
A Case Study Demonstrates That the Litter of the Rare Species Cinnamomum migao Composed of Different Tissues Can Affect the Chemical Properties and Microbial Community Diversity in Topsoil


Abstract
:1. Introduction
2. Materials and Methods
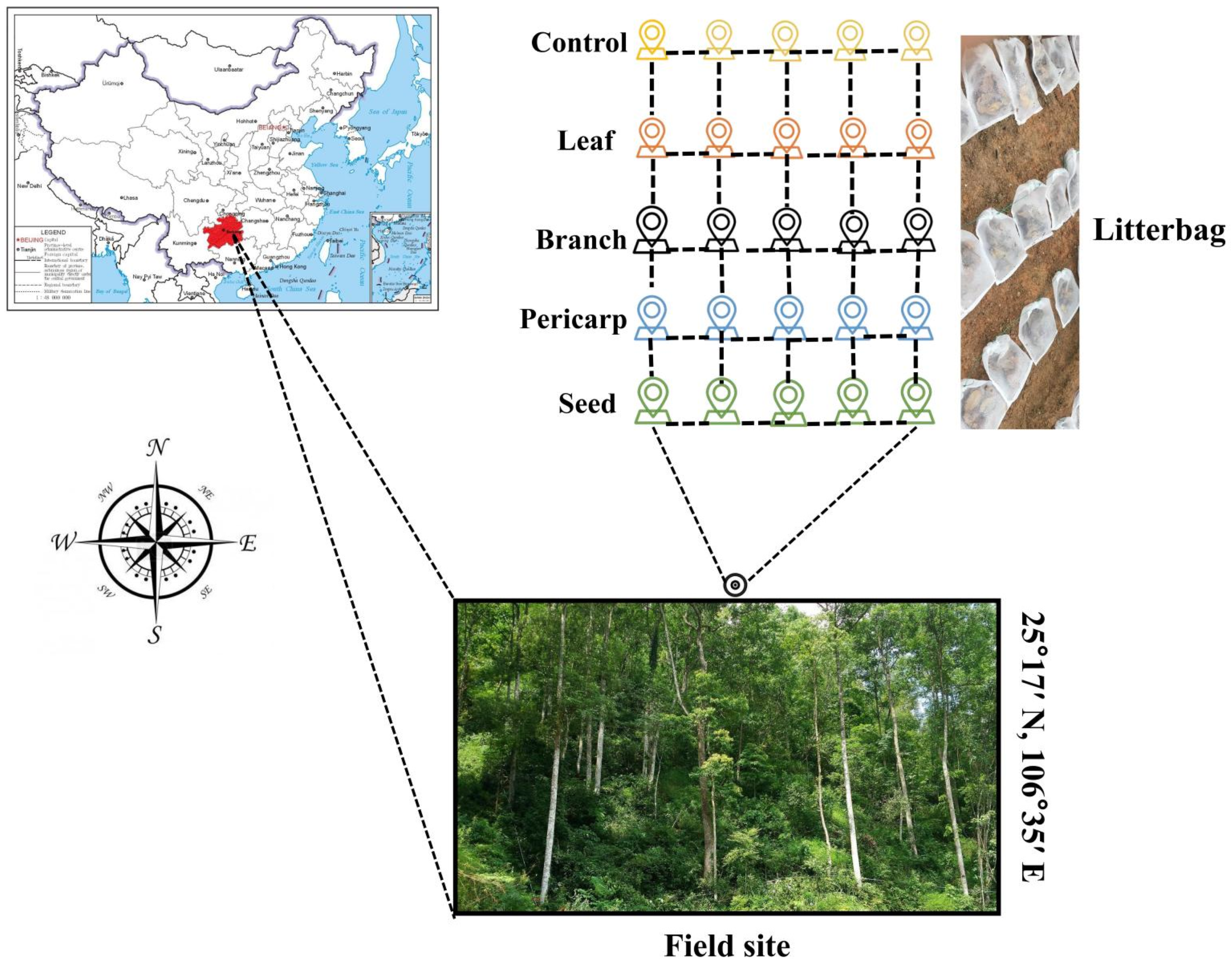
2.1. Study Site
2.2. Sample Preparation and Experimental Design
2.3. Sample Collection
2.4. Soil Nutrient and Enzyme Concentration Analysis
2.5. Soil DNA Extraction and PCR Amplification
2.6. Illumina MiSeq Sequencing
2.7. Statistical Analyses
2.7.1. Processing of Sequencing Data
2.7.2. Variance and Diversity Composition Analysis
2.7.3. Network Construction and Analysis
3. Results
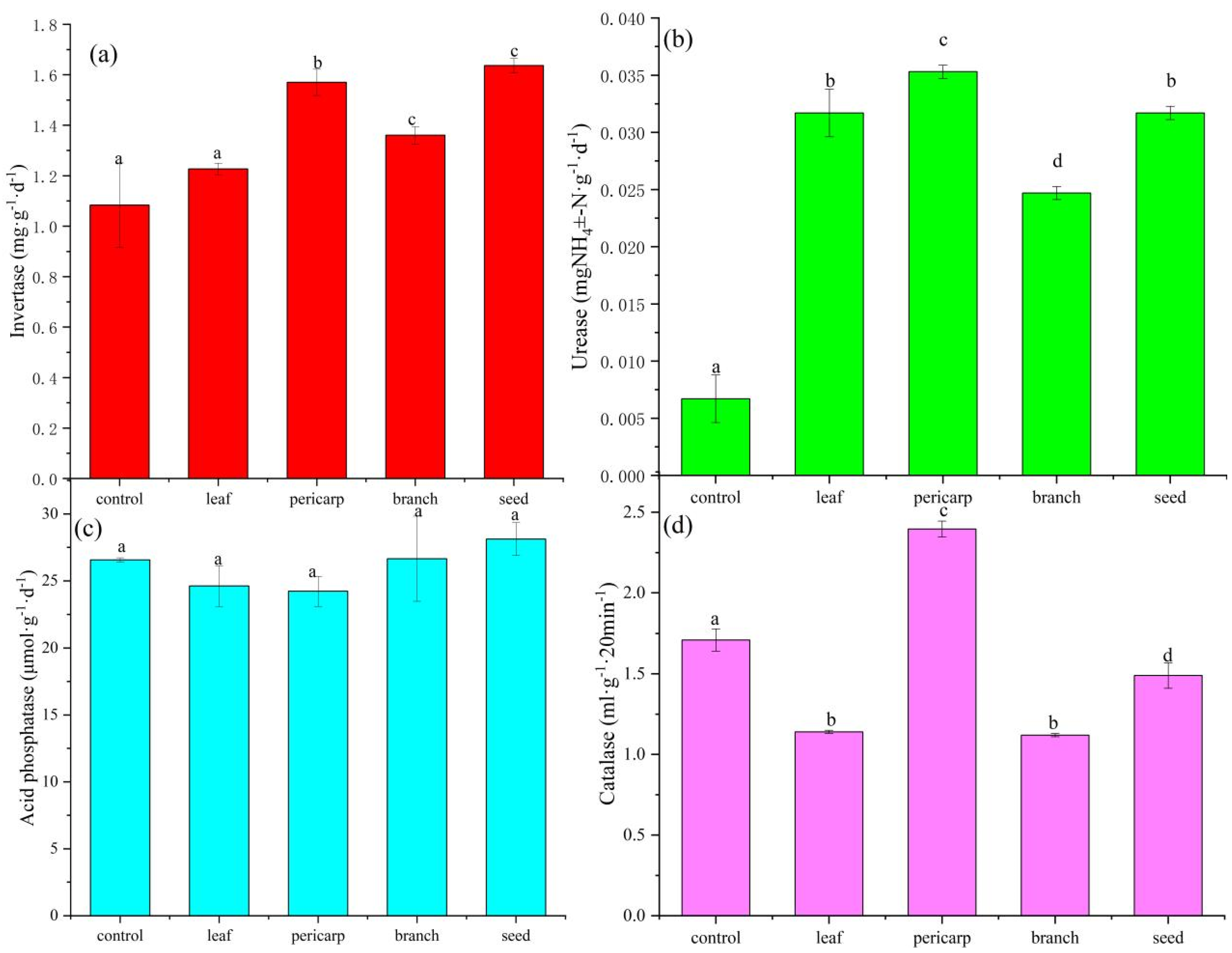
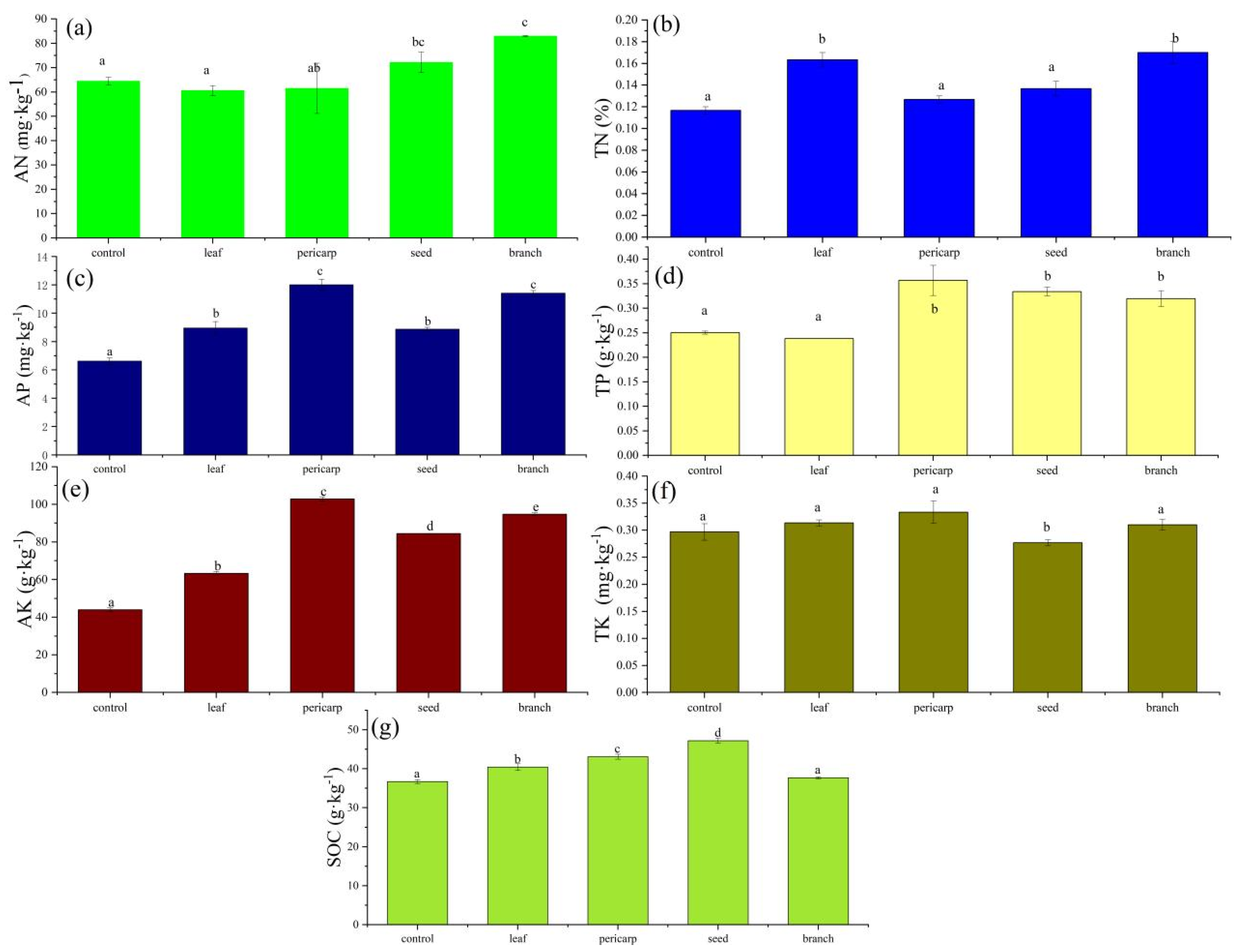
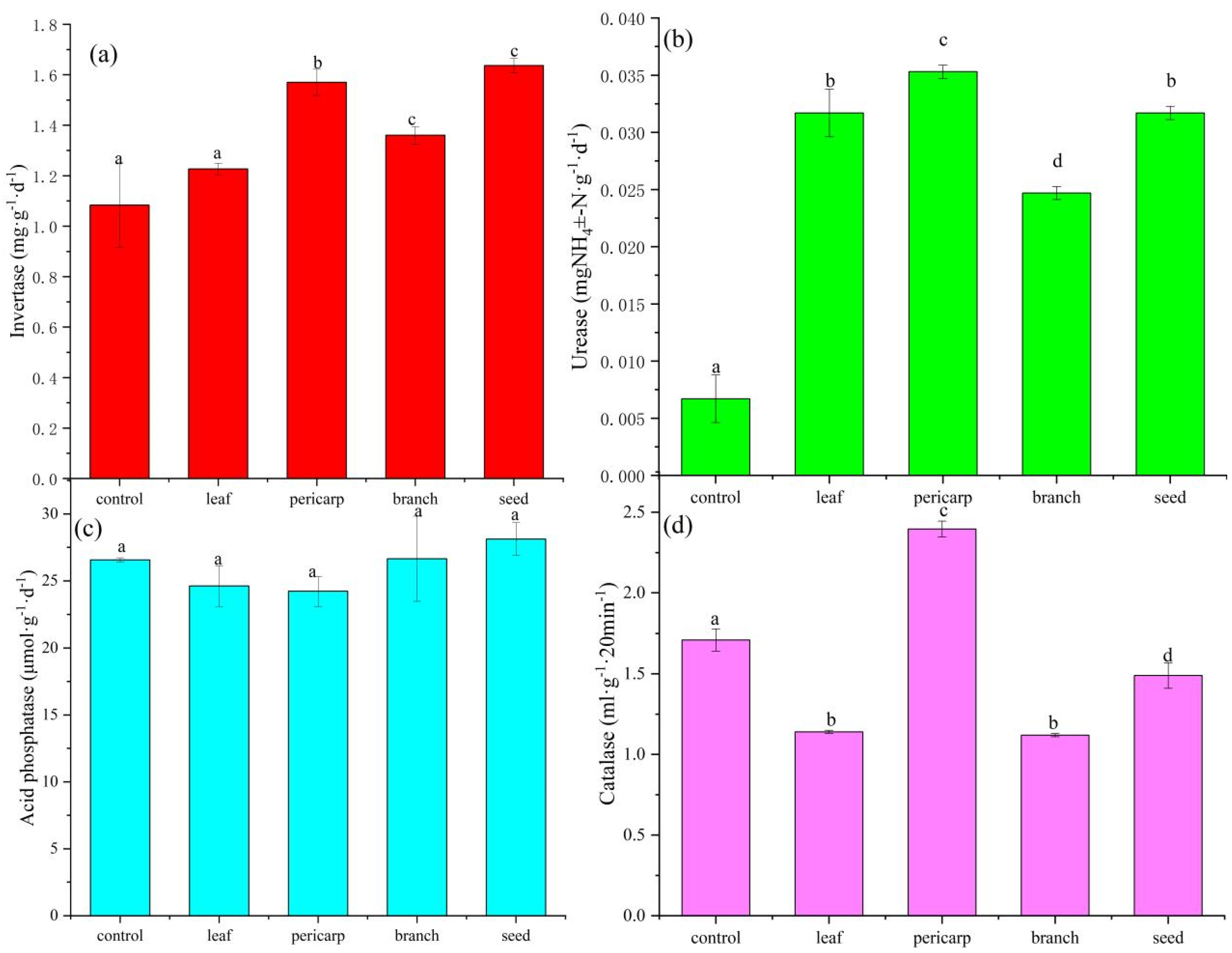
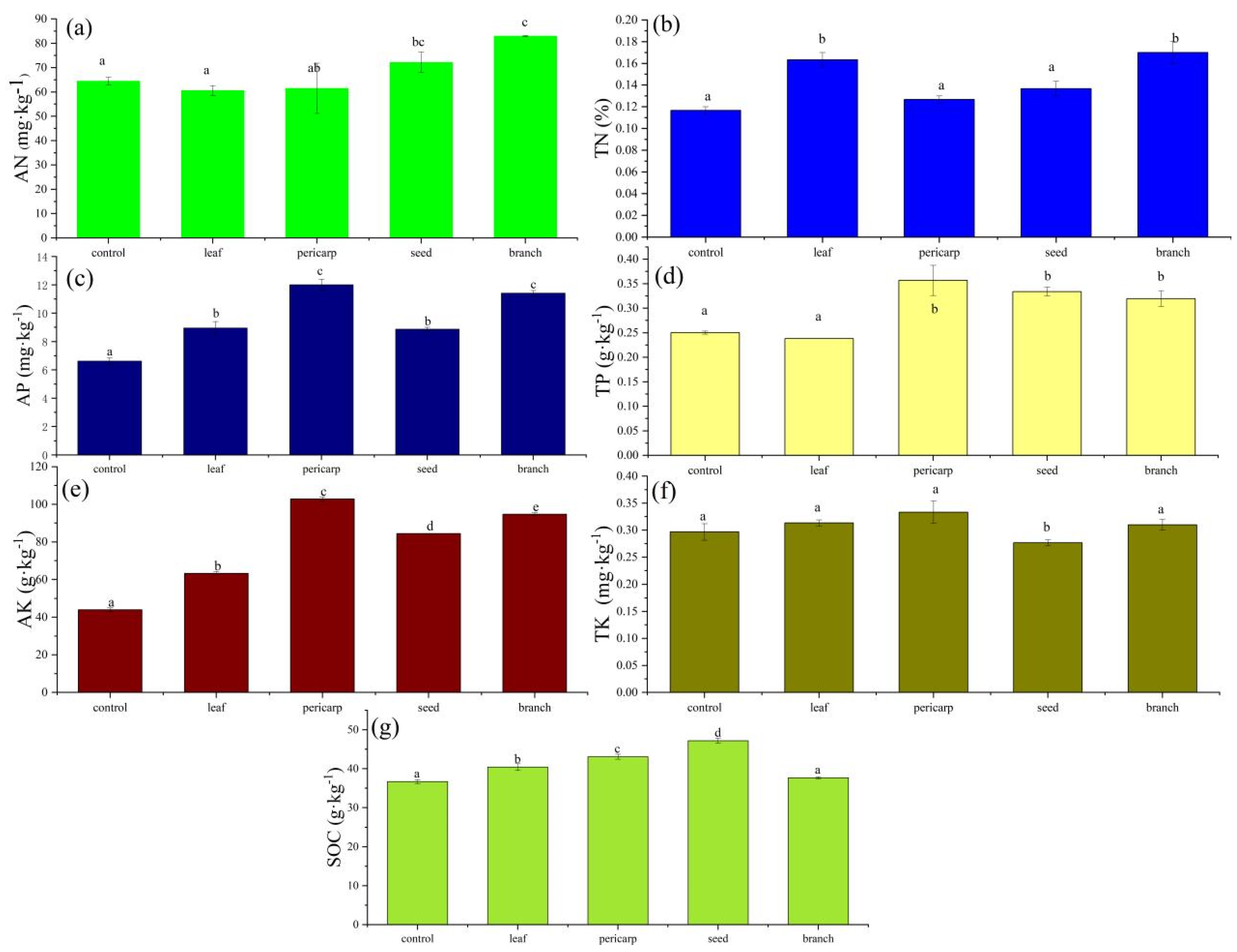
3.1. Soil Biochemical Properties
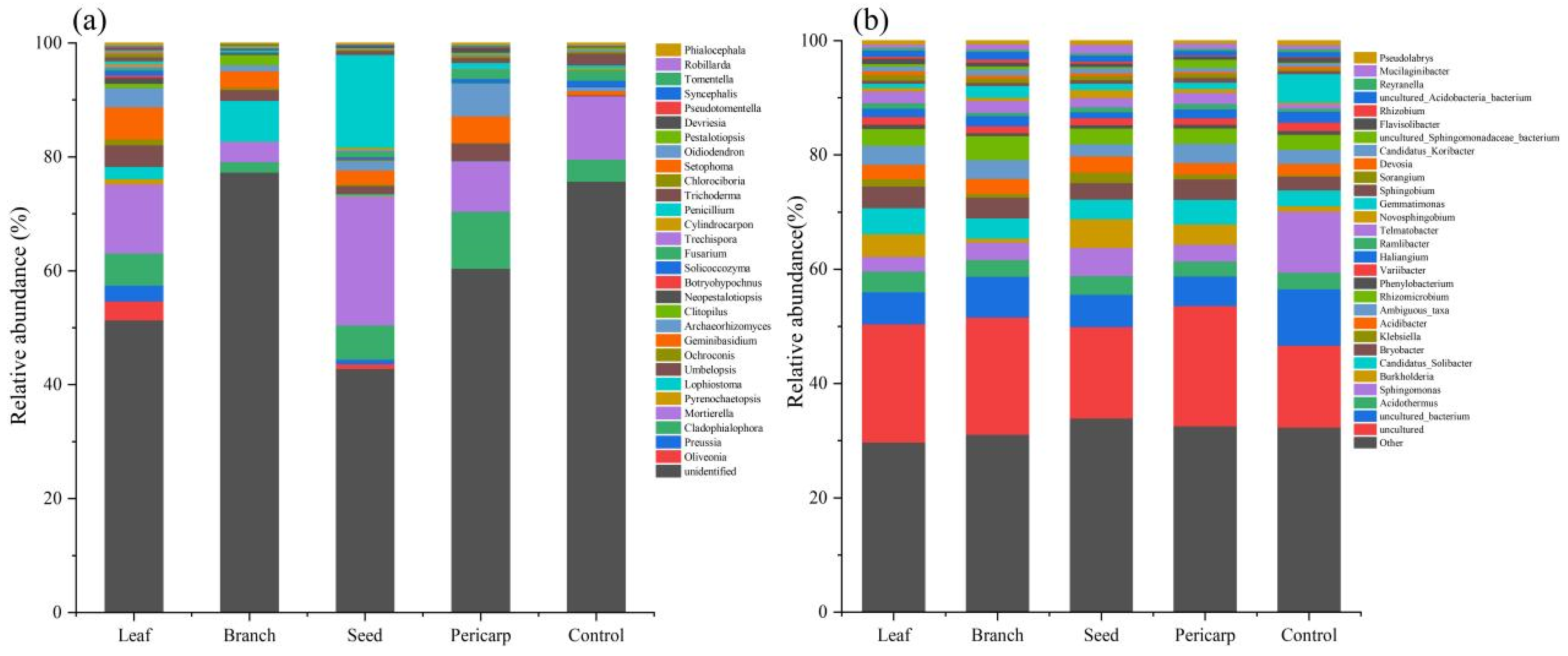
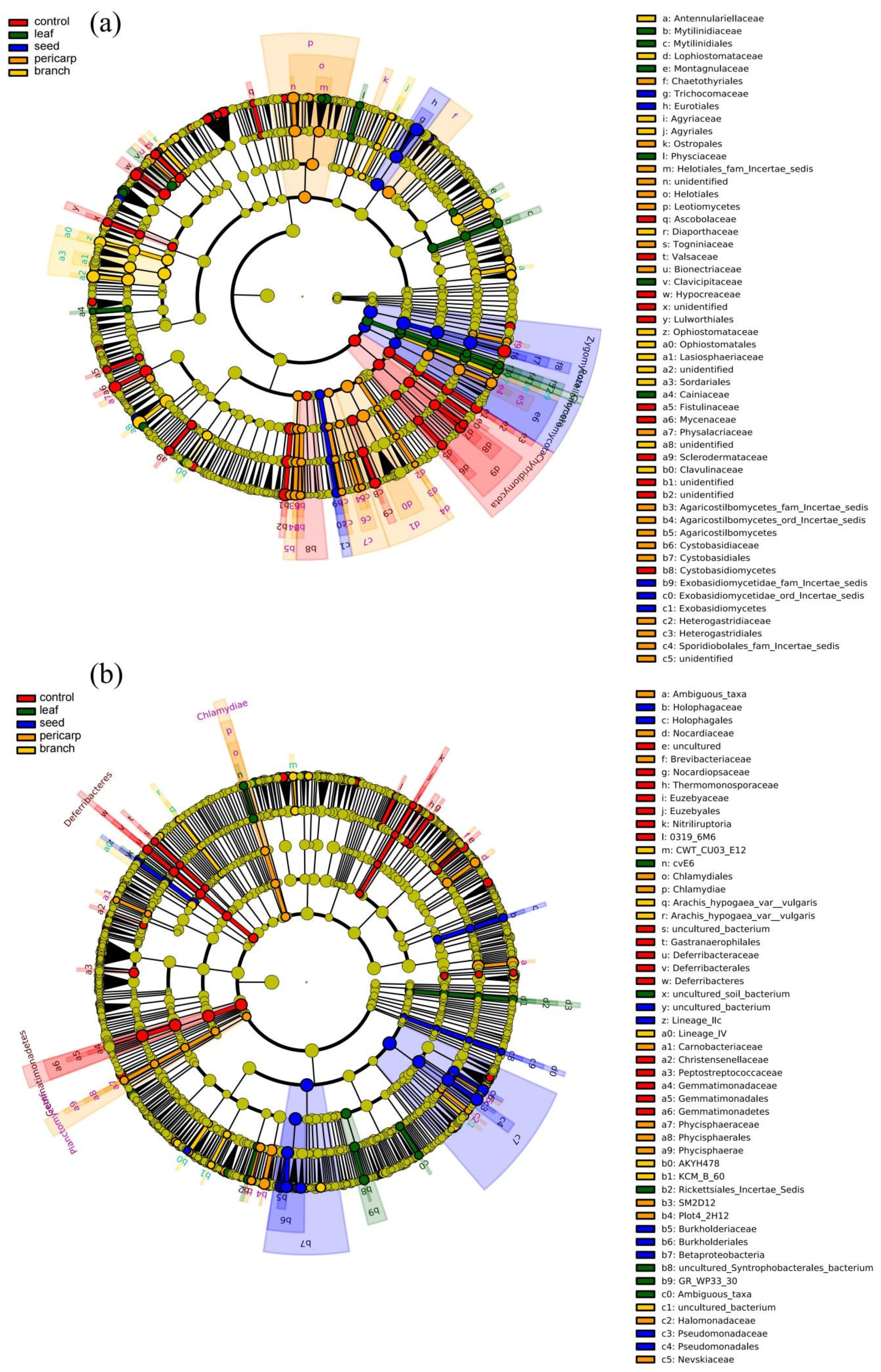
3.2. Alpha Diversity and Community Composition
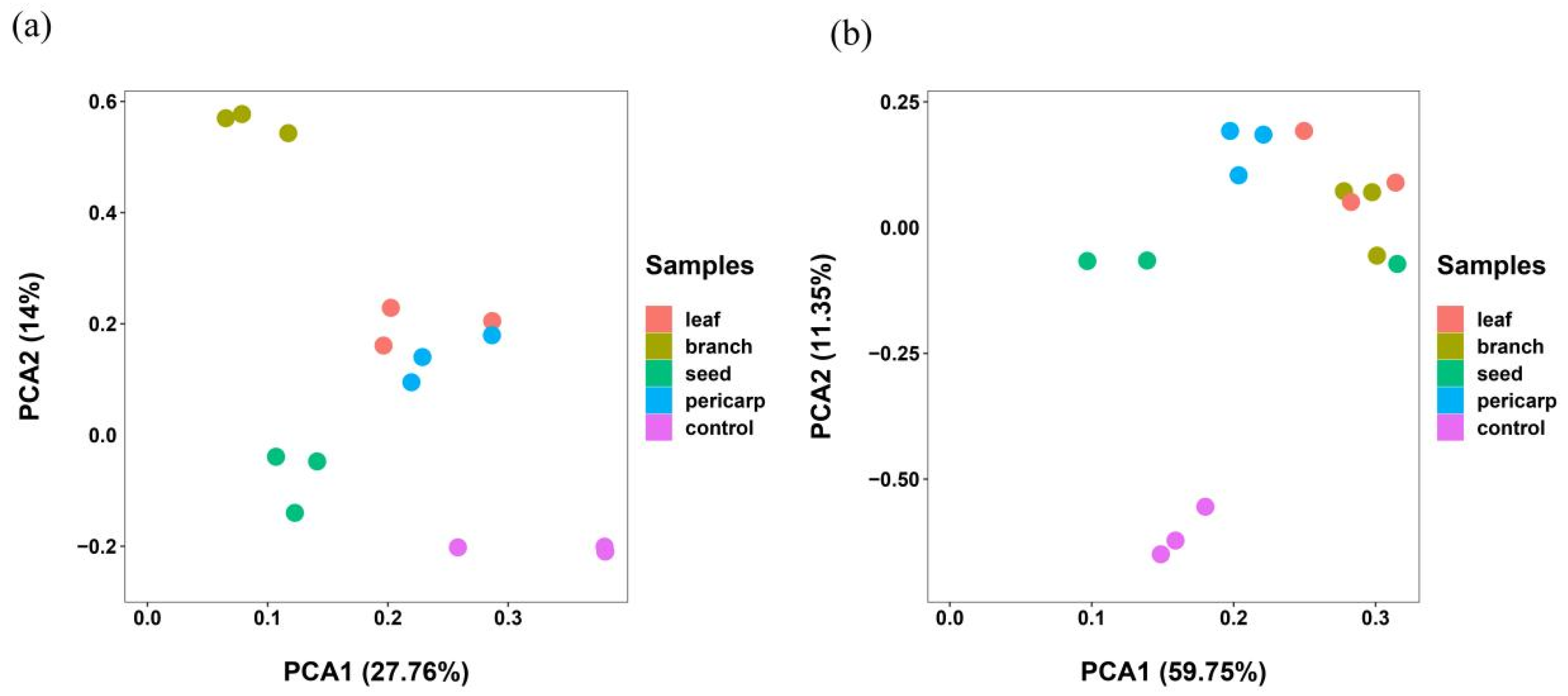
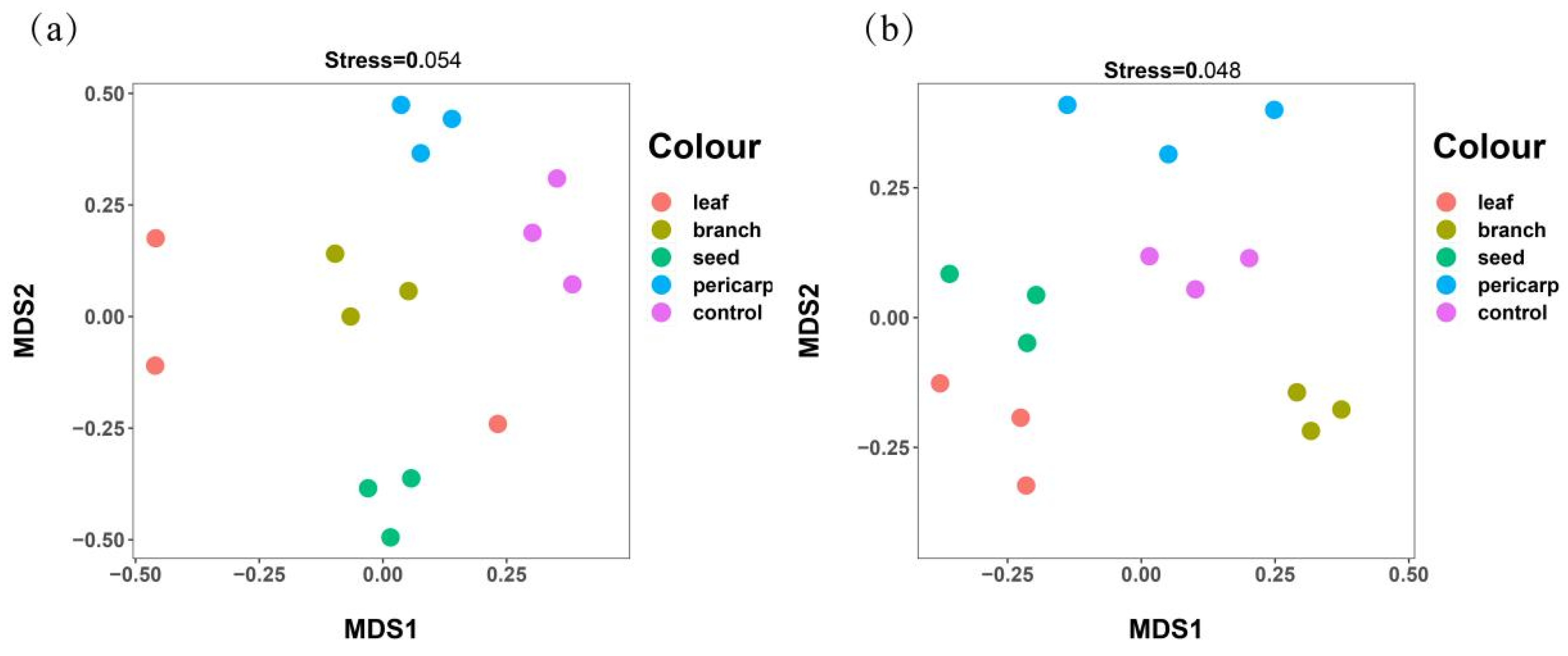
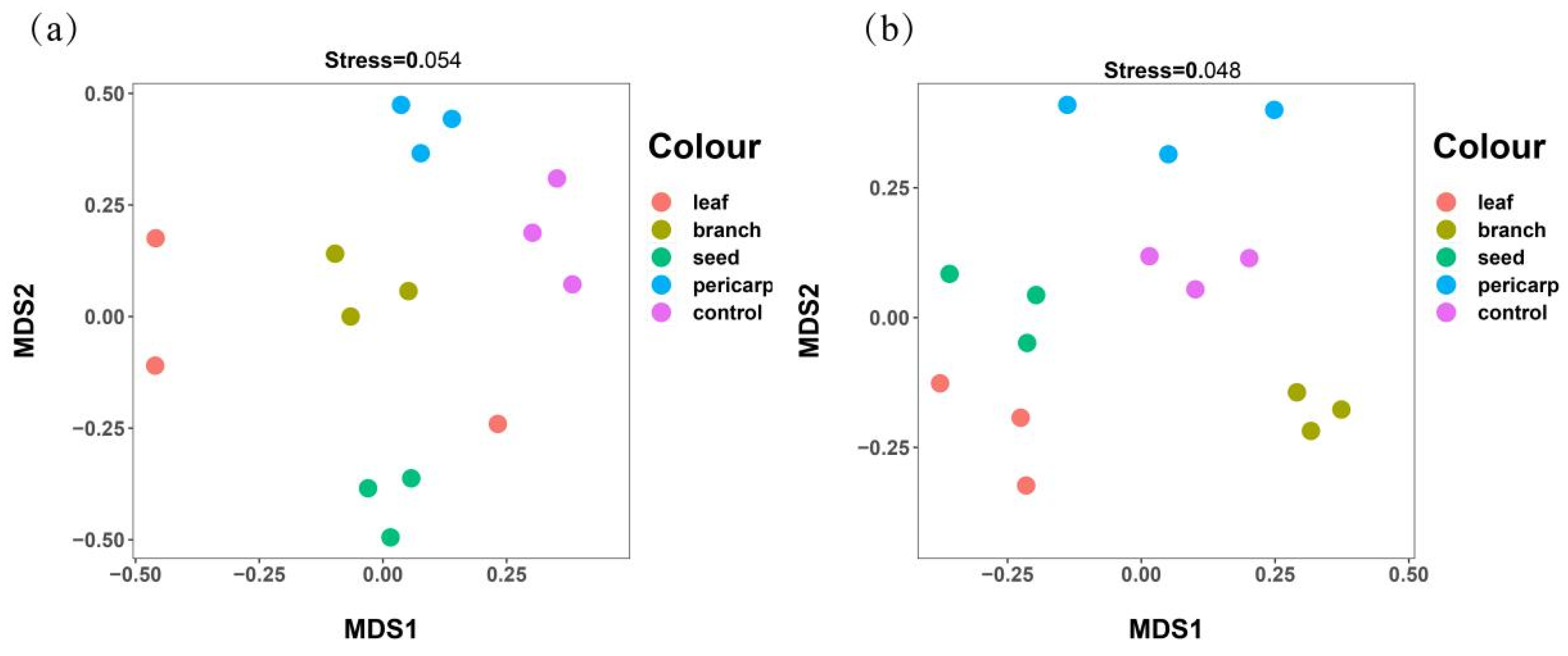
3.3. Beta Diversity Analysis
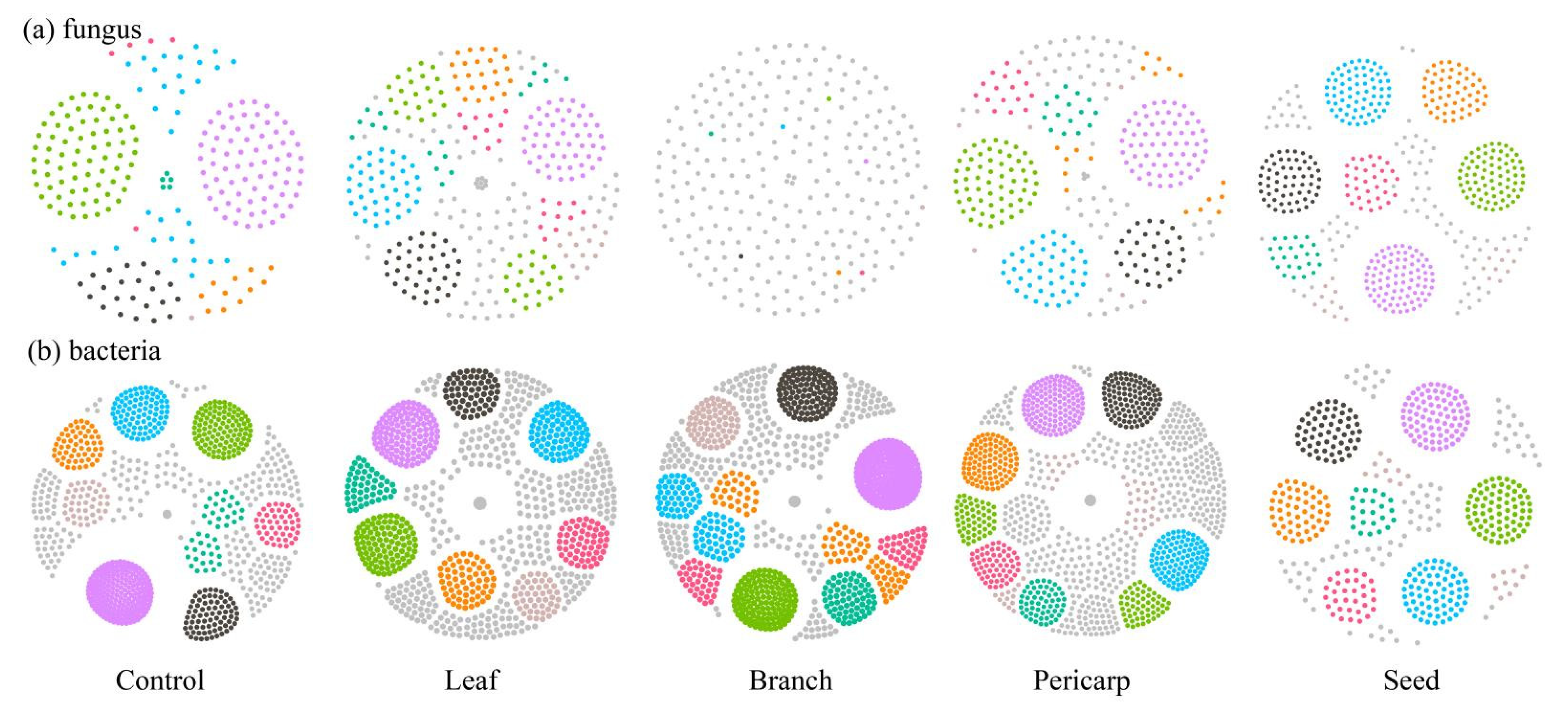
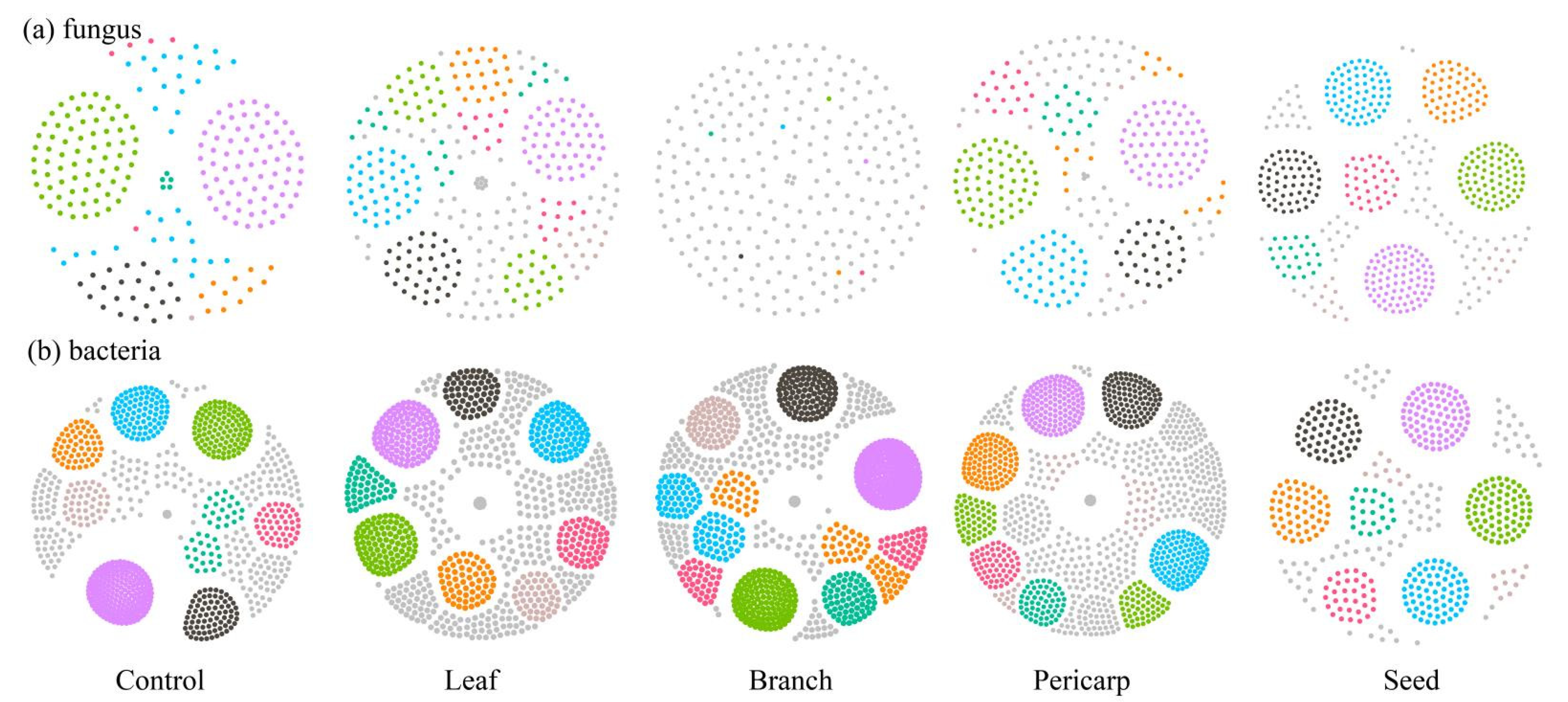
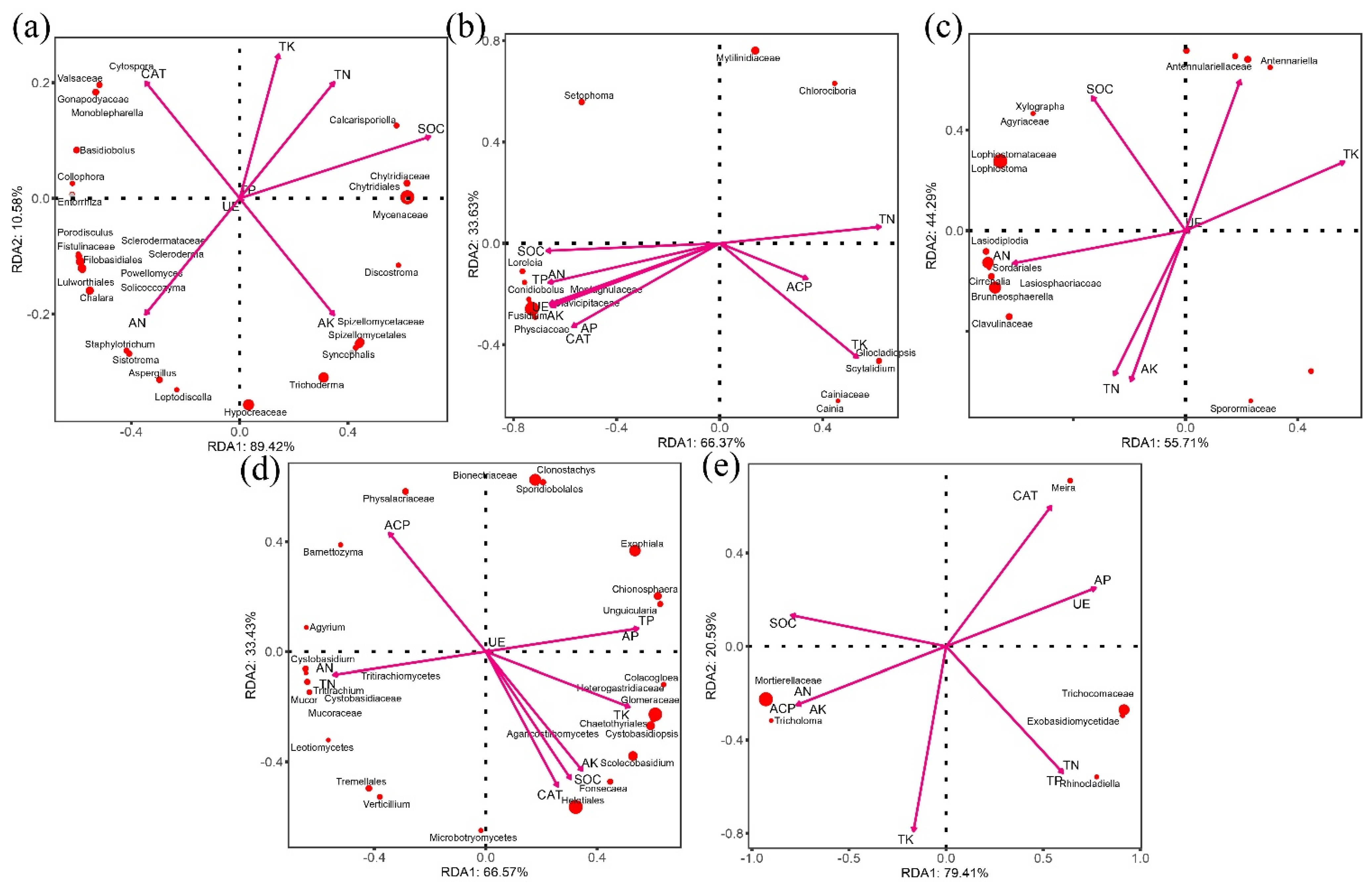
3.4. Microbial Ecological Network Construction and Correlation Analysis of Soil Chemical Properties in Different Ttreatments
4. Discussion
4.1. Effects of Litter Mulching with Different C. migao Tissues on the Chemical Properties of Topsoil
4.2. Effects of C. migao Litter Mulching on Topsoil Microbes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krishna, M.P.; Mohan, M. Litter decomposition in forest ecosystems: A review. Energy Ecol. Environ. 2017, 2, 236–249. [Google Scholar] [CrossRef]
