
Sharing Vitamin B12 between Bacteria and Microalgae Does Not Systematically Occur: Case Study of the Haptophyte Tisochrysis lutea

 ,
,  , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Screening
2.2. Bacterial Genome Sequencing
2.3. Phylogenetic Analysis of the Bacteria
2.4. T. lutea–Bacteria Co-Cultures and Analyses
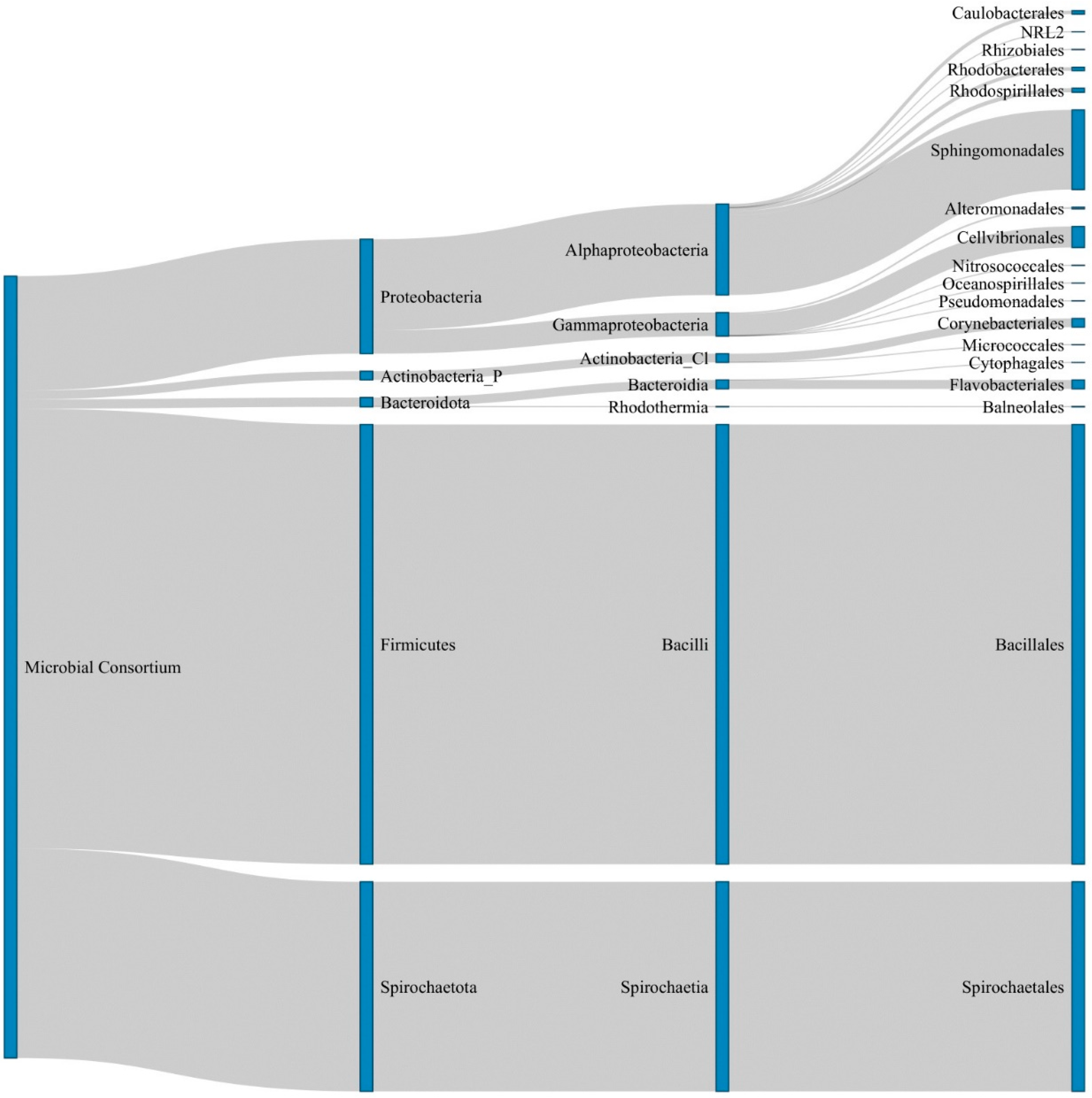
2.5. Identification of the Microbial Consortium
2.6. Statistical Analyses
3. Results
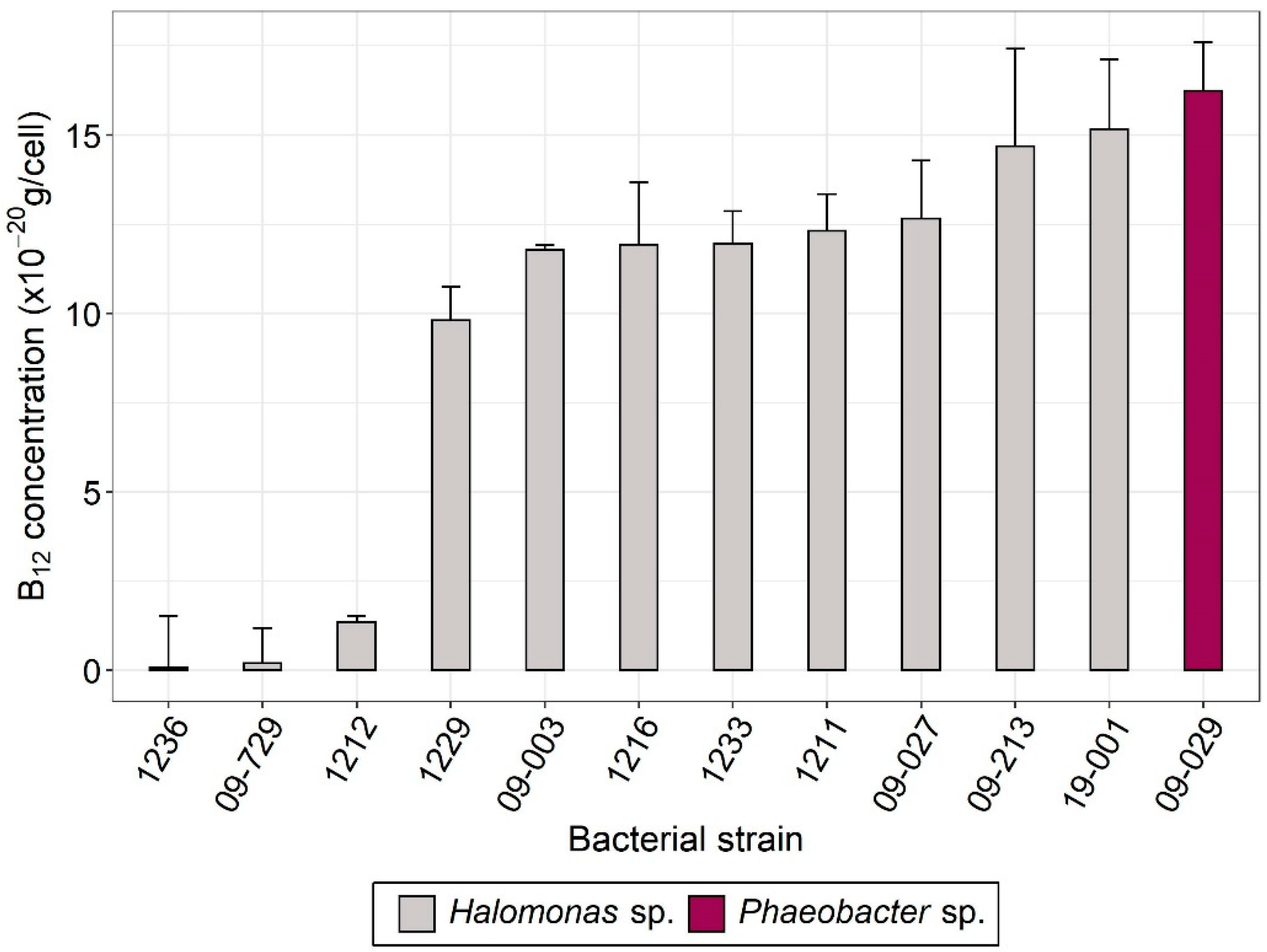
3.1. Selection of Vitamin B12-Producing Bacteria
3.2. Genomic Analyses of the B12-Producing Bacteria
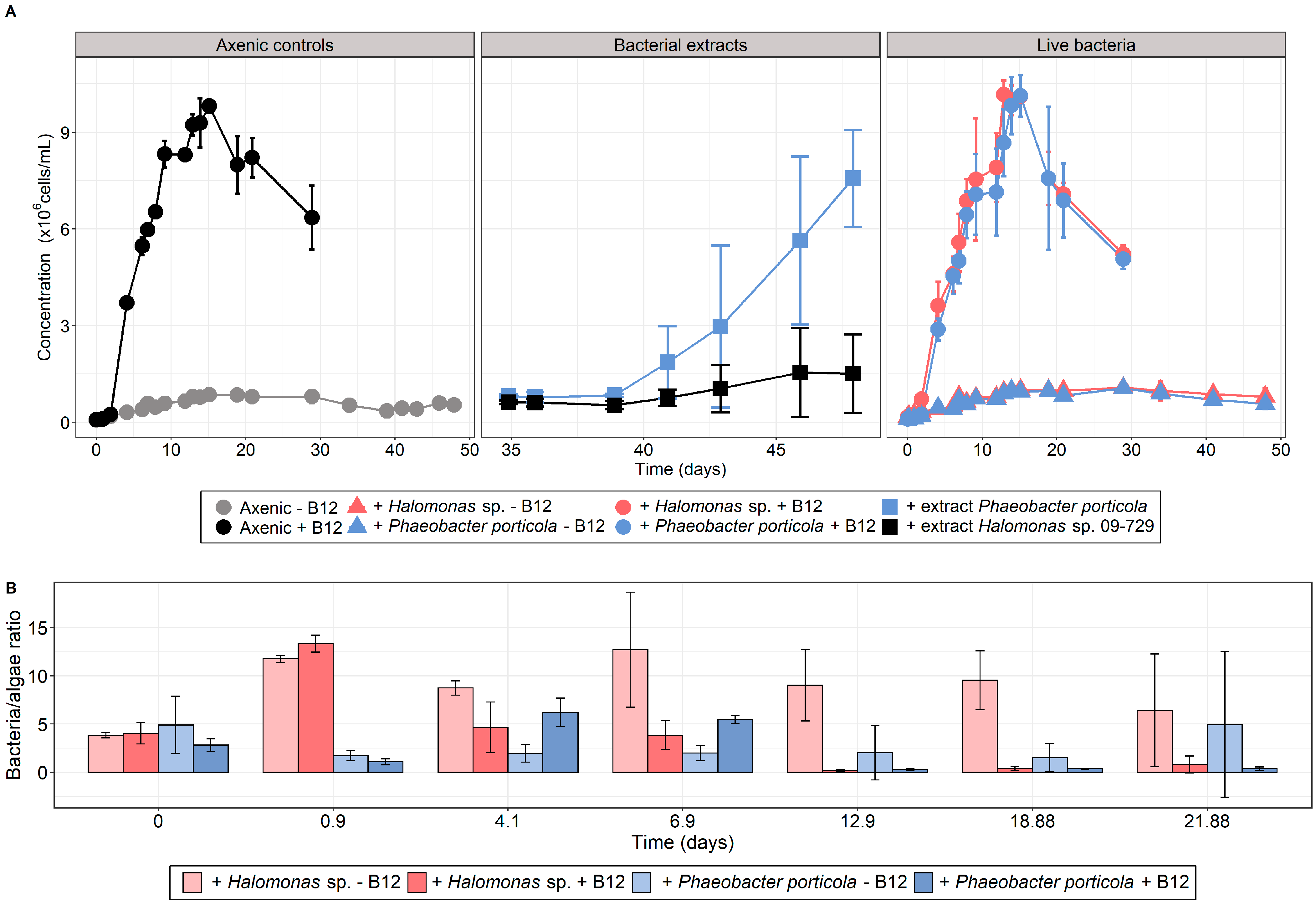
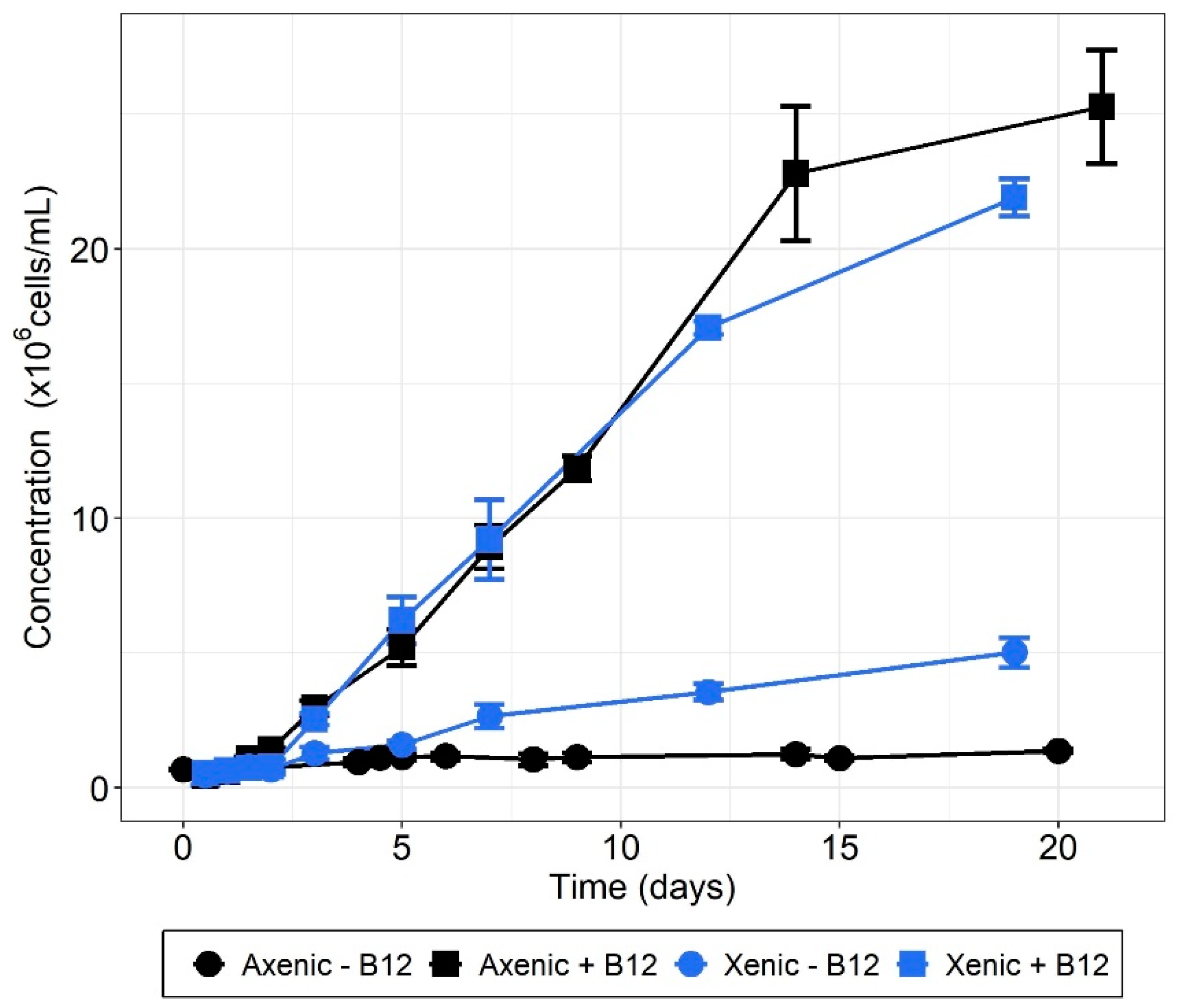
3.3. T. lutea–Bacteria Co-Cultures
3.4. Functional Prediction of De Novo Cobalamin Synthesis in the Microbial Consoritum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Banerjee, R.V.; Matthews, R.G. Cobalamin-Dependent Methionine Synthase. FASEB J. 1990, 5, 1450–1459. [Google Scholar] [CrossRef] [PubMed]
- Martens, J.-H.; Barg, H.; Warren, M.J.; Jahn, D. Microbial Production of Vitamin B12. Appl. Microbiol. Biotechnol. 2002, 58, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Provasoli, C. Vitamins and Growth Regulation. In Algal Physiology and Biochemistry; Botanical monographs; University of California Press: Berkeley, CA, USA, 1974; pp. 741–787. ISBN 978-0-520-02410-6. [Google Scholar]
- Croft, M.T.; Lawrence, A.D.; Raux-Deery, E.; Warren, M.J.; Smith, A.G. Algae Acquire Vitamin B12 through a Symbiotic Relationship with Bacteria. Nature 2005, 483, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Croft, M.T.; Warren, M.J.; Smith, A.G. Algae Need Their Vitamins. Eukaryot. Cell 2006, 5, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Gobler, C.J.; Norman, C.; Panzeca, C.; Taylor, G.T.; Sañudo-Wilhelmy, S.A. Effect of B-Vitamins (B1, B12) and Inorganic Nutrients on Algal Bloom Dynamics in a Coastal Ecosystem. Aquat. Microb. Ecol. 2007, 49, 181–194. [Google Scholar] [CrossRef]
- Sañudo-Wilhelmy, S.A.; Gobler, C.J.; Obkamichael, M.; Taylor, G.T. Regulation of Phytoplankton Dynamics by Vitamin B12. Geophys. Res. Lett. 2006, 33, 1–4. [Google Scholar] [CrossRef]
- Koch, F.; Marcoval, M.A.; Panzeca, C.; Bruland, K.W.; Sañudo-Wilhelmy, S.A.; Gobler, C.J. The Effect of Vitamin B12 on Phytoplankton Growth and Community Structure in the Gulf of Alaska. Limnol. Oceanogr. 2011, 56, 1023–1034. [Google Scholar] [CrossRef]
- Panzeca, C.; Tovar-Sanchez, A.; Agusti, S.; Reche, I.; Duarte, C.M.; Taylor, G.T.; Sañudo-Wilhelmy, S.A. B Vitamins as Regulators of Phytoplankton Dynamics. Eos Trans. Am. Geophys. Union 2006, 49, 594–596. [Google Scholar] [CrossRef]
