
Estimation of Carbon Metabolism in Saccharomyces cerevisiae Acclimatized to Glycerol Assimilation with Quantitative PCR



Abstract
1. Introduction
2. Materials and Methods
2.1. Experimentally Original Strains and Culturing Media
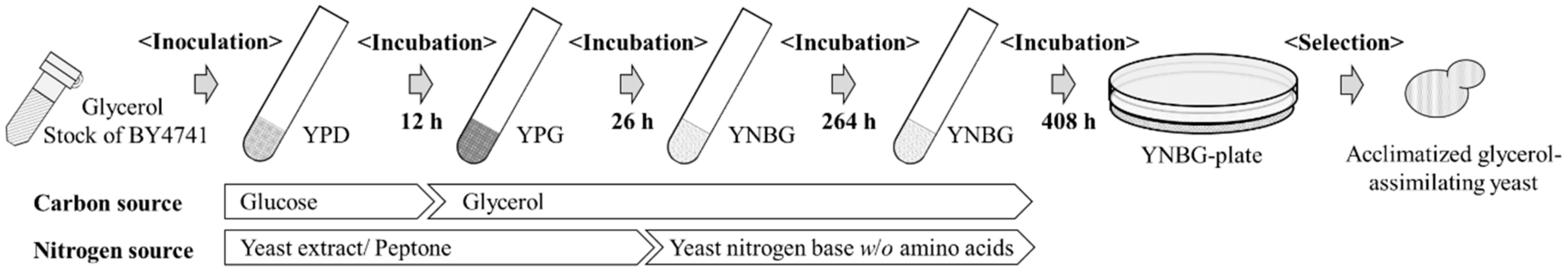
2.2. Selection of Acclimatized Strains in YNBG Medium
2.3. Primary Evaluation of Growth of Isolated Glycerol-Acclimatized Strains in YNBG Medium
2.4. Secondary Evaluation of Growth of No.7 in YNBG Medium
2.5. Evaluation of Relative Growth Rate and Doubling Time
2.6. Determination of Biomass Production
2.7. Quantification of Carbon Source and Ethanol by HPLC
2.8. Confirmation of Relevance of Glycerol-Acclimatized Strain No.7 to Wildtype
2.9. Transcript Analysis
2.10. Yeast Transformation
3. Results and Discussion
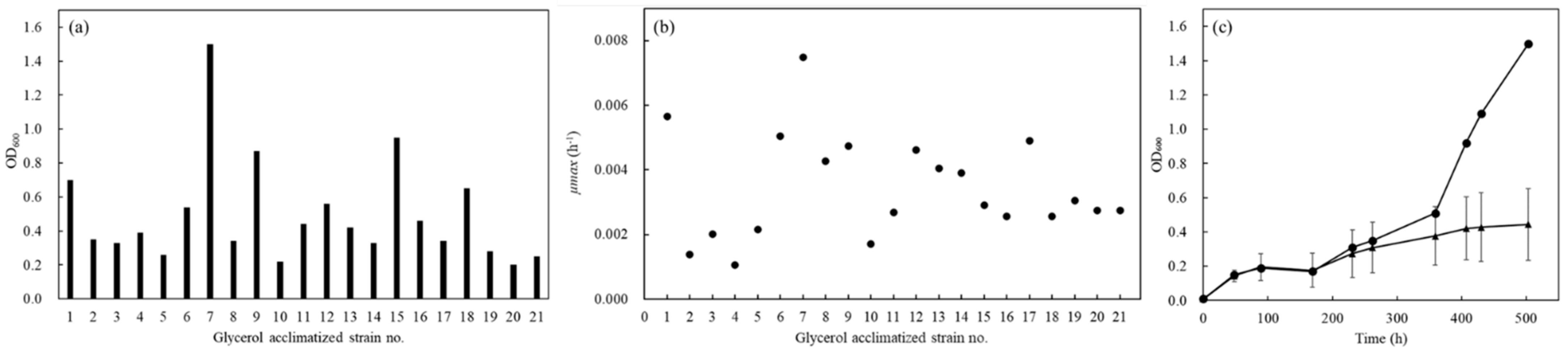
3.1. Overview of Picking up BY4741 Strains Acclimatized to Glycerol Assimilation
3.2. Growth Profiles of Glycerol-Acclimatized Strains
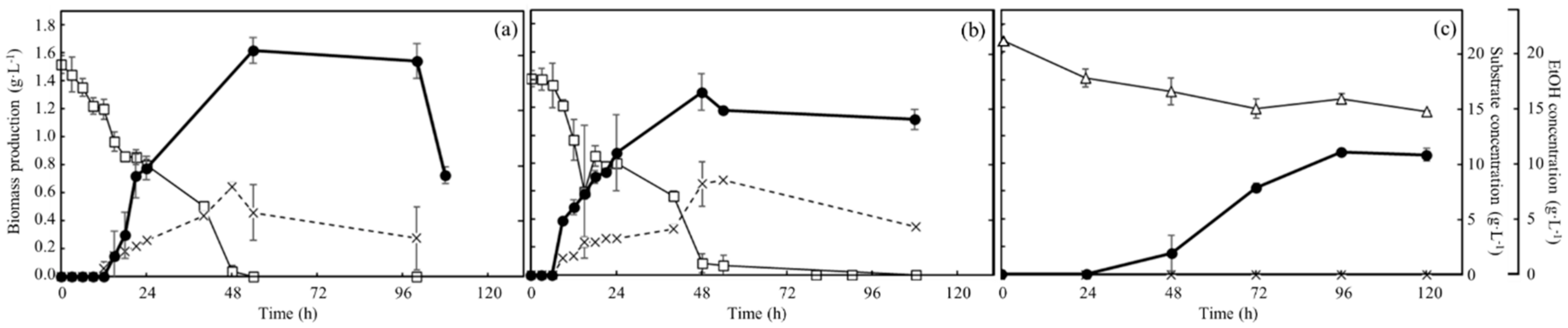
3.3. Time-Course Profiles of Biomass Production, Carbon Source Consumption, and Ethanol Production of Wildtype and No.7 in Each Medium under Aerobic Condition
3.4. Time-Course Profiles of Carbon Source Consumption and Ethanol Production of Wildtype and No.7 in Each Medium under Anaerobic Condition
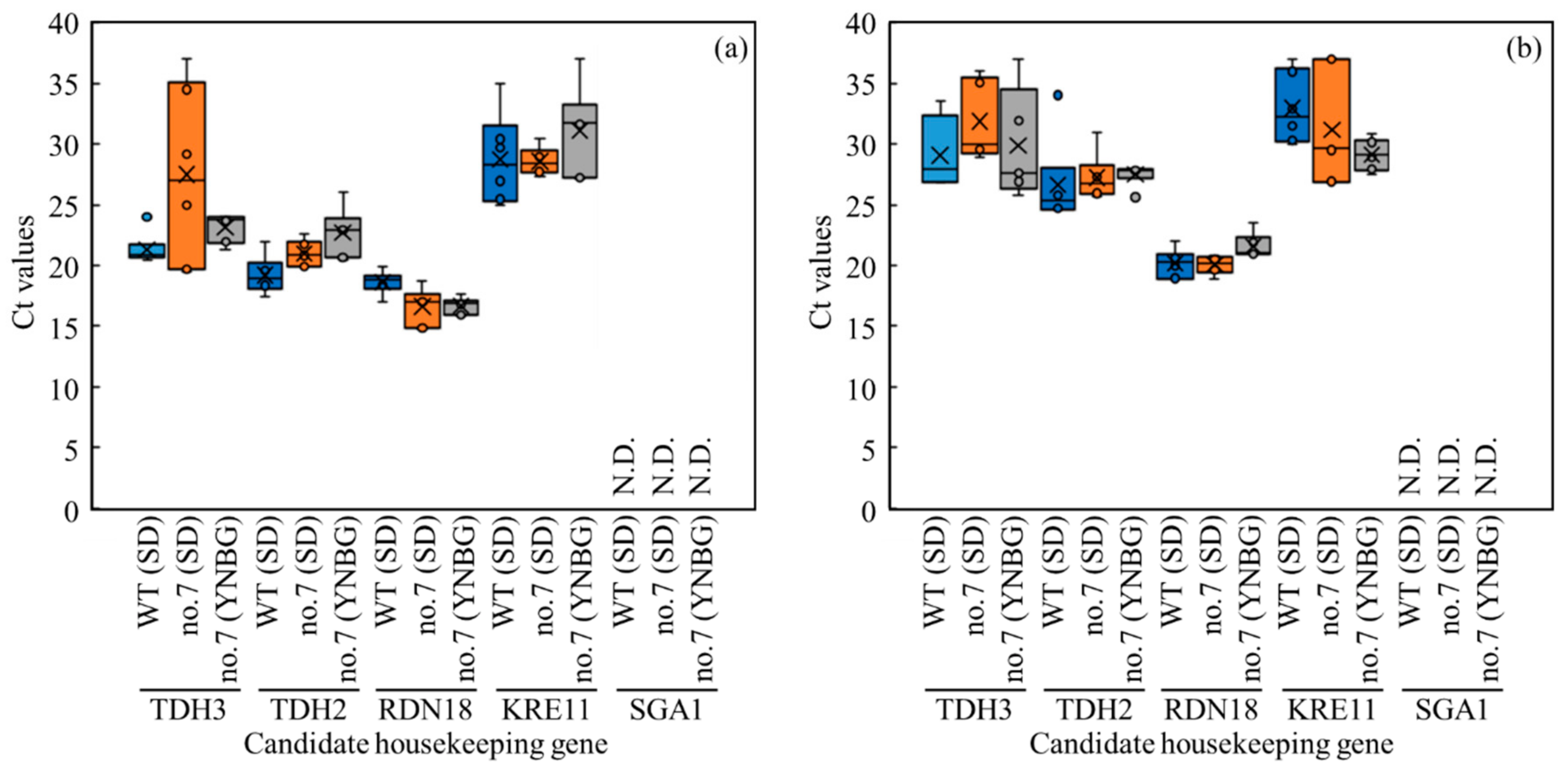
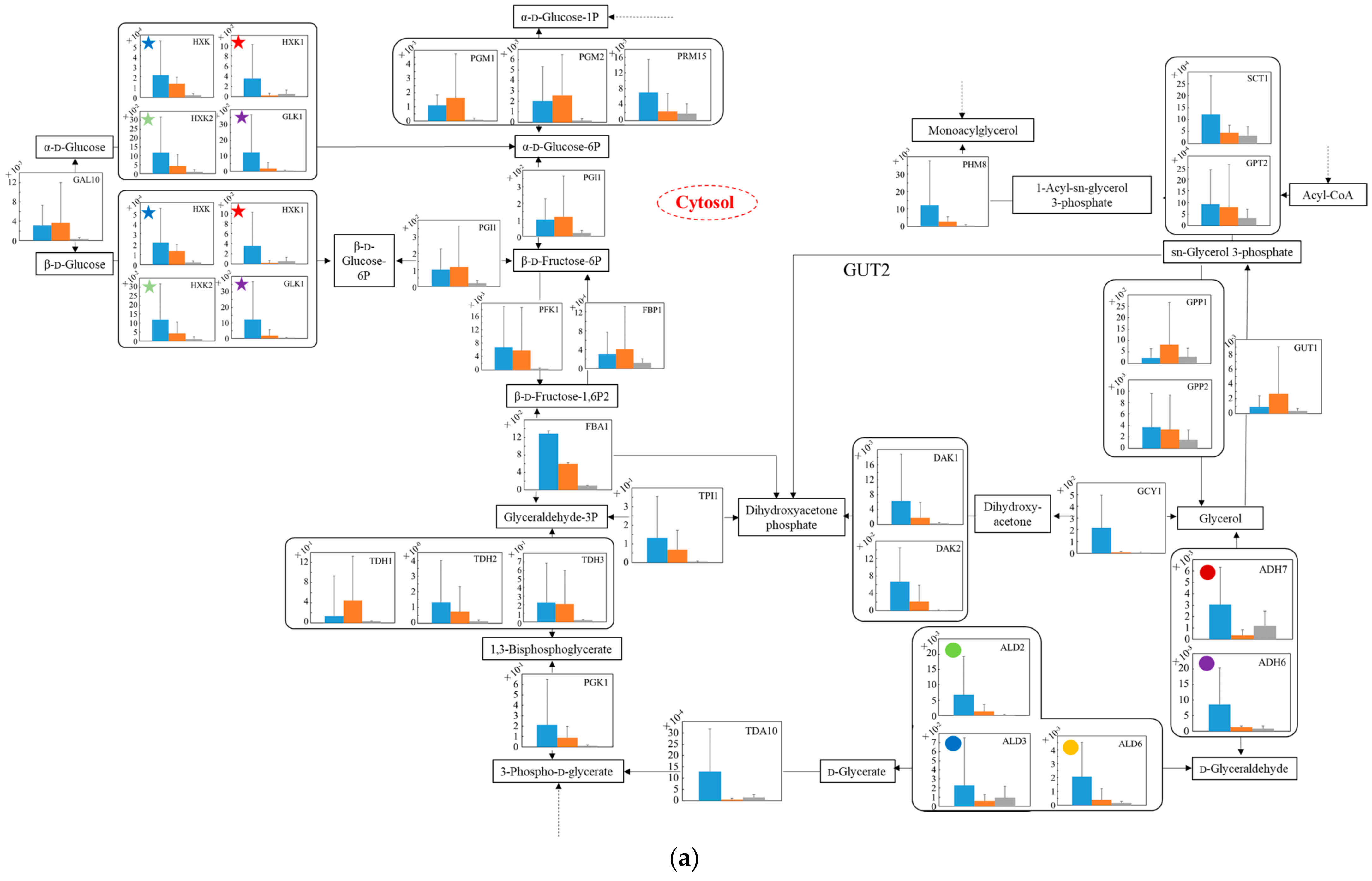
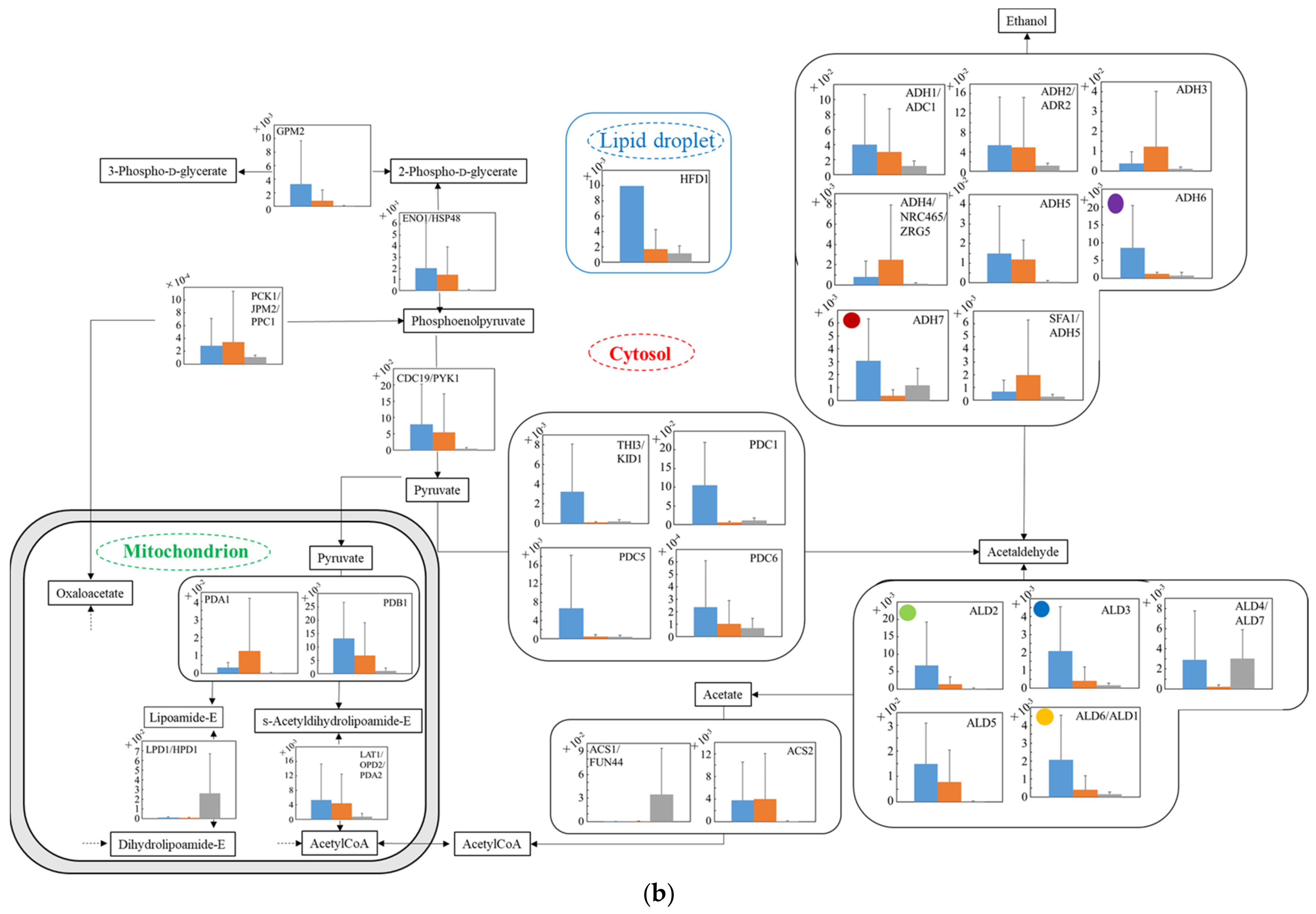
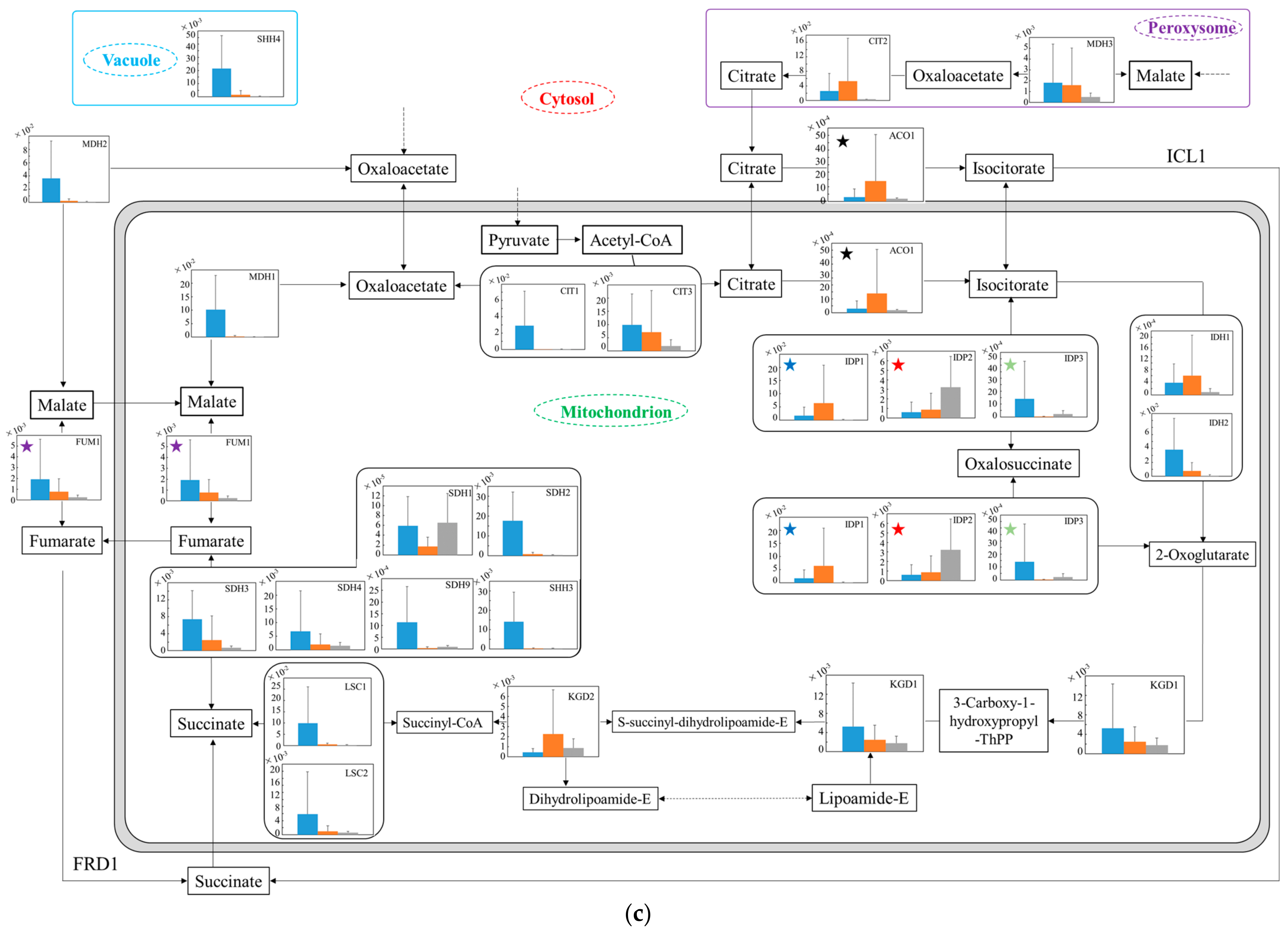
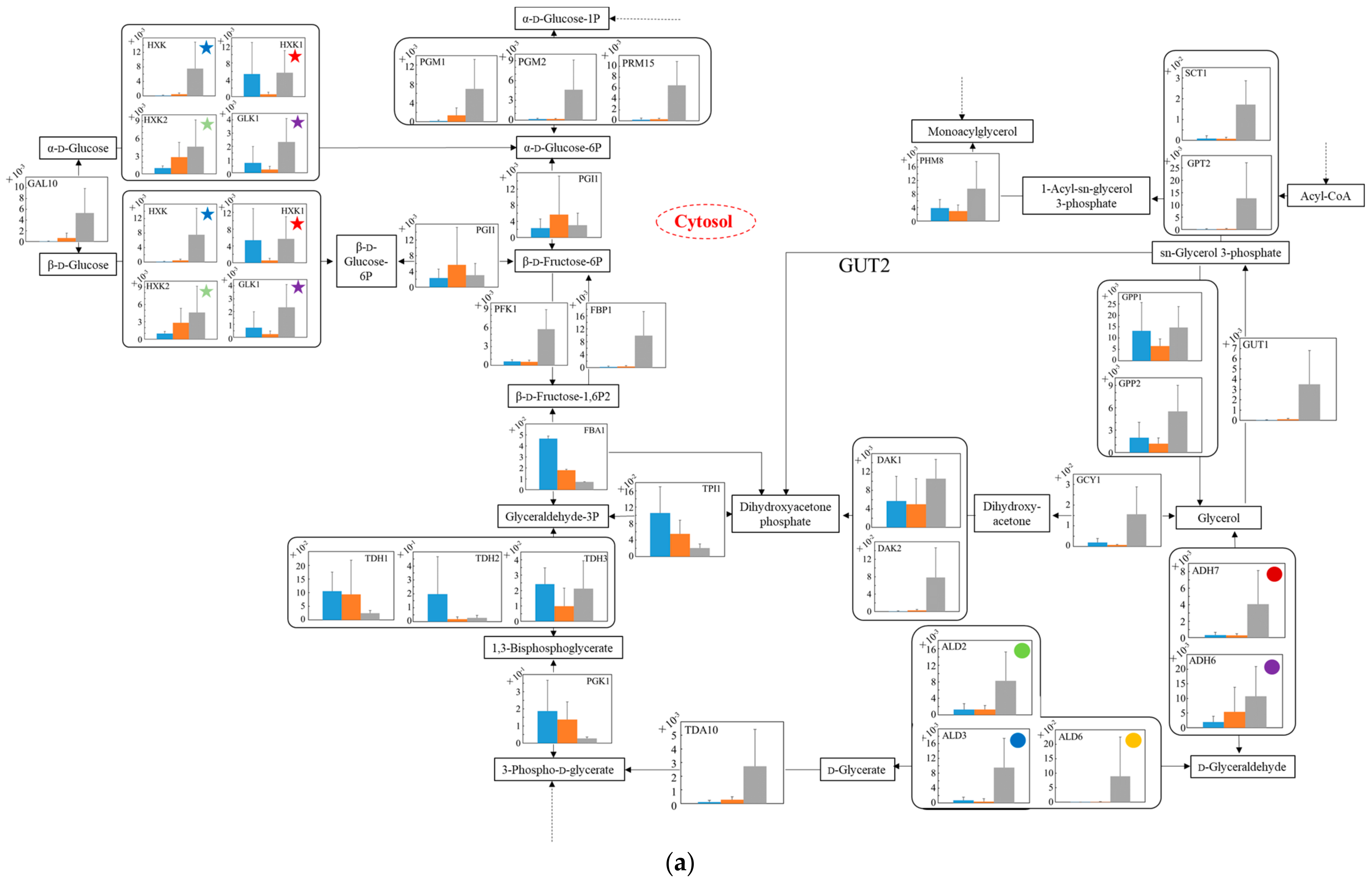
3.5. Distribution of Transcription Levels of Candidate Genes as Housekeeping Genes in Wildtype and No.7
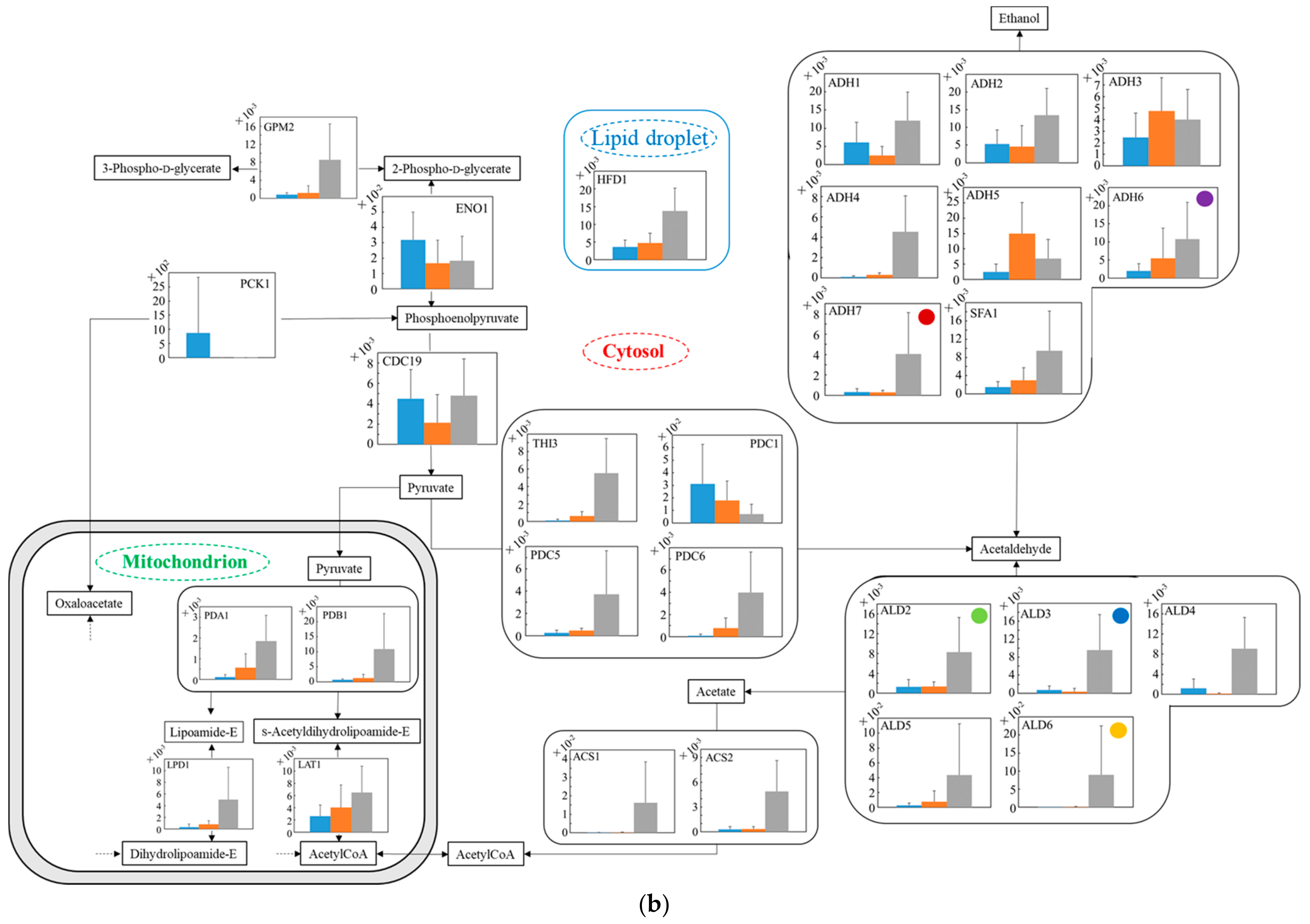
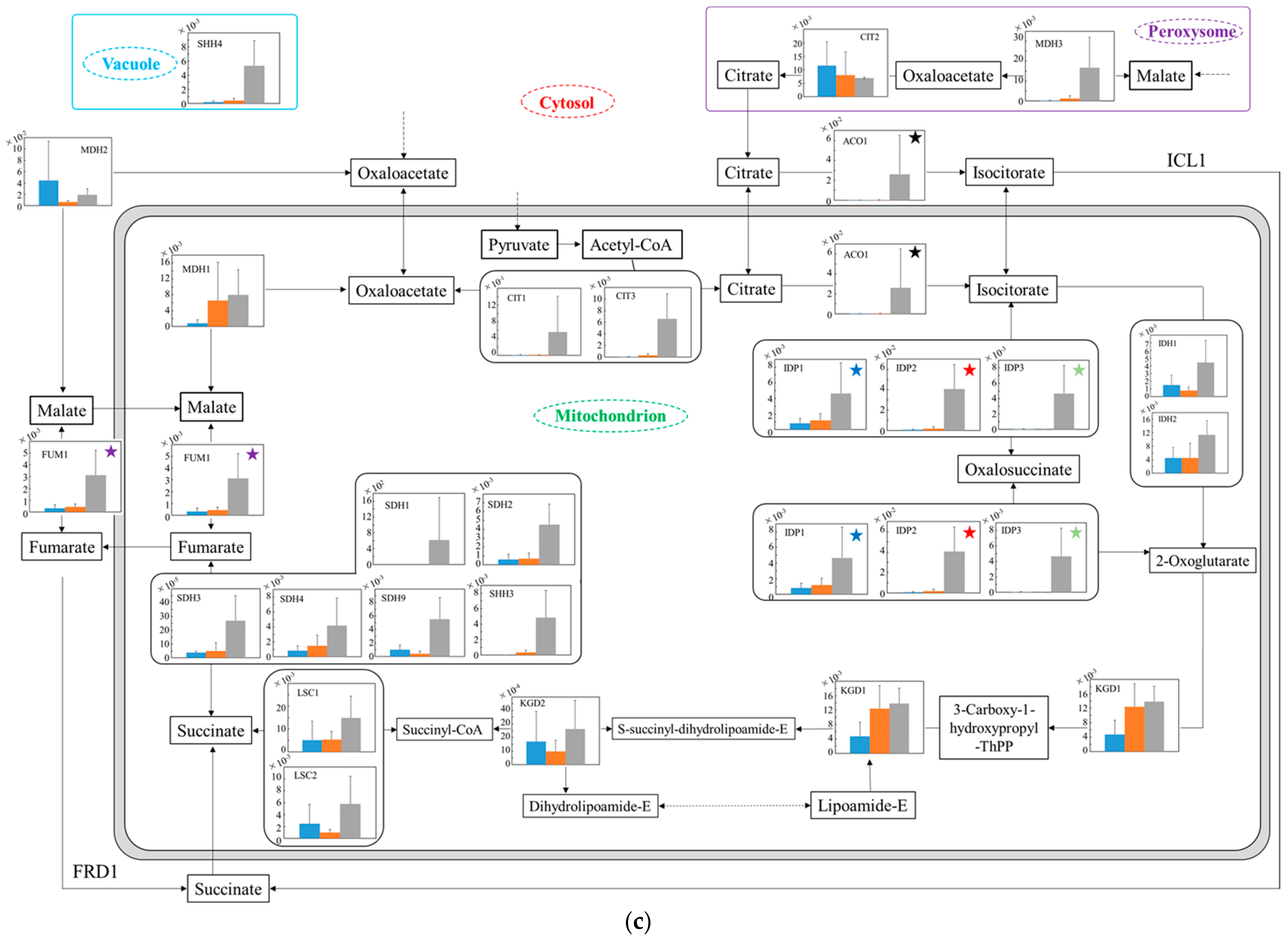
