
Construction of HGF-Displaying Yeast by Cell Surface Engineering

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain and Media
2.2. Plasmid Construction and Transformation of Yeast Cell
2.3. Immunostaining
2.4. Immunofluorescence Observation
3. Results and Discussion
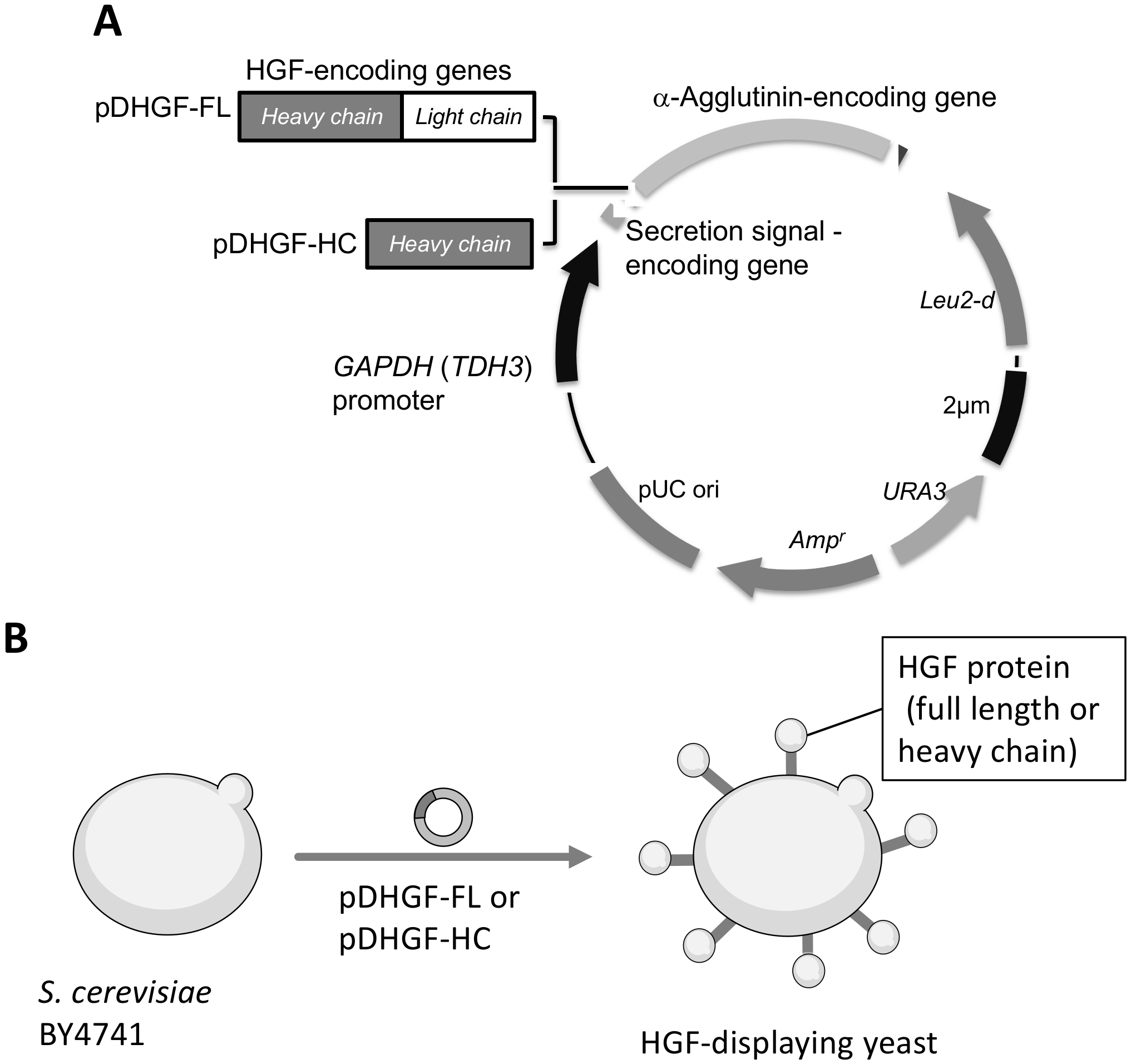
3.1. Plasmid Construction and HGF-Displaying Yeast
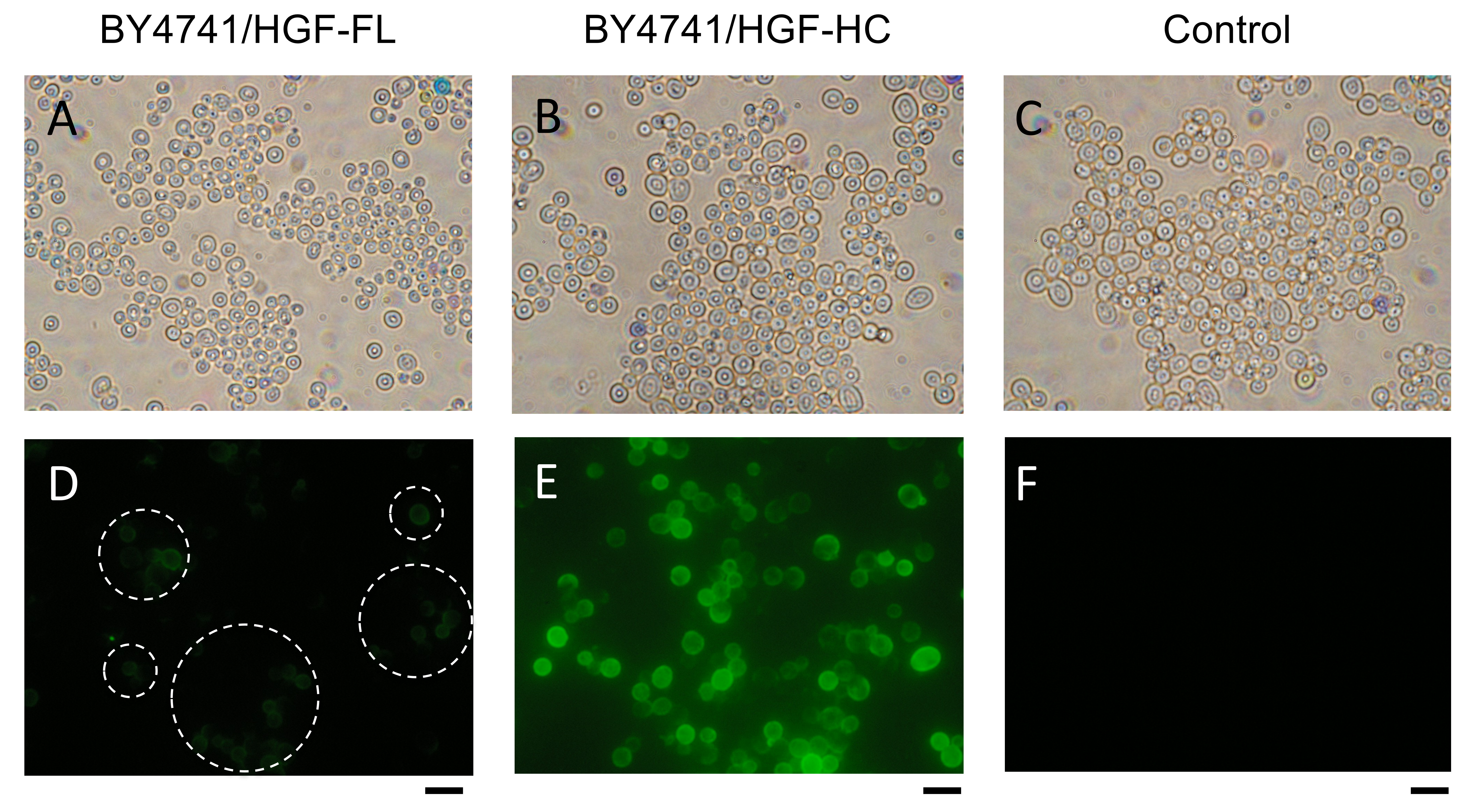
3.2. Immunofluorescence Observation
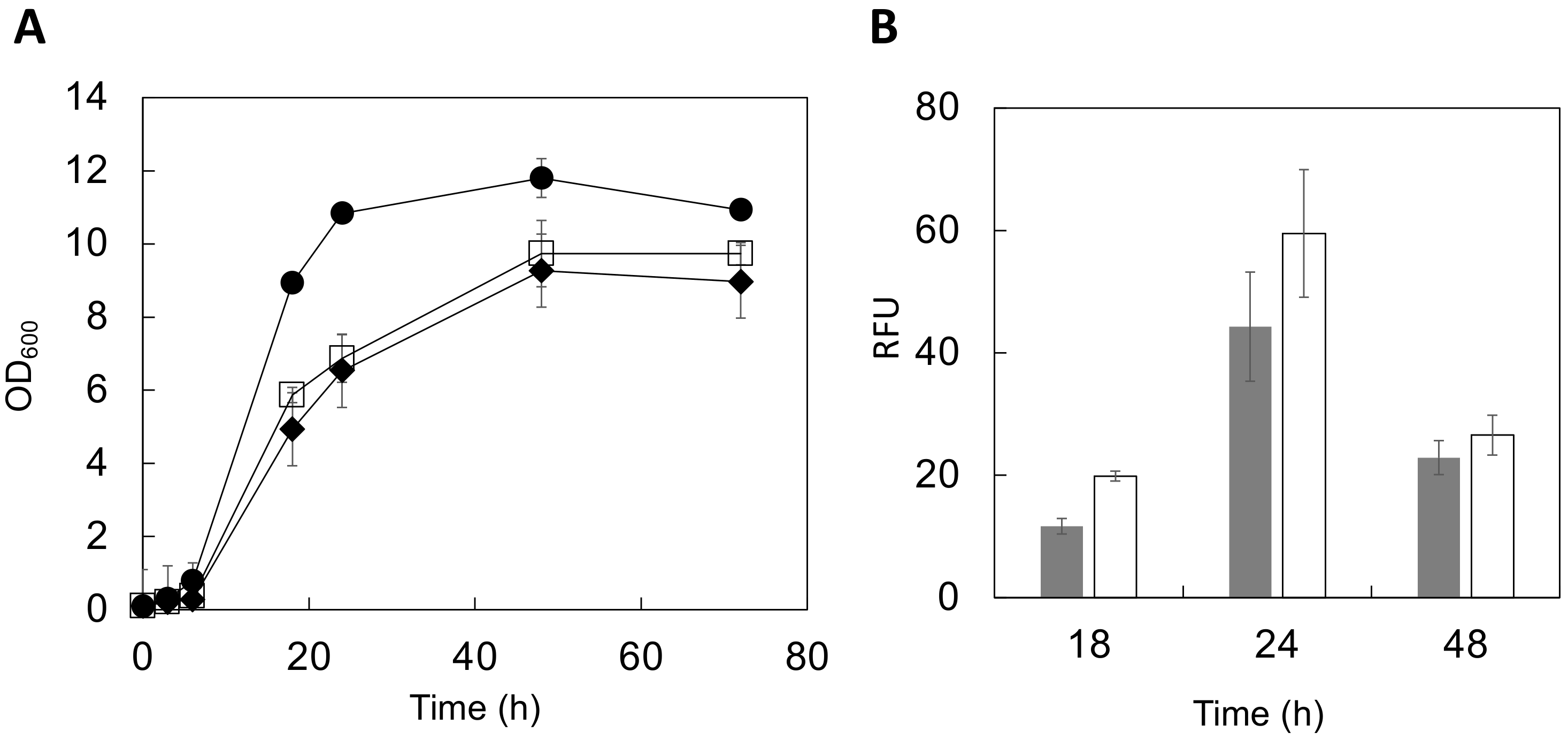
3.3. Cultivation Conditions of HGF-Displaying Yeast
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Miyazawa, K.; Shimomura, T.; Kitamura, A.; Kondo, J.; Morimoto, Y.; Kitamura, N. Molecular Cloning and Sequence Analysis of the cDNA for a Human Serine Protease Responsible for Activation of Hepatocyte Growth Factor. Structural Similarity of the Protease Precursor to Blood Coagulation Factor XII. J. Biol. Chem. 1993, 268, 10024–10028. [Google Scholar] [CrossRef]
- Gallo, S.; Sala, V.; Gatti, S.; Crepaldi, T. Cellular and Molecular Mechanisms of HGF/Met in the Cardiovascular System. Clin. Sci. 2015, 129, 1173–1193. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.; Malhotra, S.S.; Babu, G.S.; Sobti, R.C.; Gupta, S.K. HGF Promotes HTR-8/SVneo Cell Migration Through Activation of MAPK/PKA Signaling Leading to Up-Regulation of WNT Ligands and Integrins That Target β-Catenin. Mol. Cell. Biochem. 2019, 453, 11–32. [Google Scholar] [CrossRef] [PubMed]
- Inampudi, C.; Akintoye, E.; Ando, T.; Briasoulis, A. Angiogenesis in Peripheral Arterial Disease. Curr. Opin. Pharmacol. 2018, 39, 60–67. [Google Scholar] [CrossRef]
- Yamashita, T.; Abe, K. Recent Progress in Therapeutic Strategies for Ischemic Stroke. Cell. Transplant. 2016, 25, 893–898. [Google Scholar] [CrossRef]
