
Development of a Real-Time Quantitative PCR Assay for the Specific Detection of Bacillus velezensis and Its Application in the Study of Colonization Ability



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Culture Conditions and DNA Extraction
2.2. Design and Selection of Species-Specific Primers
2.3. Primer Specificity Verification and Real-Time qPCR Assays
2.4. Standard Curve Determination and Sensitivity Test
2.5. Colonization Ability of B. velezensis ZF2 in Different Soils
2.6. Determination of Physicochemical Properties of the Different Soils
2.7. Colonization Ability of B. velezensis ZF2 under Different Environmental Conditions (pH and Nutrient Elements)
3. Results
3.1. Establishment of Specific Detection System
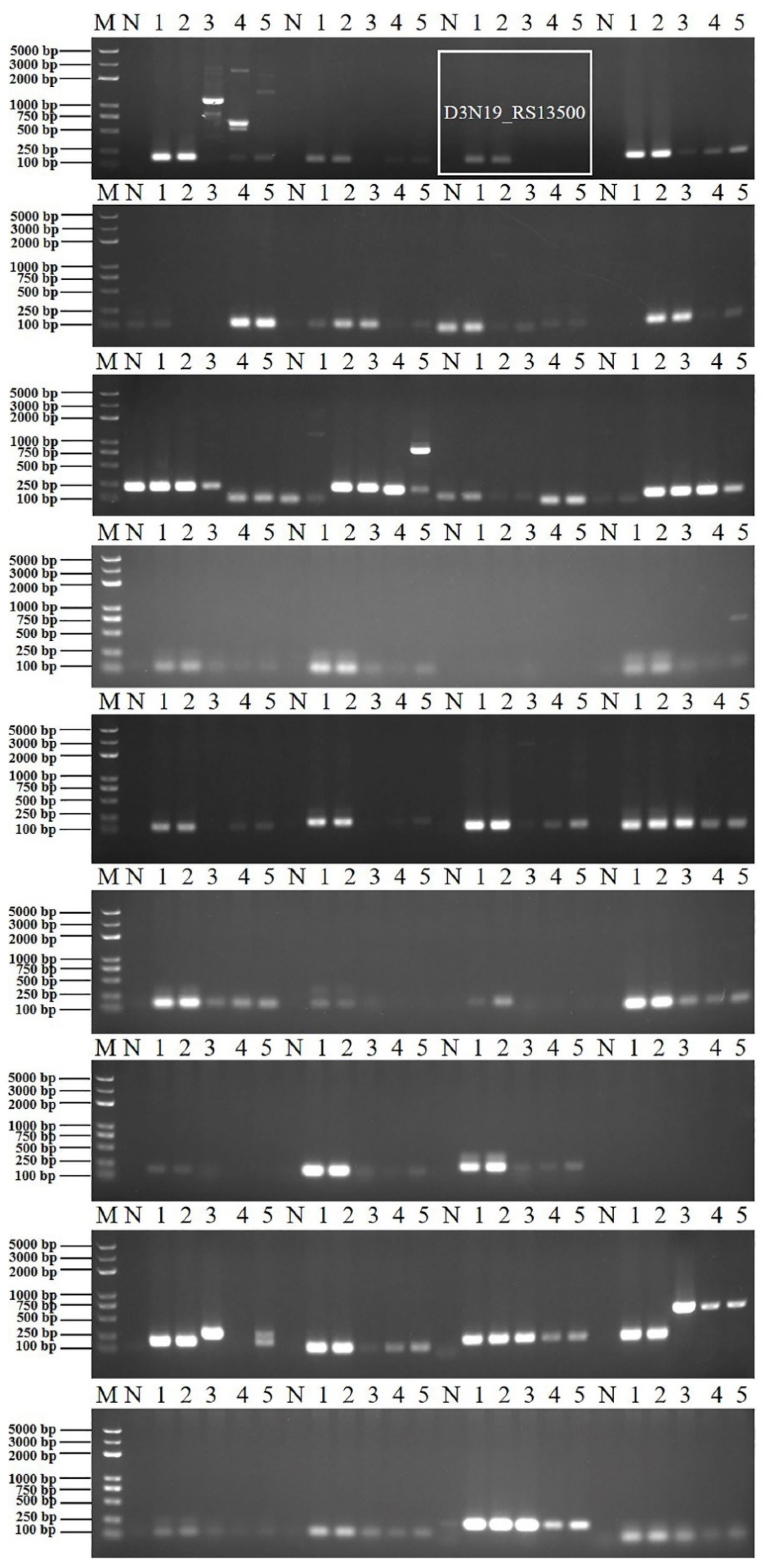
3.1.1. Design and Selection of Specific Primers
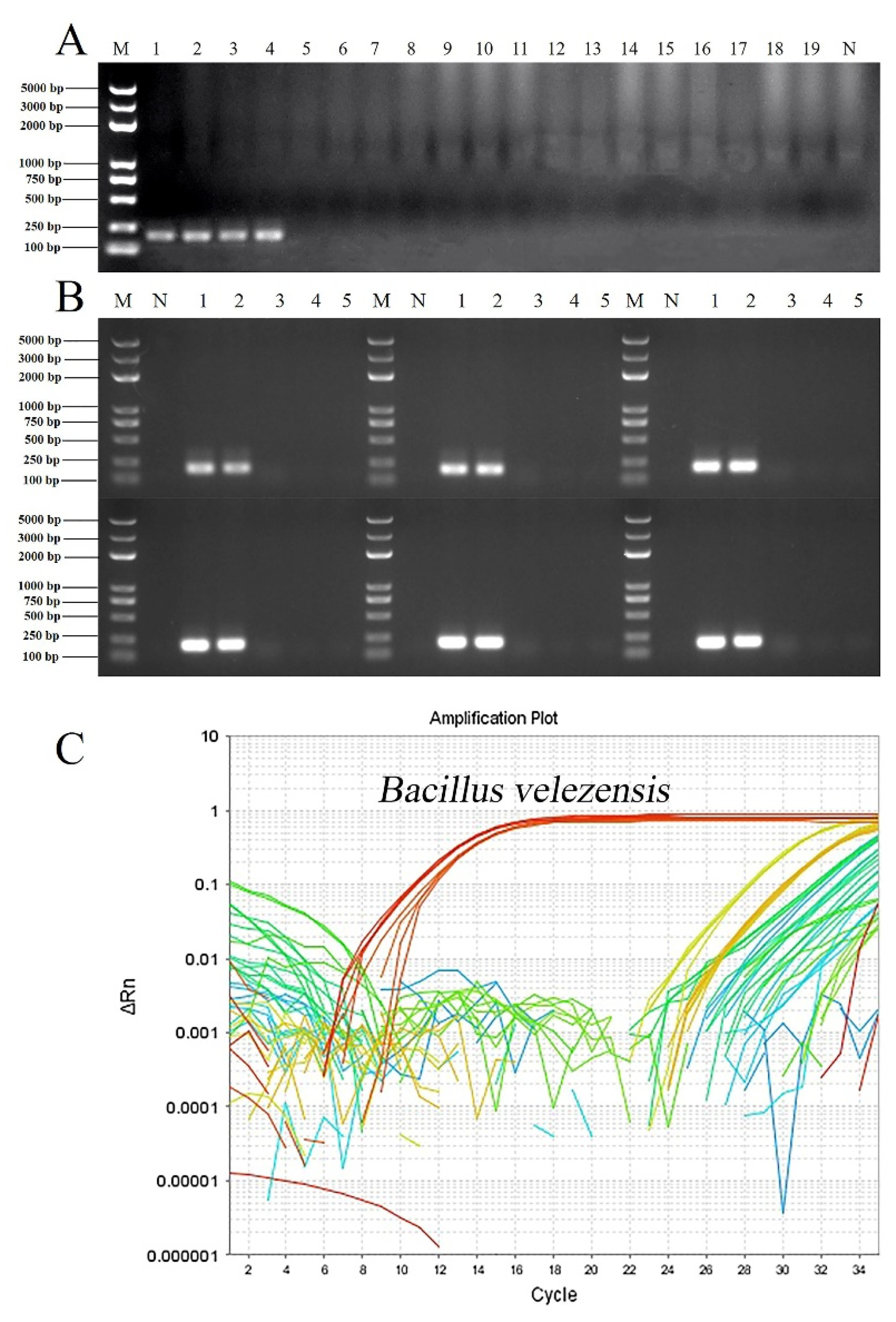
3.1.2. Verification of Primer Specificity and Optimization of PCR Reaction Conditions
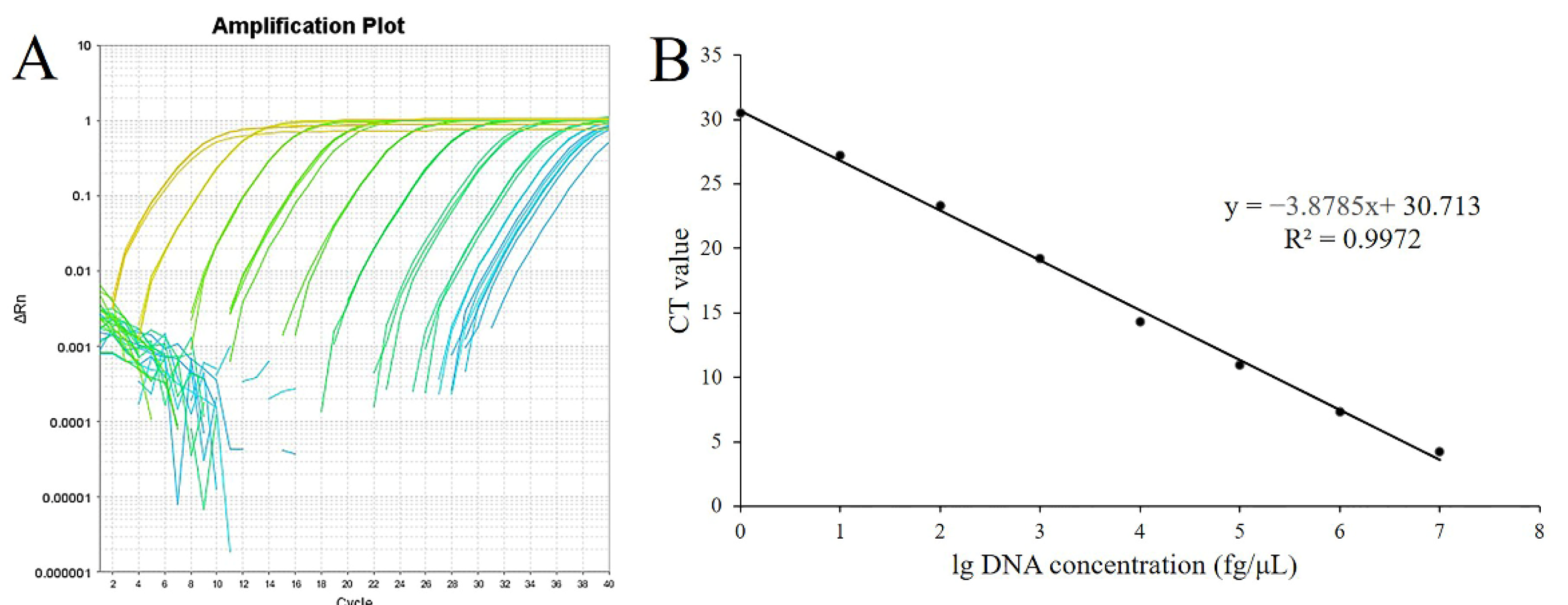
3.1.3. Standard Curve Determination and Detection Limit
3.2. Detection of the Colonization Ability of B. velezensis ZF2
3.2.1. Colonization Ability of B. velezensis ZF2 in Different Soils
3.2.2. Determination of Soil Physicochemical Properties
3.2.3. Colonization Ability of B. velezensis ZF2 under Different Environmental Conditions (pH)
3.2.4. Colonization Ability of B. velezensis ZF2 under Different Nutrient Additions (Carbon Source, Nitrogen Source, Inorganic Compounds)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruiz-García, C.; Béjar, V.; Martínez-Checa, F.; Llamas, I.; Quesada, E. Bacillus velezensis sp. nov., a surfactant-producing bacterium isolated from the river Vélez in Málaga, southern Spain. Int. J. Syst. Evol. Microbiol. 2005, 55, 191–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.T.; Lee, F.L.; Tai, C.J.; Kuo, H.P. Bacillus velezensis is a later heterotypic synonym of Bacillus amyloliquefaciens. Int. J. Syst. Evol. Microbiol. 2008, 58, 671–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunlap, C.A.; Kim, S.J.; Kwon, S.W.; Rooney, A.P. Bacillus velezensis is not a later heterotypic synonym of Bacillus amyloliquefaciens; Bacillus methylotrophicus, Bacillus amyloliquefaciens subsp. plantarum and ‘Bacillus oryzicola’ are later heterotypic synonyms of Bacillus velezensis based on phylogenomics. Int. J. Syst. Evol. Microbiol. 2016, 66, 1212–1217. [Google Scholar] [CrossRef] [PubMed]
- Johansson, A.H.; Bejai, S.; Niazi, A.; Manzoor, S.; Bongcam-Rudloff, E.; Meijer, J. Studies of plant colonisation by closely related Bacillus amyloliquefaciens biocontrol agents using strain specific quantitative PCR assays. Antonie Van Leeuwenhoek 2014, 106, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ma, Y.; Jiang, W.; Meng, L.; Cao, X.; Hu, J.; Chen, J.; Li, J. Development of a Strain-Specific Quantification Method for Monitoring Bacillus amyloliquefaciens TF28 in the Rhizospheric Soil of Soybean. Mol. Biotechnol. 2020, 62, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Tan, F.; Xu, A.; Deng, K.; Zeng, Y.; Huang, H. UV-induced peroxidase-like activity of gold nanoclusters for differentiating pathogenic bacteria and detection of enterotoxin with colorimetric readout. Sens. Actuators 2019, 279, 289–297. [Google Scholar] [CrossRef]
- Luo, C.; Chen, Y.; Liu, X.; Wang, X.; Wang, X.; Li, X.; Zhao, Y.; Wei, L. Engineered biosynthesis of cyclic lipopeptide locillomycins in surrogate host Bacillus velezensis FZB42 and derivative strains enhance antibacterial activity. Appl. Microbiol. Biotechnol. 2019, 103, 4467–4481. [Google Scholar] [CrossRef]