- Giweta, M. Role of litter production and its decomposition, and factors affecting the processes in a tropical forest ecosystem: A review. J. Ecol. Environ. 2020, 44, 2–9. [Google Scholar] [CrossRef]
- Xiao, W.; Chen, H.Y.H.; Kumar, P.; Chen, C.; Guan, Q. Multiple interactions between tree composition and diversity and microbial diversity underly litter decomposition. Geoderma 2019, 341, 161–171. [Google Scholar] [CrossRef]
- Wang, J.H.; Zhang, N.; Wang, J.; Wu, K.Y. Main nutritional components in Cinnamomum migao seed. Guizhou Agric Sci. 2016, 44, 117–119. [Google Scholar]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The microbial efficiency-m atrix stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol. Biochem. 2019, 42, 391–404. [Google Scholar] [CrossRef]
- Kellner, H.; Luis, P.; Schlitt, B.; Buscot, F. Temporal changes in diversity and expression patterns of fungal laccase genes within the organic horizon of a brown forest soil. Soil Biol. Biochem. 2009, 41, 1380–1389. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; He, T.; Liu, L.; Wu, J. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Biol. Biochem. 2014, 71, 13–20. [Google Scholar] [CrossRef]
- Guo, C.; Dannenmann, M.; Gasche, R.; Zeller, B.; Papen, H. Preferential use of root litter compared to leaf litter by beech seedlings and soil microorganisms. Plant Soil 2013, 368, 519–534. [Google Scholar] [CrossRef]
- Sandhu, J.; Sinha, M.; Ambasht, R.S. Nitrogen release from decomposing litter of Leucaena leucocephala in the dry tropics. Pergamon 1990, 22, 859–863. [Google Scholar] [CrossRef]
- Suriani, M.; Bengen, D.G.; Prartono, T. The Production of Organic Matter from Rhizophora mucronata and Sonneratia alba at the Kajhu and Meunasah Mesjid Villages, Aceh Besar. Omni-Akuatika 2017, 13, 43–53. [Google Scholar] [CrossRef]
- Zhao, S.; Li, H.H.; Qiu, D.W.; Liu, Z.Y.; Liu, N. Resources, ecological investigation of Cinnamomum migao in Guizhou, Northern Guangxi, Hunan. J. GuiZhou Univ. Trad. Chin. Med. 1991, 3, 59–61. [Google Scholar]