- Ido, A.; Moriuchi, A.; Numata, M.; Murayama, T.; Teramukai, S.; Marusawa, H.; Yamaji, N.; Setoyama, H.; Kim, I.D.; Chiba, T.; et al. Safety and Pharmacokinetics of Recombinant Human Hepatocyte Growth Factor (Rh-HGF) in Patients with Fulminant Hepatitis: A phase I/II Clinical Trial, Following Preclinical Studies to Ensure Safety. J. Transl. Med. 2011, 9, 55. [Google Scholar] [CrossRef]
- Matsumoto, K.; Nakamura, T. Hepatocyte Growth Factor: Renotropic Role and Potential Therapeutics for Renal Diseases. Kidney Int. 2001, 59, 2023–2038. [Google Scholar] [CrossRef]
- Moon, S.H.; Lee, C.M.; Park, S.H.; Jin Nam, M. Effects of Hepatocyte Growth Factor Gene-Transfected Mesenchymal Stem Cells on Dimethylnitrosamine-Induced Liver Fibrosis in Rats. Growth Factors 2019, 37, 105–119. [Google Scholar] [CrossRef]
- Yamagami, T.; Kanda, K.; Okuyama, C.; Nishimura, T. Tc-99M-MIBI Scintigraphy in Evaluating the Effect of Hepatocyte Growth Factor Gene Therapy for Peripheral Arteriosclerosis Obliterans. Ann. Nucl. Med. 2009, 23, 205–208. [Google Scholar] [CrossRef]
- Azuma, J.; Taniyama, Y.; Takeya, Y.; Iekushi, K.; Aoki, M.; Dosaka, N.; Matsumoto, K.; Nakamura, T.; Ogihara, T.; Morishita, R. Angiogenic and Antifibrotic Actions of Hepatocyte Growth Factor Improve Cardiac Dysfunction in Porcine Ischemic Cardiomyopathy. Gene. Ther. 2006, 13, 1206–1213. [Google Scholar] [CrossRef]
- Kuroiwa, T.; Iwasaki, T.; Imado, T.; Sekiguchi, M.; Fujimoto, J.; Sano, H. Hepatocyte Growth Factor Prevents Lupus Nephritis in a Murine Lupus Model of Chronic Graft-Versus-Host Disease. Arthritis Res. Ther. 2006, 8, R123. [Google Scholar] [CrossRef] [PubMed]