3.6. Relative Quantification of mRNA of Wildtype and No.7 in SD and in SD/YNBG under Aerobic Condition
3.7. Relative Quantification of mRNA of Wildtype and No.7 in SD and in SD/YNBG under Anaerobic Condition
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Clomburg, J.M.; Gonzalez, R. Anaerobic fermentation of glycerol: A platform for renewable fuels and chemicals. Trends Biotechnol. 2013, 31, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Hanna, M.A.; Sun, R. Value-added uses for crude glycerol—A byproduct of biodiesel production. Biotechnol. Biofuels 2012, 5, 13. [Google Scholar] [CrossRef]
- Johnson, E.; Sarchami, T.; Kießlich, S.; Munch, G.; Rehmann, L. Consolidating biofuel platforms through the fermentative bioconversion of crude glycerol to butanol. World J. Microbiol. Biotechnol. 2016, 32, 103. [Google Scholar] [CrossRef]
- Raza, M.; Inayat, A.; Abu-Jdayil, B. Crude glycerol as a potential feedstock for future energy via thermochemical conversion processes: A review. Sustainability 2021, 13, 12813. [Google Scholar] [CrossRef]
- Nandy, S.K.; Srivastava, R.K. A review on sustainable yeast biotechnological processes and applications. Microbiol. Res. 2018, 207, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Parapouli, M.; Vasileiadis, A.; Afendra, A.S.; Hatziloukas, E. Saccharomyces cerevisiae and its industrial applications. AIMS Microbiol. 2020, 6, 1–31. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific opinion on the safety and efficacy of Saccharomyces cerevisiae (NBRC 0203), Lactobacillus plantarum (NBRC 3070) and Lactobacillus casei (NBRC 3425) as a silage additive for all species. EFSA J. 2013, 11, 3362. [Google Scholar]