- Chen, J.; Huang, X.; Tong, B.; Wang, D.; Liu, J.; Liao, X.; Sun, Q. Effects of rhizosphere fungi on the chemical composition of fruits of the medicinal plant Cinnamomum migao endemic to southwestern China. BMC Microbiol. 2021, 21, 206. [Google Scholar] [CrossRef] [PubMed]
- Pol, R.G.; Sagario, M.C.; Marone, L. Grazing impact on desert plants and soil seed banks: Implications for seed eating animals. Acta Oecol. 2014, 55, 58–65. [Google Scholar] [CrossRef]
- Lavee, S. Alternate bearing in olive initiated by abiotic induction leading to biotic responses. Adv. Hortic. Sci. 2015, 29, 213–220. [Google Scholar]
- Martínez-Alcántara, B.; Iglesias, D.J.; Reig, C.; Mesejo, C.; Agustí, M. Carbon utilization by fruit limits shoot growth in alternate-bearing citrus trees. J. Plant Physiol. 2015, 176, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.W. Physiological and Molecular Responses of “FUJI” Apple (Malus Domestica Borkh.) to GA3 Application and Alternate Bearing Signal during Floral Bud Differentiation. Master’s Thesis, Northwest University of Agriculture and Forestry Science and Technology, Xianyang, China, 2016. [Google Scholar]
- Apriyanto, E.; Hidayat, F.; Nugroho, P.B.; Tarigan, I. Litterfall production and Decomposition in three types of land use in bengkulu protection forest. J. Agro Sci. 2021, 9, 36–41. [Google Scholar] [CrossRef]
- Ni, X.; Lin, C.; Chen, G.; Xie, J.; Yang, Y. Decline in nutrient inputs from litterfall following forest plantation in subtropical China. For. Ecol. Manag. 2021, 496, 119445. [Google Scholar] [CrossRef]
- Liu, G.; Liu, Y.; Kang, X. Content Determination of Total Flavonoids and Sugar Components in Cinnamomum migao. J. Anhui Agri. Sci. 2017, 45, 141–144. [Google Scholar] [CrossRef]