- Imado, T.; Iwasaki, T.; Kataoka, Y.; Kuroiwa, T.; Hara, H.; Fujimoto, J.; Sano, H. Hepatocyte Growth Factor Preserves Graft-Versus-Leukemia Effect and T-Cell Reconstitution After Marrow Transplantation. Blood 2004, 104, 1542–1549. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.B.; Lee, S.J.; Schubert, M.M. Graft-Vs.-Host Disease. Crit. Rev. Oral. Biol. Med. 1997, 8, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Nassereddine, S.; Rafei, H.; Elbahesh, E.; Tabbara, I. Acute Graft Versus Host Disease: A Comprehensive Review. Anticancer. Res. 2017, 37, 1547–1555. [Google Scholar] [CrossRef] [PubMed]
- Kuroiwa, T.; Kakishita, E.; Hamano, T.; Kataoka, Y.; Seto, Y.; Iwata, N.; Kaneda, Y.; Matsumoto, K.; Nakamura, T.; Ueki, T.; et al. Hepatocyte Growth Factor Ameliorates Acute Graft-Versus-Host Disease and Promotes Hematopoietic Function. J. Clin. Investig. 2001, 107, 1365–1373. [Google Scholar] [CrossRef]
- Hartmann, G.; Naldini, L.; Weidner, K.M.; Sachs, M.; Vigna, E.; Comoglio, P.M.; Birchmeier, W. A Functional Domain in the Heavy Chain of Scatter Factor/Hepatocyte Growth Factor Binds the c-Met Receptor and Induces Cell Dissociation but Not Mitogenesis. Proc. Natl. Acad. Sci. USA 1992, 89, 11574–11578. [Google Scholar] [CrossRef]
- Iwasaki, T.; Shibasaki, S. Hepatocyte Growth Factor Regulates Immune Reactions Caused by Transplantation and Autoimmune Diseases. Yakugaku Zasshi 2013, 133, 1159–1167. [Google Scholar] [CrossRef][Green Version]
- Suzuki, Y.; Sakai, K.; Ueki, J.; Xu, Q.; Nakamura, T.; Shimada, H.; Nakamura, T.; Matsumoto, K. Inhibition of Met/HGF Receptor and Angiogenesis by NK4 Leads to Suppression of Tumor Growth and Migration in Malignant Pleural Mesothelioma. Int. J. Cancer 2010, 127, 1948–1957. [Google Scholar] [CrossRef]