- Sun, X.; Xu, Z.; Xie, J.; Hesselberg-Thomsen, V.; Tan, T.; Zheng, D.; Strube, M.L.; Dragoš, A.; Shen, Q.; Zhang, R.; et al. Bacillus velezensis stimulates resident rhizosphere Pseudomonas stutzeri for plant health through metabolic interactions. ISME J. 2021, 30, 774–787. [Google Scholar] [CrossRef]
- Pandin, C.; Le Coq, D.; Deschamps, J.; Védie, R.; Rousseau, T.; Aymerich, S.; Briandet, R. Complete genome sequence of Bacillus velezensis QST713: A biocontrol agent that protects Agaricus bisporus crops against the green mould disease. J. Biotechnol. 2018, 278, 10–19. [Google Scholar] [CrossRef]
- Xu, S.; Xie, X.W.; Zhao, Y.R.; Shi, Y.X.; Chai, A.L.; Li, L.; Li, B.J. Whole-genome analysis of Bacillus velezensis ZF2, a biocontrol agent that protects cucumis sativus against corynespora leaf spot diseases. 3 Biotech 2020, 10, 186. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parnell, J.J.; Berka, R.; Young, H.A.; Sturino, J.M.; Kang, Y.; Barnhart, D.M.; DiLeo, M.V. From the Lab to the Farm: An Industrial Perspective of Plant Beneficial Microorganisms. Front. Plant Sci. 2016, 7, 1110. [Google Scholar] [CrossRef] [PubMed]
- Finkel, O.M.; Castrillo, G.; Herrera Paredes, S.; Salas González, I.; Dangl, J.L. Understanding and exploiting plant beneficial microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Cheeke, T.E.; Darby, H.; Rosenstiel, T.N.; Bever, J.D.; Cruzan, M.B. Effect of Bacillus thuringiensis (Bt) maize cultivation history on arbuscular mycorrhizal fungal colonization, spore abundance and diversity, and plant growth. Agric. Ecosyst. Environ. 2014, 195, 29–35. [Google Scholar] [CrossRef]
- Lee, B.Y.; Lim, H.R.; Choi, J.Y.; Ryu, K.H. Development of Molecular Detection of Three Species of Seed-Transmissible Viruses Useful for Plant Quarantine. Plant Pathol. J. 2004, 20, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Mangalam, A.K.; Dwivedi, S.; Naik, S. Primer premier: Program for design of degenerate primers from a protein sequence. Biotechniques 1998, 24, 318–319. [Google Scholar] [CrossRef] [Green Version]
- Borriss, R.; Chen, X.H.; Rueckert, C.; Blom, J.; Becker, A.; Baumgarth, B.; Fan, B.; Pukall, R.; Schumann, P.; Spröer, C.; et al. Relationship of Bacillus amyloliquefaciens clades associated with strains DSM 7T and FZB42T: A proposal for Bacillus amyloliquefaciens subsp. amyloliquefaciens subsp. nov. and Bacillus amyloliquefaciens subsp. plantarum subsp. nov. based on complete genome sequence comparisons. Int. J. Syst. Evol. Microbiol. 2011, 61, 1786–1801. [Google Scholar] [CrossRef]
- Tye, A.J.; Siu, F.K.; Leung, T.Y.; Lim, B.L. Molecular cloning and the biochemical characterization of two novel phytases from B. subtilis 168 and B. licheniformis. Appl. Microbiol. Biotechnol. 2002, 59, 190–197. [Google Scholar] [CrossRef]
- Liu, G.; Kong, Y.; Fan, Y.; Geng, C.; Peng, D.; Sun, M. Whole-genome sequencing of Bacillus velezensis LS69, a strain with a broad inhibitory spectrum against pathogenic bacteria. J. Biotechnol. 2017, 249, 20–24. [Google Scholar] [CrossRef]
- Li, J.Y.; Gao, T.T.; Wang, Q. Comparative and Functional Analyses of Two Sequenced Paenibacillus polymyxa Genomes Provides Insights into Their Potential Genes Related to Plant Growth-Promoting Features and Biocontrol Mechanisms. Front. Genet. 2020, 11, 564939. [Google Scholar] [CrossRef]