- Raux, E.; Lanois, A.; Levillayer, F.; Warren, M.J.; Brody, E.; Rambach, A.; Thermes, C. Salmonella Typhimurium Cobalamin (Vitamin B12) Biosynthetic Genes: Functional Studies in S. Typhimurium and Escherichia Coli. J. Bacteriol. 1996, 178, 753–767. [Google Scholar] [CrossRef]
- Raux, E.; Schubert, H.L.; Warren, M.J. Biosynthesis of Cobalamin (Vitamin B12): A Bacterial Conundrum. Cell. Mol. Life Sci. 2000, 57, 1880–1893. [Google Scholar] [CrossRef]
- Doxey, A.C.; Kurtz, D.A.; Lynch, M.D.; Sauder, L.A.; Neufeld, J.D. Aquatic Metagenomes Implicate Thaumarchaeota in Global Cobalamin Production. ISME J. 2015, 9, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Romine, M.F.; Rodionov, D.A.; Maezato, Y.; Osterman, A.L.; Nelson, W.C. Underlying Mechanisms for Syntrophic Metabolism of Essential Enzyme Cofactors in Microbial Communities. ISME J. 2017, 11, 1434–1446. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Moran, M.A. Evolutionary Ecology of the Marine Roseobacter Clade. Microbiol. Mol. Biol. Rev. 2014, 78, 573–587. [Google Scholar] [CrossRef]
- Kazamia, E.; Czesnick, H.; Van Nguyen, T.T.; Croft, M.T.; Sherwood, E.; Sasso, S.; Hodson, S.J.; Warren, M.J.; Smith, A.G. Mutualistic Interactions between Vitamin B12-Dependent Algae and Heterotrophic Bacteria Exhibit Regulation. Environ. Microbiol. 2012, 14, 1466–1476. [Google Scholar] [CrossRef]
- Helliwell, K.E.; Pandhal, J.; Cooper, M.B.; Longworth, J.; Kudahl, U.J.; Russo, D.A.; Tomsett, E.V.; Bunbury, F.; Salmon, D.L.; Smirnoff, N.; et al. Quantitative Proteomics of a B12 -Dependent Alga Grown in Coculture with Bacteria Reveals Metabolic Tradeoffs Required for Mutualism. New Phytol. 2018, 217, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.B.; Kazamia, E.; Helliwell, K.E.; Kudahl, U.J.; Sayer, A.; Wheeler, G.L.; Smith, A.G. Cross-Exchange of B-Vitamins Underpins a Mutualistic Interaction between Ostreococcus Tauri and Dinoroseobacter Shibae. ISME J. 2019, 13, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Z.; Koch, F.; Gobler, C.J. Most Harmful Algal Bloom Species Are Vitamin B1 and B12 Auxotrophs. Proc. Natl. Acad. Sci. USA 2010, 107, 20756–20761. [Google Scholar] [CrossRef]
- Nef, C.; Henry, C.; Nicolau, É.; Bérard, J.-B.; Hervé, F.; Caruana, A.M.N.; Kaas, R.; Mairet, F.; Garnier, M. Cobalamin Scarcity Modifies Carbon Allocation and Impairs DMSP Production Through Methionine Metabolism in the Haptophyte Microalgae Tisochrysis Lutea. Front. Mar. Sci. 2020, 7, 569560. [Google Scholar] [CrossRef]