- Huang, K.; Liu, J.; Huang, C.H.; Liu, Y.; Cheng, C.; Zhang, Y.P.; Xu, J. Comparative analysis of volatile oil and fatty oil constituents from Cinnamomum migao in different sources. China Phar. 2020, 31, 1961–1966. [Google Scholar]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, D.W.; Turner, J. Tamm Review: Nutrient cycling in forests: A historical look and newer developments. For. Ecol. Manag. 2019, 444, 344–373. [Google Scholar] [CrossRef]
- Sayer, E.J.; Rodtassana, C.; Sheldrake, M.; Bréchet, L.M.; Tanner, E. Revisiting nutrient cycling by litterfall-Insights from 15 years of litter manipulation in old-growth lowland tropical forest. Adv. Ecol. Res. 2020, 62, 173–223. [Google Scholar] [CrossRef]
- Liu, J.; Wu, N.; Hui, W.; Sun, J.; Bo, P. Nitrogen addition affects chemical compositions of plant tissues, litter and soil organic matter. Ecology 2016, 97, 1796–1806. [Google Scholar] [CrossRef]
- Kooch, Y.; Samadzadeh, B.; Hosseini, S.M. The effects of broad-leaved tree species on litter quality and soil properties in a plain forest stand. Catena 2017, 150, 223–229. [Google Scholar] [CrossRef]
- Carolina, C.S.; Olk, D.C.; Dietzel, R.N.; Mueller, K.E.; Archontouilis, S.V. Plant litter quality affects the accumulation rate, composition, and stability of mineral-associated soil organic matter. Soil Biol. Biochem. 2018, 125, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Freschet, G.T.; Cornwell, W.K.; Wardle, D.A.; Elumeeva, T.G.; Liu, W.; Jackson, B.G.; Onipchenko, V.G. Linking litter decomposition of above-and below-ground tissue to plant-soil feedbacks worldwide. J. Ecol. 2013, 101, 943–952. [Google Scholar] [CrossRef]
- Hou, Q.; Wang, W.; Yang, Y.; Bian, C.; Jin, L.; Li, G.; Xiong, X. Rhizosphere microbial diversity and community dynamics during potato cultivation. Eur. J. Soil Biol. 2020, 98, 103176. [Google Scholar] [CrossRef]