- Yuan, L.; Li, L.; Zheng, F.; Shi, Y.X.; Xie, X.W.; Chai, A.; Li, B.J. The complete genome sequence of Rahnella aquatilis ZF7 reveals potential beneficial properties and stress tolerance capabilities. Arch. Microbiol. 2019, 202, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yuan, L.F.; Shi, Y.X.; Xie, X.W.; Chai, A.L.; Wang, Q.; Li, B.J. Comparative genomic analysis of Pectobacterium carotovorum subsp. brasiliense SX309 provides novel insights into its genetic and phenotypic features. BMC Genom. 2019, 20, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Yuan, L.F.; Shi, Y.X.; Xie, X.W.; Chai, A.L.; Wang, Q.; Li, B.J. Comparative genomic analysis of Pseudomonas amygdali pv. lachrymans NM002: Insights into its potential virulence genes and putative invasion determinants. Genomics 2019, 111, 1493–1503. [Google Scholar] [CrossRef]
- Bedini, E.; Carabellese, A.; Barone, G.; Parrilli, M. First synthesis of the beta-D-rhamnosylated trisaccharide repeating unit of the O-antigen from Xanthomonas campestris pv. campestris 8004. J. Org. Chem. 2005, 70, 8064–8070. [Google Scholar] [CrossRef] [PubMed]
- Petrauskene, O.V.; Cao, Y.; Zoder, P.; Wong, L.Y.; Ba Lachandran, P.; Furtado, M.R.; Tebbs, R.S. Evaluation of applied biosystems MicroSEQ real-time PCR system for detection of Listeria spp. in food and environmental samples. J. AOAC Int. 2012, 95, 1074–1083. [Google Scholar] [CrossRef] [PubMed]
- Barman, A.; Kumar, P.; Kumar, V.; Doley, R.; Ray, S.K. Heat shock at 37 °C with plasmid ligated at 37 °C yields more number of Escherichia coli transformants than plasmid ligated at 16 °C a possible role of ligated plasmid conformation during heat shock. Curr. Sci. 2013, 104, 747–751. [Google Scholar]
- Vandersea, M.W.; Kibler, S.R.; Holland, W.C.; Tester, P.A.; Schultz, T.F.; Faust, M.A.; Holmes, M.J.; Chinain, M.; Wayne Litaker, R. Development of semi-quantitative pcr assays for the detection and enumeration of Gambierdiscus species (gonyaulacales, dinophyceae)1. J. Phycol. 2012, 48, 902–915. [Google Scholar] [CrossRef]
- Farrelly, V.F.; Rainey, F.A.; Stackebrandt, E. Effect of genome size and rrn gene copy number on PCR amplification of 16S rRNA genes from a mixture of bacterial species. Appl. Environ. Microbiol. 1995, 61, 2798–2801. [Google Scholar] [CrossRef] [Green Version]
- Glandorf, D.; Brand, I.; Bakker, P.; Schippers, B. Stability of rifampicin resistance as a marker for root colonization studies of Pseudomonas putida in the field. Plant Soil 1992, 147, 135–142. [Google Scholar] [CrossRef]
- Wang, B.B.; Yuan, J.; Zhang, J.; Shen, Z.Z.; Zhang, M.J. Effects of novel bioorganic fertilizer produced by Bacillus amyloliquefaciens W19 on antagonism of Fusarium wilt of banana. Biol. Fertil. Soils 2013, 49, 435–446. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, D.; Qi, G.; Mao, Z.; Hu, X.; Du, B.; Liu, K.; Ding, Y. Effects of Bacillus velezensis FKM10 for Promoting the Growth of Malus hupehensis Rehd. and Inhibiting Fusarium verticillioides. Front. Microbiol. 2020, 10, 2889. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Lee, Y.; Cheon, W.; Park, J.; Kwon, H.T.; Balaraju, K.; Kim, J.; Yoon, Y.J.; Jeon, Y. Characterization of Bacillus velezensis AK-0 as a biocontrol agent against apple bitter rot caused by Colletotrichum gloeosporioides. Sci. Rep. 2021, 11, 626. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.A.; Webster, J.A.; Straus, N. Rapid identification of bacteria on the basis of Polymerase Chain Reaction-amplified ribosomal DNA spacer polymorphism. Appl. Environ. Microbiol. 1993, 59, 945–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golpayegani, A.; Nodehi, R.N.; Rezaei, F.; Alimohammadi, M.; Douraghi, M. Real-time polymerase chain reaction assays for rapid detection and virulence evaluation of the environmental Pseudomonas aeruginosa isolates. Mol. Biol. Rep. 2019, 46, 4049–4061. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.G.; Wang, J.F.; Zhang, X.H.; Zhang, S.S.; Hu, X.F.; Chen, J.S. A gyrB-targeted PCR for rapid identification of Paenibacillus mucilaginosus. Appl. Microbiol. Biotechnol. 2010, 87, 739–747. [Google Scholar] [CrossRef]