- Soares, E.V.; Hebbelinck, K.; Soares, H.M. Toxic effects caused by heavy metals in the yeast Saccharomyces cerevisiae: A comparative study. Can. J. Microbiol. 2003, 49, 336–343. [Google Scholar] [CrossRef]
- Yasokawa, D.; Murata, S.; Iwahashi, Y.; Kitagawa, E.; Nakagawa, R.; Hashido, T.; Iwahashi, H. Toxicity of methanol and formaldehyde towards Saccharomyces cerevisiae as assessed by DNA microarray analysis. Appl. Biochem. Biotechnol. 2010, 160, 1685–1698. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D. Genetic engineering and synthetic genomics in yeast to understand life and boost biotechnology. Bioengineering 2020, 7, 137. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Estopier, A.; Lesage, J.; Gorret, N.; Guillouet, S.E. Kinetic analysis of a Saccharomyces cerevisiae strain adapted for improved growth on glycerol: Implications for the development of yeast bioprocesses on glycerol. Bioresour. Technol. 2010, 102, 1521–1527. [Google Scholar] [CrossRef]
- Klein, M.; Swinnen, S.; Thevelein, J.M.; Nevoigt, E. Glycerol metabolism and transport in yeast and fungi: Established knowledge and ambiguities. Environ. Microbiol. 2017, 19, 878–893. [Google Scholar] [CrossRef]
- Brachmann, C.B.; Davies, A.; Cost, G.J.; Caputo, E.; Li, J.; Hieter, P.; Boeke, J.D. Designer deletion strains derived from Saccharomyces cerevisiae S288C: A useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 1998, 14, 115–132. [Google Scholar] [CrossRef]