- Guo, H.; Yao, J.; Cai, M.; Qian, Y.; Guo, Y.; Richnow, H.H. Effects of petroleum contamination on soil microbial numbers, metabolic activity and urease activity. Chemosphere 2012, 87, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Tian, K.; Kong, X.; Yuan, L.; Lin, H.; Tian, X. Priming effect of litter mineralization: The role of root exudate depends on its interactions with litter quality and soil condition. Plant Soil 2019, 40, 457–471. [Google Scholar] [CrossRef]
- Karki, H.; Bargali, K.; Bargali, S.S. Spatial and temporal trends in soil N-mineralization rates under the agroforestry systems in Bhabhar belt of Kumaun Himalaya, India. Agrofor. Syst. 2021, 95, 1603–1617. [Google Scholar] [CrossRef]
- Wang, N.; Li, L.; Dannenmann, M.; Luo, Y.; Wang, C. Seasonality of gross ammonification and nitrification altered by precipitation in a semi-arid grassland of Northern China. Soil Biol. Biochem. 2021, 154, 108146. [Google Scholar] [CrossRef]
- Ishaq, M.; Wei, L.; Muhammad, S.R.; Guang, L.L.; Syed, S.U.H. Guaiane-type sesquiterpenoids from Cinnamomum migao H. W. Li: And their anti-inflammatory activities. Phytochemistry 2021, 190, 1–10. [Google Scholar] [CrossRef]
- Chen, Y.; Ma, S.; Jiang, H.; Hu, Y.; Lu, X. Influences of litter diversity and soil moisture on soil microbial communities in decomposing mixed litter of alpine steppe species. Geoderma 2020, 377, 114577. [Google Scholar] [CrossRef]
- Joly, F.X.; Fromin, N.; Kiikkilä, O.; Hättenschwiler, S. Diversity of leaf litter leachates from temperate forest trees and its consequences for soil microbial activity. Biogeochemistry 2016, 129, 373–388. [Google Scholar] [CrossRef]