- Irenge, L.M.; Durant, J.F.; Tomaso, H.; Pilo, P.; Olsen, J.S.; Ramisse, V.; Mahillon, J.; Gala, J.-L. Development and validation of a real-time quantitative PCR assay for rapid identification of Bacillus anthracis in environmental samples. Appl. Microbiol. Biotechnol. 2010, 88, 1179–1192. [Google Scholar] [CrossRef]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Borriss, R. Comparative analysis of the complete genome sequence of the plant growth-promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Shao, J.; Li, B.; Yan, X.; Shen, Q.; Zhang, R. Contribution of bacillomycin D in Bacillus amyloliquefaciens SQR9 to antifungal activity and biofilm formation. Appl. Environ. Microbiol. 2013, 79, 808–815. [Google Scholar] [CrossRef] [Green Version]
- Hesse, C.; Schulz, F.; Bull, C.T.; Shaffer, B.T.; Yan, Q.; Shapiro, N.; Hassan, K.A.; Varghese, N.; Elbourne, L.D.H.; Paulsen, I.T.; et al. Genome-based evolutionary history of Pseudomonas spp. Environ. Microbiol. 2018, 20, 2142–2159. [Google Scholar] [CrossRef]
- Xun, W.; Li, W.; Xiong, W.; Ren, Y.; Liu, Y.; Miao, Y.; Xu, Z.; Zhang, N.; Shen, Q.; Zhang, R. Diversity-triggered deterministic bacterial assembly constrains community functions. Nat. Commun. 2019, 10, 3833. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Rio, T.G.D.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciancio, A.; Pieterse, C.M.; Mercado-Blanco, J. Editorial: Harnessing Useful Rhizosphere Microorganisms for Pathogen and Pest Biocontrol. Front. Microbiol. 2016, 7, 1620. [Google Scholar] [CrossRef]
- Weller, D.M. Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology 2007, 97, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.F.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere interactions: Root exudates, microbes, and microbial commumities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- Bashan, Y.; Puente, M.E.; Rodriguez-Mendoza, M.N.; Toledo, G.; Holguin, G.; Ferrera-Cerrato, R.; Pedrin, S. Survival of Azospirillum brasilense in the Bulk Soil and Rhizosphere of 23 Soil Types. Appl. Environ. Microbiol. 1995, 61, 1938–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadhave, K.R.; Devlin, P.F.; Ebertz, A.; Ross, A.; Gange, A.C. Soil Inoculation with Bacillus spp. Modifies Root Endophytic Bacterial Diversity, Evenness, and Community Composition in a Context-Specific Manner. Microb. Ecol. 2018, 76, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Mallon, C.A.; Le Roux, X.; van Doorn, G.S.; Dini-Andreote, F.; Poly, F.; Salles, J.F. The impact of failure: Unsuccessful bacterial invasions steer the soil microbial community away from the invader’s niche. ISME J. 2018, 12, 728–741. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Source of Strain or Accession |
|---|---|
| Bacillus velezensis ZF2 | CP032154.1 |
| Bacillus velezensis ZF128 | Isolated in lab |
| Bacillus velezensis ZF145 | Isolated in lab |
| Bacillus velezensis LS69 | CP015911.1 |
| Bacillus velezensis SQR9 | CP006890.1 |
| Bacillus velezensis FZB42T | NC_009725.2 |
| Bacillus amyloliquefaciens ZF75 | Isolated in lab |
