulphoniopropionate (DMSP) Lyase Activity Using Headspace Analysis of Dimethylsulphide (DMS). J. Sea Res. 2000, 43, 233–244. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER V6: User Manual/Tutorial. 2006, p. 192. Available online: https://www.scienceopen.com/document?vid=2cd68314-640b-4288-8316-532e8932d7a1 (accessed on 2 March 2020).
- Anderson, M.J. Permanova: A Fortran Computer Program for Permutational Multivariate Analysis of Variance; Department of Statistics, University of Auckland: Auckland, New Zealand, 2005. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Winder, M.; Sommer, U. Phytoplankton Response to a Changing Climate. Hydrobiologia 2012, 698, 5–16. [Google Scholar] [CrossRef]
- Suthers, I.M.; Young, J.W.; Baird, M.E.; Roughan, M.; Everett, J.D.; Brassington, G.B.; Byrne, M.; Condie, S.A.; Hartog, J.R.; Hassler, C.S.; et al. The Strengthening East Australian Current, Its Eddies and Biological Effects—An Introduction and Overview. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 538–546. [Google Scholar] [CrossRef]
- Ajani, P.A.; Allen, A.P.; Ingleton, T.; Armand, L. A Decadal Decline in Relative Abundance and a Shift in Microphytoplankton Composition at a Long-Term Coastal Station off Southeast Australia. Limnol. Oceanogr. 2014, 59, 519–531. [Google Scholar] [CrossRef]
- Doblin, M.A.; Petrou, K.; Sinutok, S.; Seymour, J.R.; Messer, L.F.; Brown, M.V.; Norman, L.; Everett, J.D.; McInnes, A.S.; Ralph, P.J.; et al. Nutrient Uplift in a Cyclonic Eddy Increases Diversity, Primary Productivity and Iron Demand of Microbial Communities Relative to a Western Boundary Current. PeerJ 2016, 4, e1973. [Google Scholar] [CrossRef] [Green Version]
- Hassler, C.S.; Djajadikarta, J.R.; Doblin, M.A.; Everett, J.D.; Thompson, P.A. Characterisation of Water Masses and Phytoplankton Nutrient Limitation in the East Australian Current Separation Zone during Spring 2008. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 664–677. [Google Scholar] [CrossRef]
- Capone, D.G.; Zehr, J.P.; Paerl, H.W.; Bergman, B.; Carpenter, E.J. Trichodesmium, a Globally Significant Marine Cyanobacterium. Science 1997, 276, 1221–1229. [Google Scholar] [CrossRef] [Green Version]
- Zingone, A.; Phlips, E.J.; Harrison, P.J. Multiscale Variability of Twenty-Two Coastal Phytoplankton Time Series: A Global Scale Comparison. Estuaries Coasts 2010, 33, 224–229. [Google Scholar] [CrossRef]
- Lee, M.D.; Walworth, N.G.; McParland, E.L.; Fu, F.-X.; Mincer, T.J.; Levine, N.M.; Hutchins, D.A.; Webb, E.A. The Trichodesmium Consortium: Conserved Heterotrophic Co-Occurrence and Genomic Signatures of Potential Interactions. ISME J. 2017, 11, 1813–1824. [Google Scholar] [CrossRef] [Green Version]
- McParland, E.L.; Wright, A.; Art, K.; He, M.; Levine, N.M. Evidence for Contrasting Roles of Dimethylsulfoniopropionate Production in Emiliania huxleyi and Thalassiosira oceanica. New Phytol. 2020, 226, 396–409. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; McParland, E.; Bramucci, A.; Ostrowski, M.; Siboni, N.; Ingleton, T.; Brown, M.; Levine, N.; Laverock, B.; Petrou, K.; et al. The Microbiological Drivers of Temporally Dynamic DMSP Cycling Processes in Australian Coastal Shelf Waters. Front. Microbiol. 2022, 13, 894026. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.D. Dimethyl Sulfide Production and Marine Phytoplankton: The Importance of Species Composition and Cell Size. Biol. Oceanogr. 1989, 6, 375–383. [Google Scholar]
- Speeckaert, G.; Borges, A.V.; Champenois, W.; Royer, C.; Gypensa, N. Annual Cycle of Dimethylsulfoniopropionate (DMSP) and Dimethylsulfoxide (DMSO) Related to Phytoplankton Succession in the Southern North Sea. Sci. Total Environ. 2018, 622–623, 362–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefels, J. Physiological Aspects of the Production and Conversion of DMSP in Marine Algae and Higher Plants. J. Sea Res. 2000, 43, 183–197. [Google Scholar] [CrossRef]