- Merico, A.; Ragni, E.; Galafassi, S.; Popolo, L.; Compagno, C. Generation of an evolved Saccharomyces cerevisiae strain with a high freeze tolerance and an improved ability to grow on glycerol. J. Ind. Microbiol. Biotechnol. 2011, 38, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, A.; Bae, J.; Kuroda, K.; Ueda, M. Construction of a novel selection system for endoglucanases exhibiting carbohydrate-binding modules optimized for biomass using yeast cell-surface engineering. AMB Express 2012, 2, 56. [Google Scholar] [CrossRef]
- Ishii, J.; Kondo, T.; Makino, H.; Ogura, A.; Matsuda, F.; Kondo, A. Three gene expression vector sets for concurrently expressing multiple genes in Saccharomyces cerevisiae. FEMS Yeast Res. 2014, 14, 399–411. [Google Scholar] [CrossRef]
- Swinnen, S.; Klein, M.; Carrillo, M.; McInnes, J.; Nguyen, H.T.T.; Nevoigt, E. Re-evaluation of glycerol utilization in Saccharomyces cerevisiae: Characterization of an isolate that grows on glycerol without supporting supplements. Biotechnol. Biofuels 2013, 6, 157. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.A.; Payne, R.W.; Yarro, D. Yeasts: Characteristics and Identification, 3rd ed.; Cambridge University: Cambridge, UK, 2000. [Google Scholar]
- Paciello, L.; de Alteriis, E.; Mazzoni, C.; Palermo, V.; Zueco, J.; Parascandola, P. Performance of the auxotrophic Saccharomyces cerevisiae BY4741 as host for the production of IL-1β in aerated fed-batch reactor: Role of ACA supplementation, strain viability, and maintenance energy. Microbial. Cell Factories 2009, 8, 70. [Google Scholar] [CrossRef]
- Xiberras, J.; Klein, M.; Nevoigt, E. Glycerol as a substrate for Saccharomyces cerevisiae based bioprocesses—Knowledge gaps regarding the central carbon catabolism of this ‘non-fermentable’ carbon source. Biotechnol. Adv. 2019, 37, 107378. [Google Scholar] [CrossRef] [PubMed]