| Bacillus amyloliquefaciens DSM 7T | NC_014551.1 |
| Bacillus subtilis ZF161 | Isolated in lab |
| Bacillus subtilis 168 | AL009126.3 |
| Bacillus safensis ZF438 | Isolated in lab |
| Paenibacillus polymyxa ZF129 | NZ_CP040829.1 |
| Paenibacillus peoriae ZF390 | Isolated in lab |
| Rahnella aceries ZF458 | Isolated in lab |
| Rahnella aquatilis ZF7 | NZ_CP032296.1 |
| Lysobacter enzymogenes CX03 | Isolated in lab |
| Pectobacterium Brasiliense | CP020350.1 |
| Pseudomonas amygdali pv. lachrymans | CP020351.1 |
| Xanthomonas campestris pv. campestris 8004 | NC_007086.1 |
| Strains | Identities % | Sequence ID | locus_tag |
|---|---|---|---|
| Bacillus velezensis ZF2 | 100.00 | CP032154.1 | D3N19_13500 |
| Bacillus velezensis WLYS23 | 100.00 | CP055160.1 | HUW56_06825 |
| Bacillus velezensis A2 | 100.00 | CP053717.1 | HNV93_RS18185 |
| Bacillus velezensis BIM B-1312D | 100.00 | CP050448.1 | HB674_13380 |
| Bacillus velezensis UB2017 | 100.00 | CP049741.1 | G7X27_RS13380 |
| Bacillus velezensis FJAT-52631 | 100.00 | CP045186.1 | F9286_13510 |
| Bacillus velezensis LC1 | 100.00 | CP044349.1 | F6467_13510 |
| Bacillus velezensis LG37 | 100.00 | CP023341.1 | CMV18_07105 |
| Bacillus velezensis ANSB01E | 100.00 | CP036518.1 | EYB46_16460 |
| Bacillus velezensis JT3-1 | 100.00 | CP032506.1 | D5H27_11180 |
| Bacillus velezensis LDO2 | 100.00 | CP029034.1 | DDE72_03255 |
| Bacillus velezensis DR-08 | 100.00 | CP028437.1 | DA376_13590 |
| Bacillus velezensis GQJK49 | 100.00 | CP021495.1 | BAGQ_RS19690 |
| Bacillus velezensis CBMB205 | 100.00 | CP011937.1 | AAV34_05955 |
| Bacillus velezensis JTYP2 | 100.00 | CP020375.1 | BAJT_13390 |
| Bacillus velezensis sx01604 | 100.00 | CP018200.1 | BLL65_RS19535 |
| Bacillus velezensis LS69 | 100.00 | CP015911.1 | A8142_RS19645 |
| Bacillus velezensis S3-1 | 100.00 | CP016373.1 | A5891_RS19675 |
| Bacillus velezensis CBMB205 | 100.00 | CP014838.1 | BCBMB205_RS13490 |
| Bacillus velezensis GUIA | 100.00 | CP094930.1 | MUB47_13415 |
| Bacillus velezensis MBI600 | 100.00 | CP094686.1 | MTR96_14500 |
| Bacillus velezensis AP3 | 100.00 | CP094294.1 | MRS49_RS03450 |
| Bacillus velezensis JJ47 | 100.00 | CP091288.1 | L3C11_13755 |
| Bacillus velezensis AB191 | 100.00 | CP089996.1 | LVY83_13415 |
| Bacillus velezensis B4-7 | 100.00 | CP080760.1 | K3A92_13320 |
| Bacillus velezensis BS-G1 | 100.00 | CP078149.1 | KV103_RS13320 |
| Bacillus velezensis J17-4 | 100.00 | CP060420.1 | H8P15_13320 |
| Bacillus velezensis KOF112 | 100.00 | AP024603.1 | KJS48_RS13235 |
| Bacillus velezensis WSM-1 | 100.00 | CP068989.1 | IPZ53_13320 |
| Bacillus velezensis GY65 | 100.00 | CP063157.1 | IMZ24_05720 |
| Bacillus velezensis BSC16a | 100.00 | CP062074.1 | IGB10_13320 |
| Bacillus velezensis ZF145 | 100.00 | CP061176.1 | IAQ68_13320 |
| Bacillus velezensis YB-130 | 100.00 | CP054562.1 | HUF97_RS13715 |
| Bacillus velezensis K01 | 100.00 | CP059344.1 | HU024_03380 |
| Bacillus velezensis ZeaDK315 | 95.94 | CP043809.1 | ETZ92_008920 |
| Bacillus velezensis K26 | 95.94 | CP023075.1 | CK238_14205 |
| Bacillus velezensis SGAir0473 | 95.94 | CP027868.1 | C1N92_05090 |
| Bacillus velezensis 10075 | 95.94 | CP025939.1 | C0W57_02140 |
| Bacillus velezensis ATR2 | 95.94 | CP018133.1 | BMJ37_RS20275 |
| Bacillus velezensis SRCM100072 | 95.94 | CP021888.1 | S100072_02830 |
| Bacillus velezensis C1 | 95.94 | CP064091.1 | IRJ21_RS06175 |
| Bacillus velezensis K203 | 95.94 | CP092185.1 | MF598_05880 |
| Bacillus velezensis LOH112 | 95.94 | CP092110.1 | LGL65_RS16875 |
| Bacillus velezensis Pilsner2-2 | 95.94 | OU015476.1 | NA |