- Shibasaki, S.; Kitano, S.; Karasaki, M.; Tsunemi, S.; Sano, H.; Iwasaki, T. Blocking c-Met Signaling Enhances Bone Morphogenetic protein-2-Induced Osteoblast Differentiation. FEBS Open Bio 2015, 5, 341–347. [Google Scholar] [CrossRef]
- Shibasaki, S.; Ueda, M. Bioadsorption Strategies with Yeast Molecular Display Technology. Biocontrol Sci. 2014, 19, 157–164. [Google Scholar] [CrossRef]
- Shibasaki, S.; Maeda, H.; Ueda, M. Molecular Display Technology Using Yeast—Arming Technology. Anal. Sci. 2009, 25, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, S.; Ueda, M. Development of yeast molecular display systems focused on therapeutic proteins, enzymes, and foods: Functional analysis of proteins and its application to bioconversion. Recent Pat. Biotechnol. 2010, 4, 198–213. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, S.; Aoki, W.; Nomura, T.; Miyoshi, A.; Tafuku, S.; Sewaki, T.; Ueda, M. An Oral Vaccine against Candidiasis Generated by a Yeast Molecular Display System. Pathog. Dis. 2013, 69, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, S.; Karasaki, M.; Tafuku, S.; Aoki, W.; Sewaki, T.; Ueda, M. Oral Immunization Against Candidiasis Using Lactobacillus casei Displaying Enolase 1 from Candida albicans. Sci. Pharm. 2014, 82, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Shigemori, T.; Nagayama, M.; Yamada, J.; Miura, N.; Yongkiettrakul, S.; Kuroda, K.; Katsuragi, T.; Ueda, M. Construction of a Convenient System for Easily Screening Inhibitors of Mutated Influenza Virus Neuraminidases. FEBS Open Bio 2013, 3, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Adachi, K.; Kawana, K.; Yokoyama, T.; Fujii, T.; Tomio, A.; Miura, S.; Tomio, K.; Kojima, S.; Oda, K.; Sewaki, T.; et al. Oral Immunization with a Lactobacillus casei Vaccine Expressing Human Papillomavirus (HPV) type 16 E7 Is an Effective Strategy to Induce Mucosal Cytotoxic Lymphocytes Against HPV16 E7. Vaccine 2010, 28, 2810–2817. [Google Scholar] [CrossRef] [PubMed]