- Bakker, B.M.; Overkamp, K.M.; van Maris, A.J.; Kötter, P.; Luttik, M.A.; van Dijken, J.P.; Pronk, J.T. Stoichiometry and compartmentation of NADH metabolism in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2001, 25, 15–37. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.O.; Kim, S.W.; Han, S.O. Engineering of glycerol utilization pathway for ethanol production by Saccharomyces cerevisiae. Bioresour. Technol. 2010, 101, 4157–4161. [Google Scholar] [CrossRef]
- Sasaki, K.; Tsuge, Y.; Kawaguchi, H.; Yasukawa, M.; Sasaki, D.; Sazuka, T.; Kamio, E.; Ogino, C.; Matsuyama, H.; Kondo, A. Sucrose purification and repeated ethanol production from sugars remaining in sweet sorghum juice subjected to a membrane separation process. Appl. Microbiol. Biotechnol. 2017, 101, 6007–6014. [Google Scholar] [CrossRef]
- Peña-Moreno, I.C.; Parente, D.C.; da Silva, J.M.; Mendonça, A.A.; Rojas, L.A.V.; de Morais-Junior, M.A.; de Barros-Pita, W. Nitrate boosts anaerobic ethanol production in an acetate-dependent manner in the yeast Dekkera bruxellensis. J. Ind. Microbiol. Biotechnol. 2019, 46, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Teste, M.A.; Duquenne, M.; François, J.M.; Parrou, J.L. Validation of reference genes for quantitative expression analysis by real-time RT-PCR in Saccharomyces cerevisiae. BMC Mol. Biol. 2009, 10, 99. [Google Scholar] [CrossRef]
- Dohn, R.; Xie, B.; Back, R.; Selewa, A.; Eckart, H.; Rao, R.P.; Basu, A. mDrop-seq: Massively parallel single-cell RNA-seq of Saccharomyces cerevisiae and Candida albicans. Vaccines 2021, 10, 30. [Google Scholar] [CrossRef]