- Barberan, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2021, 6, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Henzi, S.P.; Lusseau, D.; Weingrill, T.; Schaik, C.; Barrett, L. Cyclicity in the structure of female baboon social networks. Behav. Ecol. Sociobiol. 2009, 63, 1015–1021. [Google Scholar] [CrossRef]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The ecology of the microbiome: Networks, competition, and stability. Science 2015, 350, 663–666. [Google Scholar] [CrossRef]
- Georgiou, K.; Abramoff, R.Z.; Harte, J.; Riley, W.J.; Torn, M.S. Microbial community-level regulation explains soil carbon responses to long-term litter manipulations. Nat. Commun. 2017, 1223, 1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, L.T.; Feng, Y.X.; Zhang, D.; Du, S.S. Comparative evaluation of the chemical composition and bioactivities of essential oils from four spice plants (Lauraceae) against stored-product insects. Ind. Crop. Prod. 2019, 140, 111640. [Google Scholar] [CrossRef]
- Francioli, D.; Rijsse, S.Q.; Ruijven, J.; Termorshuizen, A.J.; Cotton, T.E.A.; Dumbrell, A.J.; Raaijmakers, J.M.; Weigelt, A.; Mommer, L. Plant functional group drives the community structure of saprophytic fungi in a grassland biodiversity experiment. Plant Soil 2021, 461, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Khalid, M.; Du, B.; Tan, H.; Liu, X.; Hui, N. Phosphorus elevation erodes ectomycorrhizal community diversity and induces divergence of saprophytic community composition between vegetation types. Sci. Total Environ. 2021, 793, 148502. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Hodson, M.C.; Hall, B.D. Loss of the flagellum happened only once in the fungal lineage: Phylogenetic structure of kingdom fungi inferred from rna polymerase ii subunit genes. BMC Evol. Biol. 2006, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- Cooper, A.C.; Desjardin, D.E.; Perry, B.A. The genus Mycena (Basidiomycota, Agaricales, Mycenaceae) and allied genera from Republic of São Tomé and Príncipe, West Africa. Phytotaxa 2018, 383, 1–13. [Google Scholar] [CrossRef]