| Bacillus velezensis Pilsner1-2 | 95.94 | OU015424.1 | NA |
| Bacillus velezensis SWUJ1 | 95.94 | CP077672.1 | KTT68_RS13370 |
| Bacillus velezensis AD-3 | 95.94 | AP024501.1 | BVAD3_RS21170 |
| Bacillus velezensis NST6 | 95.94 | CP063687.1 | BACVE_RS14395 |
| Bacillus velezensis JSRB 08 | 95.94 | CP059497.1 | H2N97_14425 |
| Bacillus velezensis ONU553 | 94.92 | CP043416.1 | FZE25_RS19965 |
| Bacillus velezensis At1 | 94.92 | CP041145.1 | D073_RS19585 |
| Bacillus velezensis AP183 | 94.92 | CP029296.1 | RZ52_RS20115 |
| Bacillus velezensis AGVL-005 | 94.92 | CP024922.1 | CU084_15155 |
| Bacillus velezensis S141 | 94.92 | AP018402.1 | BVS141_RS20160 |
| Bacillus velezensis G341 | 94.92 | CP011686.1 | ABH13_RS20940 |
| Bacillus velezensis KS04AU | 94.92 | CP092750.1 | MKF36_13400 |
| Bacillus velezensis GMEKP1 | 94.92 | CP076450.1 | KOM03_18910 |
| Bacillus velezensis BIOMA BV10 | 94.92 | CP059318.1 | HZT45_RS19200 |
| Bacillus velezensis SRCM102742 | 94.92 | CP028206.1 | C7M20_RS13645 |
| Bacillus velezensis SRCM102741 | 94.92 | CP028205.1 | C7M19_RS05115 |
| Bacillus velezensis KD1 | 94.53 | CP014990.2 | A2I97_19425 |
| Bacillus velezensis DMB05 | 94.92 | CP083715.1 | LAZ96_04145 |
| Bacillus velezensis S4 | 94.42 | CP050424.1 | BVELS4_RS20280 |
| Bacillus velezensis SRCM102752 | 94.42 | CP028961.1 | DBK22_RS19770 |
| Bacillus velezensis SRCM102747 | 94.42 | CP028211.1 | C7M25_RS20685 |
| Bacillus velezensis SRCM102746 | 94.42 | CP028210.1 | C7M24_RS19720 |
| Bacillus velezensis SRCM102744 | 94.42 | CP028208.1 | C7M22_RS20530 |
| Bacillus velezensis SRCM102743 | 94.42 | CP028207.1 | C7M21_RS19665 |
| Bacillus velezensis UFLA258 | 94.42 | CP039297.1 | E4T61_13605 |
| Bacillus velezensis 1B-23 | 94.42 | CP033967.1 | EG882_09440 |
| Bacillus velezensis NKG-1 | 94.42 | CP024203.1 | CS376_RS21905 |
| Bacillus velezensis GH1-13 | 94.42 | CP019040.1 | BVH55_14590 |
| Bacillus velezensis LABIM44 | 94.42 | CP079719.1 | KXY09_14350 |
| Bacillus velezensis Sam8H1 | 94.42 | CP069391.1 | JR311_14195 |
| Bacillus velezensis AS43.3 | 94.42 | CP003838.1 | B938_RS20490 |
| Bacillus velezensis KMU01 | 94.42 | CP063768.1 | IM712_RS10485 |
| Bacillus velezensis AK-0 | 94.42 | CP047119.1 | GE573_RS19575 |
| Bacillus velezensis Bac57 | 94.42 | CP033054.1 | D9777_RS16315 |
| Bacillus velezensis W1 | 94.42 | CP028375.1 | C9888_RS15230 |
| Bacillus velezensis BR-01 | 94.42 | CP090150.1 | LXH20_13635 |
| Bacillus velezensis DMB07 | 94.42 | CP083764.1 | LAZ98_14795 |
| Bacillus velezensis NZ4 | 94.42 | CP076119.1 | KM132_14025 |
| Bacillus velezensis JK19 | 94.42 | CP073781.1 | KEM64_04660 |
| Bacillus velezensis PEBA20 | 94.42 | CP046145.1 | GKO36_14775 |
| Soil/Source (Province) | Copies/g (Colonies/g) 1 Day | Copies/g (Colonies/g) 7 Day | Copies/g (Colonies/g) 14 Day | Copies/g (Colonies/g) 21 Day | Copies/g (Colonies/g) 28 Day | Copies/g (Colonies/g) 35 Day |
|---|---|---|---|---|---|---|
| Black soil/Hei Long Jiang | 1.48 × 107 | 9.68 × 106 | 5.92 × 106 | 3.50 × 106 | 1.35 × 106 | 8.64 × 105 |
| (3.59 × 108) | (7.90 × 107) | (2.12 × 107) | (1.73 × 107) | (3.79 × 107) | (1.33 × 107) | |
| Red soil/Hai Nan | 4.95 × 106 | 3.92 × 106 | 2.98 × 106 | 2.42 × 106 | 9.16 × 105 | 6.27 × 105 |
| (5.25 × 108) | (1.23 × 106) | (3.28 × 106) | (6.22 × 106) | (5.02 × 106) | (5.25 × 106) | |
| Yellow brown soil/He Bei | 8.08 × 107 | 3.96 × 107 | 2.31 × 107 | 1.93 × 107 | 1.22 × 107 | 9.92 × 106 |
| (4.99 × 108) | (4.99 × 106) | (1.59 × 105) | (2.71 × 106) | (1.05 × 106) | (7.08 × 105) | |