- Lin, S.; Hu, Z.; Song, X.; Gobler, C.J.; Tang, Y.Z. Vitamin B12-Auxotrophy in Dinoflagellates Caused by Incomplete or Absent Cobalamin-Independent Methionine Synthase Genes (MetE). Fundam. Res. 2022; in press. [Google Scholar] [CrossRef]
- Cruz-López, R.; Maske, H.; Yarimizu, K.; Holland, N.A. The B-Vitamin Mutualism between the Dinoflagellate Lingulodinium polyedrum and the Bacterium Dinoroseobacter shibae. Front. Mar. Sci. 2018, 5, 274. [Google Scholar] [CrossRef]
- Nef, C.; Jung, S.; Mairet, F.; Kaas, R.; Grizeau, D.; Garnier, M. How Haptophytes Microalgae Mitigate Vitamin B12 Limitation. Sci. Rep. 2019, 9, 8417. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.J.; Peloquin, J.A.; Vogt, M.; Heinle, M.; Gruber, N.; Ajani, P.; Andruleit, H.; Arístegui, J.; Beaufort, L.; Estrada, M.; et al. Global Marine Plankton Functional Type Biomass Distributions: Coccolithophores. Earth Syst. Sci. Data 2013, 5, 259–276. [Google Scholar] [CrossRef]
- Sommeria-Klein, G.; Watteaux, R.; Ibarbalz, F.M.; Pierella Karlusich, J.J.; Iudicone, D.; Bowler, C.; Morlon, H. Global Drivers of Eukaryotic Plankton Biogeography in the Sunlit Ocean. Science 2021, 374, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, H.; Buck, K.R.; Chavez, F.P. Haptophytes as Components of Marine Phytoplankton. In The Haptophyte Algae; Clarendon Press: Oxford, UK, 1994; pp. 187–208. [Google Scholar]
- Not, F.; Latasa, M.; Scharek, R.; Viprey, M.; Karleskind, P.; Balagué, V.; Ontoria-Oviedo, I.; Cumino, A.; Goetze, E.; Vaulot, D.; et al. Protistan Assemblages across the Indian Ocean, with a Specific Emphasis on the Picoeukaryotes. Deep Sea Res. Part I Oceanogr. Res. Pap. 2008, 55, 1456–1473. [Google Scholar] [CrossRef]
- Milliman, J.D. Production and Accumulation of Calcium Carbonate in the Ocean: Budget of a Nonsteady State. Glob. Biogeochem. Cycles 1993, 7, 927–957. [Google Scholar] [CrossRef]
- Liu, H.; Probert, I.; Uitz, J.; Claustre, H.; Aris-Brosou, S.; Frada, M.; Not, F.; De Vargas, C. Extreme Diversity in Noncalcifying Haptophytes Explains Major Pigment Paradox in Open Oceans. Proc. Natl. Acad. Sci. USA 2009, 31, 12803–12808. [Google Scholar] [CrossRef]
- Keller, M.D.; Bellows, W.K.; Guillard, R.R.L. Dimethyl Sulfide Production in Marine Phytoplankton. In Biogenic Sulfur in the Environment; Saltzman, E.S., Cooper, W.J., Eds.; American Chemical Society: Washington, DC, USA, 1989; Volume 393, pp. 167–182. ISBN 978-0-8412-1612-9. [Google Scholar]
- Franklin, D.; Steinke, M.; Young, J.; Probert, I.; Malin, G. Dimethylsulphoniopropionate (DMSP), DMSP-Lyase Activity (DLA) and Dimethylsulphide (DMS) in 10 Species of Coccolithophore. Mar. Ecol. Prog. Ser. 2010, 410, 13–23. [Google Scholar] [CrossRef]