- Kajiwara, K.; Aoki, W.; Koike, N.; Ueda, M. Development of a Yeast Cell Surface Display Method Using the SpyTag/SpyCatcher System. Sci. Rep. 2021, 11, 11059. [Google Scholar] [CrossRef]
- Boyd, J.; Oza, M.N.; Murphy, J.R. Molecular cloning and DNA sequence analysis of a diphtheria tox iron-dependent regulatory element (dtxR) from Corynebacterium diphtheriae. Proc. Natl. Acad. Sci. USA 1990, 87, 5968–5972. [Google Scholar] [CrossRef]
- Kuroda, K.; Matsui, K.; Higuchi, S.; Kotaka, A.; Sahara, H.; Hata, Y.; Ueda, M. Enhancement of display efficiency in yeast display system by vector engineering and gene disruption. Appl. Microbiol. Biotechnol. 2009, 82, 713–719. [Google Scholar] [CrossRef]
- Ito, H.; Fukuda, Y.; Murata, K.; Kimura, A. Transformation of Intact Yeast Cells Treated with Alkali Cations. J. Bacteriol. 1983, 153, 163–168. [Google Scholar] [CrossRef]
- Shibasaki, S.; Kawabata, A.; Ishii, J.; Yagi, S.; Kadonosono, T.; Kato, M.; Fukuda, N.; Kondo, A.; Ueda, M. Construction of a Novel Synergistic System for Production and Recovery of Secreted Recombinant Proteins by the Cell Surface Engineering. Appl. Microbiol. Biotechnol. 2007, 75, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Tsubouchi, H.; Naka, D.; Takahashi, K.; Okigaki, M.; Arakaki, N.; Nakayama, H.; Hirono, S.; Sakiyama, O.; Takahashi, K.; et al. Molecular Cloning and Sequence Analysis of cDNA for Human Hepatocyte Growth Factor. Biochem. Biophys. Res. Commun. 1989, 163, 967–973. [Google Scholar] [CrossRef]
- Shibasaki, S.; Tanaka, A.; Ueda, M. Development of Combinatorial Bioengineering Using Yeast Cell Surface Display-Order-Made Design of Cell and Protein for Bio-Monitoring. Biosens. Bioelectron. 2003, 19, 123–130. [Google Scholar] [CrossRef]