- Kiene, R.P.; Linn, L.J. The Fate of Dissolved Dimethylsulfoniopropionate (DMSP) in Seawater: Tracer Studies Using 35S-DMSP. Geochim. Cosmochim. Acta 2000, 64, 2797–2810. [Google Scholar] [CrossRef]
- Theseira, A.M.; Nielsen, D.A.; Petrou, K. Uptake of Dimethylsulphoniopropionate (DMSP) Reduces Free Reactive Oxygen Species (ROS) during Late Exponential Growth in the Diatom Thalassiosira weissflogii Grown under Three Salinities. Mar. Biol. 2020, 167, 127. [Google Scholar] [CrossRef]
- Finkel, Z.V.; Follows, M.J.; Irwin, A.J. Size-Scaling of Macromolecules and Chemical Energy Content in the Eukaryotic Microalgae. J. Plankton Res. 2016, 38, 1151–1162. [Google Scholar] [CrossRef] [Green Version]


| Parameter | |
|---|---|
| Salinity (psu) | 35.4 |
| Chlorophyll a (µg L−1) | 0.306 |
| Silicate (µM) | 0.5 |
| Nitrate (µM) | nd |
| Phosphate (µM) | 0.09 |
| Ammonium (µM) | 0.09 |
| DMS (nM) | 1.51 ± 0.06 |
| DMSPtotal (nM) | 16.4 ± 1.41 |
| DMSPdissolved (nM) | 1.76 ± 0.65 |
| DLAphyto (nM h−1) | 2281 ± 205 |
| DLAbacto (nM h−1) | 3347 ± 168 |
| Microbial Composition | |
|---|---|
| Synechococcus (cells × 105 mL−1) | 1.35 ± 0.01 |
| Prochlorococcus (cells × 104 mL−1) | 4.89 ± 0.58 |
| Heterotrophic Bacteria (cells × 105 mL−1) | 1.37 ± 0.34 |
| Picoeukaryotes (cells × 104 mL−1) | 2.01 ± 0.40 |
| Group | Species | Cells L−1 | Biovolume (µm3 L−1) | Size Fraction | DMSP Producer (Hi/Lo) |
|---|---|---|---|---|---|
| Centric diatoms | Skeletonema spp. | 164 | 67,453 | 3–8, >8 µm | Lo |
| Climacodium spp. | 235 | 79,516,888 | >8 µm | nd | |
| Dactyliosolen spp. | 211 | 10,370,566 | >8 µm | nd | |
| Chaetoceros spp. | 47 | 17,699 | 3–8, >8 µm | Lo | |
| Pennate diatoms | Cylindrotheca closterium | 915 | 183,099 | >8 µm | Lo |
| Cyanobacteria | Trichodesmium spp. | 3286 | 1,881,635 | >8 µm | Lo |
| Dinoflagellates | Gyrodinium spp. (20–40 µm) | 23 | 184,366 | >8 µm | Lo |
| Tripos ^ candelabrum | 23 | 1,260,876 | >8 µm | Hi | |
| Tripos ^ macroceros | 23 | 1,622,417 | >8 µm | Hi | |
| Protoperidinium spp. | 23 | 1,812,388 | >8 µm | Hi | |
| Prymnesiophyte | Phaeocystis sp. | 854 | 96,541 | 3–8 µm | Hi |
| Chlorophyte | Prasinophyte | 4268 | 1,117,367 | 3–8 µm | Lo * |
| Silicoflagellate | Dyctyocha fibula | 23 | 96,024 | >8 µm | nd |
| Other | Unid. Dinoflagellate (<10 µm) | 7682 | 2,316,973 | 3–8, >8 µm | Hi * |
| Flagellates <10 µm | 168,161 | 11,006,069 | 3–8, >8 µm | nd | |
| Cryptophytes <10 µm | 18,779 | 14,749,249 | >8 µm | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez, E.; Seymour, J.R.; Petrou, K. Phytoplankton Sources and Sinks of Dimethylsulphoniopropionate (DMSP) in Temperate Coastal Waters of Australia. Microorganisms 2022, 10, 1539. https://doi.org/10.3390/microorganisms10081539
Fernandez E, Seymour JR, Petrou K. Phytoplankton Sources and Sinks of Dimethylsulphoniopropionate (DMSP) in Temperate Coastal Waters of Australia. Microorganisms. 2022; 10(8):1539. https://doi.org/10.3390/microorganisms10081539
Chicago/Turabian StyleFernandez, Eva, Justin R. Seymour, and Katherina Petrou. 2022. "Phytoplankton Sources and Sinks of Dimethylsulphoniopropionate (DMSP) in Temperate Coastal Waters of Australia" Microorganisms 10, no. 8: 1539. https://doi.org/10.3390/microorganisms10081539
APA StyleFernandez, E., Seymour, J. R., & Petrou, K. (2022). Phytoplankton Sources and Sinks of Dimethylsulphoniopropionate (DMSP) in Temperate Coastal Waters of Australia. Microorganisms, 10(8), 1539. https://doi.org/10.3390/microorganisms10081539