- Le Chevanton, M.; Garnier, M.; Bougaran, G.; Schreiber, N.; Lukomska, E.; Bérard, J.-B.; Fouilland, E.; Bernard, O.; Cadoret, J.-P. Screening and Selection of Growth-Promoting Bacteria for Dunaliella Cultures. Algal Res. 2013, 2, 212–222. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Bosi, E.; Donati, B.; Galardini, M.; Brunetti, S.; Sagot, M.-F.; Lió, P.; Crescenzi, P.; Fani, R.; Fondi, M. MeDuSa: A Multi-Draft Based Scaffolder. Bioinformatics 2015, 31, 2443–2451. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An Empirically Improved Memory-Efficient Short-Read de Novo Assembler. GigaScience 2012, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Vallenet, D.; Calteau, A.; Cruveiller, S.; Gachet, M.; Lajus, A.; Josso, A.; Mercier, J.; Renaux, A.; Rollin, J.; Rouy, Z.; et al. MicroScope in 2017: An Expanding and Evolving Integrated Resource for Community Expertise of Microbial Genomes. Nucleic Acids Res. 2017, 45, D517–D528. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Bendif, E.M.; Probert, I.; Schroeder, D.C.; de Vargas, C. On the Description of Tisochrysis Lutea Gen. Nov. Sp. Nov. and Isochrysis Nuda Sp. Nov. in the Isochrysidales, and the Transfer of Dicrateria to the Prymnesiales (Haptophyta). J. Appl. Phycol. 2013, 25, 1763–1776. [Google Scholar] [CrossRef]
- Cho, J.-Y.; Choi, J.-S.; Kong, I.-S.; Park, S.-I.; Kerr, R.G.; Hong, Y.-K. A Procedure for Axenic Isolation of the Marine Microalga Isochrysis Galbana from Heavily Contaminated Mass Cultures. J. Appl. Phycol. 2002, 14, 385–390. [Google Scholar] [CrossRef]
- Walne, P.R. Studies on the Food Value of Nineteen Genera of Algae to Juvenile Bivalves of the Genera Ostrea, Crassostrea, Mercenaria and Mytilus. Fish. Investig. 1970, 26, 62. [Google Scholar]
- Gasol, J.M.; Zweifel, U.L.; Peters, F.; Fuhrman, J.A.; Hagström, Å. Significance of Size and Nucleic Acid Content Heterogeneity as Measured by Flow Cytometry in Natural Planktonic Bacteria. Appl. Environ. Microbiol. 1999, 65, 4475–4483. [Google Scholar] [CrossRef]
- Andersson, A.F.; Lindberg, M.; Jakobsson, H.; Bäckhed, F.; Nyrén, P.; Engstrand, L. Comparative Analysis of Human Gut Microbiota by Barcoded Pyrosequencing. PLoS ONE 2008, 3, e2836. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. Accurate Taxonomy Assignments from 16S RRNA Sequences Produced by Highly Parallel Pyrosequencers. Nucleic Acids Res. 2008, 36, e120. [Google Scholar] [CrossRef] [PubMed]