- Vaudano, E.; Noti, O.; Costantini, A.; Garcia-Moruno, E. Identification of reference genes suitable for normalization of RT-qPCR expression data in Saccharomyces cerevisiae during alcoholic fermentation. Biotechnol. Lett. 2011, 33, 1593–1599. [Google Scholar] [CrossRef]
- Ho, P.W.; Swinnen, S.; Duitama, J.; Nevoigt, E. The sole introduction of two single-point mutations establishes glycerol utilization in Saccharomyces cerevisiae CEN.PK derivatives. Biotechnol. Biofuels 2017, 10, 10. [Google Scholar] [CrossRef]
- Islam, Z.; Kleina, M.; Aßkampa, M.R.; Ødumb, A.S.R.; Nevoigta, E. A modular metabolic engineering approach for the production of 1,2-propanediol from glycerol by Saccharomyces cerevisiae. Metab. Eng. 2017, 44, 223–235. [Google Scholar] [CrossRef][Green Version]
- Kawai, K.; Kanesaki, Y.; Yoshikawa, H.; Hirasawa, T. Identification of metabolic engineering targets for improving glycerol assimilation ability of Saccharomyces cerevisiae based on adaptive laboratory evolution and transcriptome analysis. J. Biosci. Bioeng. 2019, 128, 162–169. [Google Scholar] [CrossRef]
- Papapetridis, I.; Verhoeven, M.D.; Wiersma, S.J.; Goudriaan, M.; van Maris, A.J.A.; Pronk, J.T. Laboratory evolution for forced glucose-xylose co-consumption enables identification of mutations that improve mixed-sugar fermentation by xylose-fermenting Saccharomyces cerevisiae. FEMS Yeast Res. 2018, 18, foy056. [Google Scholar] [CrossRef]
- Thakur, S.S.; Geiger, T.; Chatterjee, B.; Bandilla, P.; Fröhlich, F.; Cox, J.; Mann, M. Deep and highly sensitive proteome coverage by LC-MS/MS without prefractionation. Mol. Cell Proteom. 2011, 10, M110.003699. [Google Scholar] [CrossRef]