| Brown soil/Jiang Su | 4.12 × 107 | 2.20 × 106 | 1.65 × 106 | 8.96 × 105 | 5.77 × 105 | 4.16 × 105 |
| (2.79 × 108) | (2.01 × 106) | (3.54 × 105) | (6.75 × 105) | (5.75 × 105) | (5.54 × 105) | |
| Desert soil/Ning Xia | 3.40 × 108 | 2.76 × 108 | 1.63 × 108 | 1.12 × 108 | 1.01 × 108 | 9.36 × 107 |
| (3.64 × 108) | (2.93 × 108) | (2.44 × 108) | (2.77 × 108) | (1.59 × 108) | (1.12 × 108) |
| Soil | pH | Organic Matter (g/kg) | Total N (g/kg) | Ammonium N (mg/kg) | Nitrate N (mg/kg) | Total P (mg/g) | Available P (mg/kg) | Total K (mg/g) | Available K (mg/kg) |
|---|---|---|---|---|---|---|---|---|---|
| Brown soil | 8.28 | 29.65 | 1.97 | 6.35 | 26.81 | 1.49 | 151.42 | 16.30 | 450.5 |
| Black soil | 7.54 | 7.16 | 0.62 | 13.50 | 18.14 | 0.40 | 14.58 | 6.40 | 131.5 |
| Red soil | 8.07 | 18.00 | 1.44 | 21.81 | 31.40 | 1.04 | 75.15 | 17.10 | 368.5 |
| Desert soil | 8.40 | 3.27 | 0.42 | 24.02 | 43.75 | 0.51 | 2.93 | 14.75 | 90.5 |
| Yellow brown soil | 7.71 | 35.32 | 2.30 | 2.28 | 29.42 | 3.97 | 202.36 | 12.25 | 802.5 |
| pH Value | Copies/g (Colonies/g) 1 Day | Copies/g (Colonies/g) 7 Day | Copies/g (Colonies/g) 14 Day | Copies/g (Colonies/g) 21 Day | Copies/g (Colonies/g) 28 Day | Copies/g (Colonies/g) 35 Day |
|---|---|---|---|---|---|---|
| 4 | 1.23 × 106 | 7.22 × 105 | 4.06 × 105 | 2.97 × 105 | 1.86 × 105 | 1.30 × 105 |
| (1.35 × 107) | (1.41 × 106) | (1.31 × 106) | (3.15 × 105) | (4.27 × 105) | (1.34 × 105) | |
| 5 | 3.15 × 106 | 1.36 × 106 | 1.13 × 106 | 4.53 × 105 | 3.41 × 105 | 2.34 × 105 |
| (1.47 × 107) | (3.42 × 106) | (2.22 × 106) | (1.73 × 106) | (6.97 × 105) | (4.33 × 105) | |
| 6 | 3.70 × 107 | 2.82 × 106 | 2.02 × 106 | 1.12 × 106 | 6.68 × 105 | 6.75 × 105 |
| (7.83 × 107) | (4.28 × 106) | (6.75 × 106) | (4.51 × 106) | (9.05 × 106) | (6.58 × 106) | |
| 7 | 1.88 × 108 | 4.42 × 107 | 2.63 × 107 | 2.52 × 107 | 1.51 × 107 | 8.54 × 106 |
| (7.81 × 107) | (1.57 × 107) | (5.17 × 107) | (1.24 × 107) | (1.58 × 107) | (1.02 × 107) | |
| 8 | 6.70 × 107 | 1.95 × 107 | 1.45 × 107 | 2.27 × 107 | 1.43 × 107 | 8.11 × 106 |
| (7.11 × 107) | (6.90 × 106) | (2.16 × 107) | (9.87 × 106) | (1.25 × 107) | (8.98 × 106) | |
| 9 | 8.59 × 106 | 2.81 × 106 | 2.34 × 106 | 2.22 × 106 | 2.27 × 106 | 1.55 × 106 |
| (6.43 × 107) | (6.51 × 106) | (7.16 × 106) | (4.21 × 106) | (3.98 × 106) | (4.56 × 106) | |
| 10 | 1.57 × 106 | 5.47 × 105 | 2.82 × 105 | 2.71 × 105 | 1.97 × 106 | 1.56 × 105 |
| (3.17 × 107) | (2.88 × 106) | (4.61 × 106) | (3.17 × 106) | (2.15 × 106) | (2.52 × 106) |
| Nutrient | Copies/g (Colonies/g) 1 Day | Copies/g (Colonies/g) 7 Day | Copies/g (Colonies/g) 14 day | Copies/g (Colonies/g) 21 Day | Copies/g (Colonies/g) 28 Day | Copies/g (Colonies/g) 35 Day |
|---|---|---|---|---|---|---|
| CK | 4.35 × 108 | 1.10 × 106 | 5.74 × 105 | 1.18 × 105 | 1.88 × 105 | 8.42 × 104 |
| (2.93 × 108) | (6.00 × 105) | (1.60 × 105) | (7.05 × 104) | (8.5 × 104) | (7.22 × 104) | |
| Fructose | 5.43 × 108 | 3.39 × 106 | 5.01 × 105 | 6.85 × 104 | 8.95 × 104 | 1.56 × 105 |
| (3.27 × 108) | (8.27 × 106) | (1.12 × 105) | (2.12 × 104) | (2.25 × 104) | (1.28 × 105) | |
| Sucrose | 4.37 × 108 | 1.15 × 106 | 6.34 × 105 | 1.62 × 105 | 1.97 × 105 | 1.26 × 105 |
| (2.63 × 108) | (7.50 × 105) | (1.52 × 105) | (1.25 × 105) | (1.25 × 105) | (9.68 × 104) | |
| Maltose | 3.37 × 108 | 9.49 × 105 | 2.37 × 105 | 1.23 × 105 | 1.90 × 105 | 2.36 × 105 |
| (9.21 × 107) | (7.50 × 105) | (1.05 × 105) | (1.55 × 105) | (1.55 × 105) | (1.76 × 105) | |
| Molasses | 6.56 × 108 | 1.03 × 106 | 1.73 × 105 | 1.16 × 105 | 1.10 × 105 | 1.55 × 105 |