- Lluch, J.; Servant, F.; Païssé, S.; Valle, C.; Valière, S.; Kuchly, C.; Vilchez, G.; Donnadieu, C.; Courtney, M.; Burcelin, R.; et al. The Characterization of Novel Tissue Microbiota Using an Optimized 16S Metagenomic Sequencing Pipeline. PLoS ONE 2015, 10, e0142334. [Google Scholar] [CrossRef] [PubMed]
- Cormier, A.; Durand, P.; Noel, C.; Leroi, L. SAMBA: Standardized and Automated MetaBarcoding Analyses Workflow. 2021. Available online: https://github.com/ifremer-bioinformatics/samba (accessed on 5 May 2022).
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Di Tommaso, P.; Chatzou, M.; Floden, E.W.; Barja, P.P.; Palumbo, E.; Notredame, C. Nextflow Enables Reproducible Computational Workflows. Nat. Biotechnol. 2017, 35, 316–319. [Google Scholar] [CrossRef]
- Olesen, S.W.; Duvallet, C.; Alm, E.J. DbOTU3: A New Implementation of Distribution-Based OTU Calling. PLoS ONE 2017, 12, e0176335. [Google Scholar] [CrossRef]
- McKnight, D.T.; Huerlimann, R.; Bower, D.S.; Schwarzkopf, L.; Alford, R.A.; Zenger, K.R. MicroDecon: A Highly Accurate Read-subtraction Tool for the Post-sequencing Removal of Contamination in Metabarcoding Studies. Environ. DNA 2019, 1, 14–25. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Glöckner, F.O.; Yilmaz, P.; Quast, C.; Gerken, J.; Beccati, A.; Ciuprina, A.; Bruns, G.; Yarza, P.; Peplies, J.; Westram, R.; et al. 25 Years of Serving the Community with Ribosomal RNA Gene Reference Databases and Tools. J. Biotechnol. 2017, 261, 169–176. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for Prediction of Metagenome Functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Ragsdale, S.W. The Many Faces of Vitamin B12: Catalysis by Cobalamin-Dependent Enzymes. Annu. Rev. Biochem. 2003, 72, 209–247. [Google Scholar] [CrossRef] [PubMed]
- Rodionov, D.A.; Vitreschak, A.G.; Mironov, A.A.; Gelfand, M.S. Comparative Genomics of Vitamin B12 Metabolism and Regulation in Prokaryotes. J. Biol. Chem. 2003, 42, 41148–41159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Rodionov, D.A.; Gelfand, M.S.; Gladyshev, V.N. Comparative Genomic Analyses of Nickel, Cobalt and Vitamin B12 Utilization. BMC Genom. 2009, 10, 78. [Google Scholar] [CrossRef] [PubMed]
- Swithers, K.S.; Petrus, A.K.; Secinaro, M.A.; Nesbø, C.L.; Gogarten, J.P.; Noll, K.M.; Butzin, N.C. Vitamin B12 Synthesis and Salvage Pathways Were Acquired by Horizontal Gene Transfer to the Thermotogales. Genome Biol. Evol. 2012, 4, 842–851. [Google Scholar] [CrossRef]
- Sañudo-Wilhelmy, S.A.; Gómez-Consarnau, L.; Suffridge, C.; Webb, E.A. The Role of B Vitamins in Marine Biogeochemistry. Annu. Rev. Mar. Sci. 2014, 6, 339–367. [Google Scholar] [CrossRef]