- Petelenz-Kurdziel, E.; Kuehn, C.; Nordlander, B.; Klein, D.; Hong, K.-K.; Jacobson, T.; Dahl, P.; Schaber, J.; Nielsen, J.; Hohmann, S.; et al. Quantitative analysis of glycerol accumulation, glycolysis and growth under hyper osmotic stress. PLoS Comput. Biol. 2013, 9, e1003084. [Google Scholar] [CrossRef] [PubMed]
- Marta, V.; Kostyantyn, V. Metabolic engineering for high glycerol productionby the anaerobic cultures of Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2016, 101, 4403–4416. [Google Scholar]
- Daran-Lapujade, P.; Jansen, M.L.A.; Daran, J.M.; van Gulik, W.; de Winde, J.H.; Pronk, J.T. Role of transcriptional regulation in controlling fluxes in central carbon metabolism of Saccharomyces cerevisiae: A chemostat culture study. J. Biol. Chem. 2004, 279, 9125–9138. [Google Scholar] [CrossRef]
- Yu, M.; Juwono, N.K.P.; Foo, J.L.; Leong, S.S.; Chang, M.W. Metabolic engineering of Saccharomyces cerevisiae for the overproduction of short branched-chain fatty acids. Meta Eng. 2016, 34, 36–43. [Google Scholar] [CrossRef]
- Bakker, B.M.; Bro, C.; Kotter, P.; Luttik, M.A.H.; Dijken, J.P.; Pronk, J.T. The Mitochondrial alcohol Dehydrogenase Adh3p Is Involved in a Redox Shuttle in Saccharomyces cerevisiae. J. Bacteriol. 2000, 182, 4730–4737. [Google Scholar] [CrossRef]
- Hoang, P.T.N.; Ko, J.K.; Gong, G.; Um, Y.; Lee, S.M. Genomic and phenotypic characterization of a refactored xylose-utilizing Saccharomyces cerevisiae strain for lignocellulosic biofuel production. Biotechnol. Biofuel Bioprod. 2018, 11, 268. [Google Scholar] [CrossRef]
- Costenoble, R.; Valadi, H.; Gustafsson, L.; Niklasson, C.; Franzén, C.J. Microaerobic glycerol formation in Saccharomyces cerevisiae. Yeast 2000, 16, 1483–1495. [Google Scholar] [CrossRef]