| (2.03 × 108) | (9.00 × 105) | (8.15 × 104) | (6.24 × 104) | (6.16 × 104) | (6.32 × 104) | |
| Dextrin | 7.95 × 108 | 2.75 × 106 | 1.53 × 106 | 1.42 × 105 | 1.21 × 105 | 1.30 × 105 |
| (4.24 × 108) | (3.55 × 106) | (1.24 × 106) | (8.48 × 104) | (8.28 × 104) | (7.60 × 104) | |
| Corn flour | 9.44 × 108 | 2.59 × 106 | 6.21 × 105 | 6.61 × 105 | 3.665 × 105 | 5.17 × 105 |
| (8.65 × 108) | (4.10 × 106) | (3.90 × 105) | (4.2 × 105) | (3.73 × 105) | (3.18 × 105) | |
| Yeast extract | 5.61 × 108 | 2.98 × 106 | 1.35 × 106 | 6.79 × 105 | 5.11 × 105 | 5.22 × 105 |
| (2.41 × 108) | (5.07 × 106) | (1.04 × 106) | (5.07 × 105) | (2.53 × 105) | (3.35 × 105) | |
| Peanut meal | 7.69 × 108 | 3.52 × 106 | 1.14 × 106 | 9.32 × 105 | 6.51 × 105 | 7.77 × 105 |
| (7.91 × 108) | (8.43 × 106) | (1.30 × 106) | (6.62 × 105) | (4.06 × 105) | (5.62 × 105) | |
| Peptone | 8.44 × 108 | 1.99 × 106 | 1.04 × 106 | 1.14 × 106 | 1.51 × 106 | 1.02 × 106 |
| (6.97 × 108) | (4.83 × 106) | (1.37 × 106) | (7.93 × 105) | (1.07 × 106) | (1.12 × 106) | |
| Soybean flour | 6.51 × 108 | 6.63 × 106 | 1.11 × 106 | 1.02 × 106 | 5.65 × 105 | 6.24 × 105 |
| (6.5 × 108) | (1.13 × 107) | (1.74 × 106) | (7.62 × 105) | (7.67 × 105) | (6.74 × 105) | |
| Soluble starch | 5.27 × 108 | 1.64 × 106 | 4.59 × 105 | 6.03 × 105 | 1.76 × 105 | 1.59 × 105 |
| (3.51 × 108) | (1.25 × 106) | (3.93 × 105) | (1.35 × 105) | (9.52 × 104) | (9.25 × 104) | |
| MgCl2 | 5.91 × 108 | 7.19 × 105 | 3.77 × 105 | 1.45 × 105 | 1.78 × 105 | 1.32 × 105 |
| (3.99 × 108) | (5.50 × 105) | (1.60 × 105) | (7.06 × 104) | (8.55 × 104) | (7.20 × 104) | |
| CaCl2 | 5.59 × 108 | 1.23 × 106 | 3.37 × 105 | 1.40 × 105 | 1.07 × 105 | 1.06 × 105 |
| (3.78 × 108) | (3.87 × 106) | (6.00 × 105) | (5.05 × 104) | (6.25 × 104) | (5.62 × 104) | |
| FeSO4 | 3.35 × 108 | 8.39 × 105 | 3.80 × 105 | 1.23 × 105 | 1.25 × 105 | 1.19 × 105 |
| (7.75 × 107) | (1.05 × 106) | (3.77 × 105) | (1.16 × 105) | (6.05 × 104) | (5.04 × 104) | |
| KCl | 4.24 × 108 | 5.96 × 105 | 5.35 × 105 | 1.18 × 105 | 1.46 × 105 | 1.23 × 105 |
| (1.02 × 108) | (8.00 × 105) | (7.97 × 105) | (2.21 × 104) | (5.28 × 104) | (5.52 × 104) | |
| NaCl | 5.31 × 108 | 6.79 × 106 | 3.19 × 105 | 1.46 × 105 | 1.75 × 105 | 1.38 × 105 |
| (1.38 × 108) | (9.05 × 105) | (1.20 × 105) | (6.53 × 104) | (6.20 × 104) | (6.02 × 104) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, S.; Xie, X.; Shi, Y.; Chai, A.; Li, B.; Li, L. Development of a Real-Time Quantitative PCR Assay for the Specific Detection of Bacillus velezensis and Its Application in the Study of Colonization Ability. Microorganisms 2022, 10, 1216. https://doi.org/10.3390/microorganisms10061216
Xu S, Xie X, Shi Y, Chai A, Li B, Li L. Development of a Real-Time Quantitative PCR Assay for the Specific Detection of Bacillus velezensis and Its Application in the Study of Colonization Ability. Microorganisms. 2022; 10(6):1216. https://doi.org/10.3390/microorganisms10061216
Chicago/Turabian StyleXu, Shuai, Xuewen Xie, Yanxia Shi, Ali Chai, Baoju Li, and Lei Li. 2022. "Development of a Real-Time Quantitative PCR Assay for the Specific Detection of Bacillus velezensis and Its Application in the Study of Colonization Ability" Microorganisms 10, no. 6: 1216. https://doi.org/10.3390/microorganisms10061216
APA StyleXu, S., Xie, X., Shi, Y., Chai, A., Li, B., & Li, L. (2022). Development of a Real-Time Quantitative PCR Assay for the Specific Detection of Bacillus velezensis and Its Application in the Study of Colonization Ability. Microorganisms, 10(6), 1216. https://doi.org/10.3390/microorganisms10061216