- de Veaux, L.C.; Clevenson, D.S.; Bradbeer, C.; Kadner, R.J. Identification of the BtuCED Polypeptides and Evidence for Their Role in Vitamin B12 Transport in Escherichia Coli. J. Bacteriol. 1986, 167, 920–927. [Google Scholar] [CrossRef]
- Suffridge, C.P.; Gómez-Consarnau, L.; Monteverde, D.R.; Cutter, L.; Arístegui, J.; Alvarez-Salgado, X.A.; Gasol, J.M.; Sañudo-Wilhelmy, S.A. B Vitamins and Their Congeners as Potential Drivers of Microbial Community Composition in an Oligotrophic Marine Ecosystem. J. Geophys. Res. Biogeosci. 2018, 123, 2890–2907. [Google Scholar] [CrossRef]
- Sokolovskaya, O.M.; Shelton, A.N.; Taga, M.E. Sharing Vitamins: Cobamides Unveil Microbial Interactions. Science 2020, 369, eaba0165. [Google Scholar] [CrossRef]
- Jardillier, L.; Zubkov, M.V.; Pearman, J.; Scanlan, D.J. Significant CO2 Fixation by Small Prymnesiophytes in the Subtropical and Tropical Northeast Atlantic Ocean. ISME J. 2010, 4, 1180–1192. [Google Scholar] [CrossRef]
- Segev, E.; Wyche, T.P.; Kim, K.H.; Petersen, J.; Ellebrandt, C.; Vlamakis, H.; Barteneva, N.; Paulson, J.N.; Chai, L.; Clardy, J.; et al. Dynamic Metabolic Exchange Governs a Marine Algal-Bacterial Interaction. eLife 2016, 5, e17473. [Google Scholar] [CrossRef] [PubMed]
- Barak-Gavish, N.; Frada, M.J.; Ku, C.; Lee, P.A.; DiTullio, G.R.; Malitsky, S.; Aharoni, A.; Green, S.J.; Rotkopf, R.; Kartvelishvily, E.; et al. Bacterial Virulence against an Oceanic Bloom-Forming Phytoplankter Is Mediated by Algal DMSP. Sci. Adv. 2018, 4, eaau5716. [Google Scholar] [CrossRef] [PubMed]
- Chrétiennot-Dinet, M.-J.; Courties, C.; Vaquer, A.; Neveux, J.; Claustre, H.; Lautier, J.; Machado, M.C. A New Marine Picoeucaryote: Ostreococcus Tauri Gen. et Sp. Nov. (Chlorophyta, Prasinophyceae). Phycologia 1995, 34, 285–292. [Google Scholar] [CrossRef]
- Lewis, J.; Hallett, R. Lingulodinium Polyedrum (Gonyaulax Polyedra) a Blooming Dinoflagellate. Oceanogr. Mar. Biol. Annu. Rev. 1997, 35, 97–161. [Google Scholar]
- Marañón, E.; Cermeño, P.; López-Sandoval, D.C.; Rodríguez-Ramos, T.; Sobrino, C.; Huete-Ortega, M.; Blanco, J.M.; Rodríguez, J. Unimodal Size Scaling of Phytoplankton Growth and the Size Dependence of Nutrient Uptake and Use. Ecol. Lett. 2013, 16, 371–379. [Google Scholar] [CrossRef]
- Ridley, C.J.A.; Day, J.G.; Smith, A.G. Cryopreservation Studies of an Artificial Co-Culture between the Cobalamin-Requiring Green Alga Lobomonas Rostrata and the Bacterium Mesorhizobium Loti. J. Appl. Phycol. 2018, 30, 995–1003. [Google Scholar] [CrossRef]