- Norbeck, J.; Blomberg, A. Metabolic and regulatory changes associated with growth of Saccharomyces cerevisiae in 1.4 M NaCl: Evidence for osmotic induction of glycerol dissimilation via the dihydroxyacetone pathway. J. Biol. Chem. 1997, 272, 5544–5554. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wildtype in SD | No.7 in SD | No.7 in YNBG | ||||
|---|---|---|---|---|---|---|
| 0 h | 24 h | 0 h | 24 h | 0 h | 24 h | |
| Glucose (g·L−1) | 20.8 ± 1.1 | 2.6 ± 0.6 | 22.3 ± 1.2 | 0.7 ± 0.7 | ||
| Glycerol (g·L−1) | 21.7 ± 2.5 | 22.1 ± 0.2 | ||||
| Ethanol (g·L−1) | 0.0 ± 0.0 | 8.5 ± 0.2 | 0.0 ± 0.0 | 10.2 ± 0.5 | 0.0 ± 0.0 | 0.0 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakanishi, A.; Zhang, K.; Matsumoto, R.; Yamamoto, N. Estimation of Carbon Metabolism in Saccharomyces cerevisiae Acclimatized to Glycerol Assimilation with Quantitative PCR. Microorganisms 2022, 10, 1173. https://doi.org/10.3390/microorganisms10061173
Nakanishi A, Zhang K, Matsumoto R, Yamamoto N. Estimation of Carbon Metabolism in Saccharomyces cerevisiae Acclimatized to Glycerol Assimilation with Quantitative PCR. Microorganisms. 2022; 10(6):1173. https://doi.org/10.3390/microorganisms10061173
Chicago/Turabian StyleNakanishi, Akihito, Kuan Zhang, Riri Matsumoto, and Naotaka Yamamoto. 2022. "Estimation of Carbon Metabolism in Saccharomyces cerevisiae Acclimatized to Glycerol Assimilation with Quantitative PCR" Microorganisms 10, no. 6: 1173. https://doi.org/10.3390/microorganisms10061173
APA StyleNakanishi, A., Zhang, K., Matsumoto, R., & Yamamoto, N. (2022). Estimation of Carbon Metabolism in Saccharomyces cerevisiae Acclimatized to Glycerol Assimilation with Quantitative PCR. Microorganisms, 10(6), 1173. https://doi.org/10.3390/microorganisms10061173