- Shibasaki, S.; Ueda, M.; Iizuka, T.; Hirayama, M.; Ikeda, Y.; Kamasawa, N.; Osumi, M.; Tanaka, A. Quantitative Evaluation of the Enhanced Green Fluorescent Protein Displayed on the Cell Surface of Saccharomyces cerevisiae by Fluorometric and Confocal Laser Scanning Microscopic Analyses. Appl. Microbiol. Biotechnol. 2001, 55, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Yasui, M.; Shibasaki, S.; Kuroda, K.; Ueda, M.; Kawada, N.; Nishikawa, J.; Nishihara, T.; Tanaka, A. An Arming Yeast with the Ability to Entrap Fluorescent 17Beta-Estradiol on the Cell Surface. Appl. Microbiol. Biotechnol. 2002, 59, 329–331. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shibasaki, S.; Nakatani, Y.; Taketani, K.; Karasaki, M.; Matsui, K.; Ueda, M.; Iwasaki, T. Construction of HGF-Displaying Yeast by Cell Surface Engineering. Microorganisms 2022, 10, 1373. https://doi.org/10.3390/microorganisms10071373
Shibasaki S, Nakatani Y, Taketani K, Karasaki M, Matsui K, Ueda M, Iwasaki T. Construction of HGF-Displaying Yeast by Cell Surface Engineering. Microorganisms. 2022; 10(7):1373. https://doi.org/10.3390/microorganisms10071373
Chicago/Turabian StyleShibasaki, Seiji, Yuki Nakatani, Kazuaki Taketani, Miki Karasaki, Kiyoshi Matsui, Mitsuyoshi Ueda, and Tsuyoshi Iwasaki. 2022. "Construction of HGF-Displaying Yeast by Cell Surface Engineering" Microorganisms 10, no. 7: 1373. https://doi.org/10.3390/microorganisms10071373
APA StyleShibasaki, S., Nakatani, Y., Taketani, K., Karasaki, M., Matsui, K., Ueda, M., & Iwasaki, T. (2022). Construction of HGF-Displaying Yeast by Cell Surface Engineering. Microorganisms, 10(7), 1373. https://doi.org/10.3390/microorganisms10071373