- Strenkert, D.; Schmollinger, S.; Gallaher, S.D.; Salomé, P.A.; Purvine, S.O.; Nicora, C.D.; Mettler-Altmann, T.; Soubeyrand, E.; Weber, A.P.M.; Lipton, M.S.; et al. Multiomics Resolution of Molecular Events during a Day in the Life of Chlamydomonas. Proc. Natl. Acad. Sci. USA 2019, 116, 2374–2383. [Google Scholar] [CrossRef]
- Droop, M.R. Vitamins, Phytoplankton and Bacteria: Symbiosis or Scavenging? J. Plankton Res. 2007, 29, 107–113. [Google Scholar] [CrossRef]
- Panzeca, C.; Beck, A.J.; Tovar-Sanchez, A.; Segovia-Zavala, J.; Taylor, G.T.; Gobler, C.J.; Sañudo-Wilhelmy, S.A. Distributions of Dissolved Vitamin B12 and Co in Coastal and Open-Ocean Environments. Estuar. Coast. Shelf Sci. 2009, 85, 223–230. [Google Scholar] [CrossRef]
- Fiala, M.; Oriol, L. Vitamine B12 et phytoplancton dans l’Océan Antarctique. Distribution et approche expérimentale. Mar. Biol. 1984, 79, 325–332. [Google Scholar] [CrossRef]
- Menzel, D.W.; Spaeth, J.P. Occurrence of Vitamin B12 in the Sargasso Sea. Limnol. Oceanogr. 1962, 7, 151–154. [Google Scholar] [CrossRef]
- Bertrand, E.M.; Moran, D.M.; McIlvin, M.R.; Hoffman, J.M.; Allen, A.E.; Saito, M.A. Methionine Synthase Interreplacement in Diatom Cultures and Communities: Implications for the Persistence of B12 Use by Eukaryotic Phytoplankton. Limnol. Oceanogr. 2013, 58, 1431–1450. [Google Scholar] [CrossRef]
- Seyedsayamdost, M.R.; Carr, G.; Kolter, R.; Clardy, J. Roseobacticides: Small Molecule Modulators of an Algal-Bacterial Symbiosis. J. Am. Chem. Soc. 2011, 133, 18343–18349. [Google Scholar] [CrossRef] [PubMed]
- Seyedsayamdost, M.R.; Case, R.J.; Kolter, R.; Clardy, J. The Jekyll-and-Hyde Chemistry of Phaeobacter Gallaeciensis. Nat. Chem. 2011, 3, 331–335. [Google Scholar] [CrossRef]
- Helliwell, K.E.; Lawrence, A.D.; Holzer, A.; Kudahl, U.J.; Sasso, S.; Kräutler, B.; Scanlan, D.J.; Warren, M.J.; Smith, A.G. Cyanobacteria and Eukaryotic Algae Use Different Chemical Variants of Vitamin B12. Curr. Biol. 2016, 26, 999–1008. [Google Scholar] [CrossRef]
- Shelford, E.J.; Suttle, C.A. Virus-Mediated Transfer of Nitrogen from Heterotrophic Bacteria to Phytoplankton. Biogeosciences 2018, 15, 809–819. [Google Scholar] [CrossRef]
- Anderson, R.; Charvet, S.; Hansen, P.J. Mixotrophy in Chlorophytes and Haptophytes—Effect of Irradiance, Macronutrient, Micronutrient and Vitamin Limitation. Front. Microbiol. 2018, 9, 1704. [Google Scholar] [CrossRef]
- Legrand, C. Phagotrophy and Toxicity Variation in the Mixotrophic Prymnesium Patelliferum (Haptophyceae). Limnol. Oceanogr. 2001, 46, 1208–1214. [Google Scholar] [CrossRef]
- Unrein, F.; Gasol, J.M.; Not, F.; Forn, I.; Massana, R. Mixotrophic Haptophytes Are Key Bacterial Grazers in Oligotrophic Coastal Waters. ISME J. 2014, 8, 164–176. [Google Scholar] [CrossRef]