- Telagathoti, A.; Probst, M.; Peintner, U. Habitat, Snow-mulching and soil pH, affect the distribution and diversity of mortierellaceae species and their associations to bacteria. Front. Microbiol. 2021, 12, 669784. [Google Scholar] [CrossRef]
- Wang, J.; Ma, L.J.; Long, Z.H.; Min, W.; Hou, Z.A. Effects of straw biochar on soil microbial metabolism and bacterial community composition in drip-irrigated cotton field. Environ. Sci. 2020, 41, 420–429. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; Singh, R.; Eltis, L.D.; Mohn, W.W. Bacterial contributions to delignification and lignocellulose degradation in forest soils with metagenomic and quantitative stable isotope probing. ISME J. 2019, 3, 413–429. [Google Scholar] [CrossRef] [Green Version]
- Niepceron, M.; Martin, L.F.; Crampon, M.; Portet, K.F.; Akpa, V.M. GammaProteobacteria as a potential bioindicator of a multiple contamination by polycyclic aromatic hydrocarbons (PAHs) in agricultural soils. Environ. Pollut. 2013, 180, 199–205. [Google Scholar] [CrossRef]
- Ragot, S.A.; Kertesz, M.A.; Bünemann, E.K. PhoD alkaline phosphatase gene diversity in soil. Appl. Environ. Microbiol. 2015, 81, 7281. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Different Treatment | Chao1 | Shannon | Simpson |
|---|---|---|---|
| Leaf | 2208.71 ± 242.922 a | 8.98 ± 0.272 a | 0.99 ± 0.001 a |
| Branch | 2516.03 ± 44.451 a | 9.12 ± 0.123 a | 1.00 ± 0 a |
| Seed | 2208.95 ± 380.612 a | 8.43 ± 1.078 a | 0.99 ± 0.015 a |
| Pericarp | 2321.84 ± 122.665 a | 8.85 ± 0.04 a | 0.99 ± 0 a |
| Control | 2159.03 ± 51.306 a | 8.76 ± 0.085 a | 0.99 ± 0.001 a |
| Treatment | Ultraperipheral Node (%) | Peripheral Node (%) | Non-Hub Connectors (%) | Non-Hub Kinless Nodes (%) | Provincial Hubs (%) | Connector Hubs (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Community | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria |
| Control | 52.25 | 0.00 | 41.89 | 38.96 | 0.00 | 30.07 | 3.60 | 30.18 | 2.25 | 0.00 | 0.00 | 0.76 |
| Leaf | 0.00 | 0.00 | 74.37 | 80.06 | 0.00 | 0.63 | 23.42 | 17.72 | 0.00 | 0.00 | 2.22 | 1.58 |
| Branch | 0.00 | 0.00 | 80.49 | 94.18 | 2.44 | 0.00 | 15.854 | 4.50 | 0.00 | 1.31 | 1.22 | 0.00 |
| Pericarp | 0.00 | 49.02 | 3.38 | 46.42 | 80.84 | 0.00 | 14.68 | 3.57 | 0.00 | 0.97 | 1.10 | 0.00 |
| Seed | 50.18 | 2.25 | 43.17 | 28.48 | 0 | 29.91 | 5.16 | 39.13 | 1.47 | 0.00 | 0 | 0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Y.; Liao, X.; Liu, J.; Chen, J. A Case Study Demonstrates That the Litter of the Rare Species Cinnamomum migao Composed of Different Tissues Can Affect the Chemical Properties and Microbial Community Diversity in Topsoil. Microorganisms 2022, 10, 1125. https://doi.org/10.3390/microorganisms10061125
Xie Y, Liao X, Liu J, Chen J. A Case Study Demonstrates That the Litter of the Rare Species Cinnamomum migao Composed of Different Tissues Can Affect the Chemical Properties and Microbial Community Diversity in Topsoil. Microorganisms. 2022; 10(6):1125. https://doi.org/10.3390/microorganisms10061125
Chicago/Turabian StyleXie, Yuangui, Xiaofeng Liao, Jiming Liu, and Jingzhong Chen. 2022. "A Case Study Demonstrates That the Litter of the Rare Species Cinnamomum migao Composed of Different Tissues Can Affect the Chemical Properties and Microbial Community Diversity in Topsoil" Microorganisms 10, no. 6: 1125. https://doi.org/10.3390/microorganisms10061125
APA StyleXie, Y., Liao, X., Liu, J., & Chen, J. (2022). A Case Study Demonstrates That the Litter of the Rare Species Cinnamomum migao Composed of Different Tissues Can Affect the Chemical Properties and Microbial Community Diversity in Topsoil. Microorganisms, 10(6), 1125. https://doi.org/10.3390/microorganisms10061125