- Liu, Z.; Campbell, V.; Heidelberg, K.B.; Caron, D.A. Gene Expression Characterizes Different Nutritional Strategies among Three Mixotrophic Protists. FEMS Microbiol. Ecol. 2016, 92, fiw106. [Google Scholar] [CrossRef]
- Nef, C. Métabolisme et Interactions Bactériennes En Lien Avec La Vitamine B12 Chez La Microalgue Haptophyte Tisochrysis Lutea; Université de Nantes: Nantes, France, 2019. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Species | Origin |
|---|---|---|
| 09-029 | Phaeobacter sp. | Odontella aurita and Emiliania huxleyi [31] |
| 09-003 | Halomonas spp. | |
| 09-027 | ||
| 09-213 | ||
| 09-729 | ||
| 1211 | Halomonas spp. | Deep-sea ecosystems |
| 1212 | ||
| 1229 | ||
| 1233 | ||
| 1236 | ||
| 19-001 | Halomonas sp. | Amphidinium operculatum [4] |
| Culture Condition | ΔCmax (106 Cells/mL) |
|---|---|
| Axenic − B12 | 0.86 ± 0.10 |
| Axenic + B12 | 9.73 ± 0.22 |
| 19-001 − B12 | 0.97 ± 0.06 |
| 19-001 + B12 | 10.23 ± 0.21 |
| 09-029 − B12 | 0.98 ± 0.12 |
| 09-029 + B12 | 10.03 ± 0.65 |
| 09-029 extract | 6.81 ± 1.59 |
| 09-729 extract | 0.96 ± 1.35 |
| Culture Condition | ΔCmax (106 Cells/mL) |
|---|---|
| Axenic − B12 | 0.69 ± 0.07 |
| Axenic + B12 | 24.86 ± 2.16 |
| Xenic − B12 | 4.54 ± 0.72 |
| Xenic + B12 | 21.37 ± 0.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nef, C.; Dittami, S.; Kaas, R.; Briand, E.; Noël, C.; Mairet, F.; Garnier, M. Sharing Vitamin B12 between Bacteria and Microalgae Does Not Systematically Occur: Case Study of the Haptophyte Tisochrysis lutea. Microorganisms 2022, 10, 1337. https://doi.org/10.3390/microorganisms10071337
Nef C, Dittami S, Kaas R, Briand E, Noël C, Mairet F, Garnier M. Sharing Vitamin B12 between Bacteria and Microalgae Does Not Systematically Occur: Case Study of the Haptophyte Tisochrysis lutea. Microorganisms. 2022; 10(7):1337. https://doi.org/10.3390/microorganisms10071337
Chicago/Turabian StyleNef, Charlotte, Simon Dittami, Raymond Kaas, Enora Briand, Cyril Noël, Francis Mairet, and Matthieu Garnier. 2022. "Sharing Vitamin B12 between Bacteria and Microalgae Does Not Systematically Occur: Case Study of the Haptophyte Tisochrysis lutea" Microorganisms 10, no. 7: 1337. https://doi.org/10.3390/microorganisms10071337
APA StyleNef, C., Dittami, S., Kaas, R., Briand, E., Noël, C., Mairet, F., & Garnier, M. (2022). Sharing Vitamin B12 between Bacteria and Microalgae Does Not Systematically Occur: Case Study of the Haptophyte Tisochrysis lutea. Microorganisms, 10(7), 1337. https://doi.org/10.3390/microorganisms10071337