
Gut Microbiome: A Brief Review on Its Role in Schizophrenia and First Episode of Psychosis

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Gut Microbiome and Schizophrenia
2.1. Pathophysiological Mechanisms
2.2. Microbiome Diversity
2.3. Mediating Factors
2.4. Association with Clinical Characteristics of Schizophrenia
3. Gut Microbiome and First Episode of Psychosis
3.1. Alterations in Microbiome Composition
3.2. Pathophysiological Mechanisms in FEP and Association with Clinical Features and Prognosis of Psychosis
4. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nowakowski, M.E.; McCabe, R.; Rowa, K.; Pellizzari, J.; Surette, M.; Moayyedi, P.; Anglin, R. The gut microbiome: Potential innovations for the understanding and treatment of psychopathology. Can. Psychol. Psychol. Can. 2016, 57, 67–75. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Ravel, J. The vocabulary of microbiome research: A proposal. Microbiome 2015, 3, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherwin, E.; Dinan, T.G.; Cryan, J.F. Recent developments in understanding the role of the gut microbiota in brain health and disease. Ann. N. Y. Acad. Sci. 2018, 1420, 5–25. [Google Scholar] [CrossRef]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/brain axis and the microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. The impact of gut microbiota on brain and behaviour: Implications for psychiatry. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 552–558. [Google Scholar] [CrossRef]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W.J. Psychobiotics and the Manipulation of Bacteria-Gut-Brain Signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef] [Green Version]
- Kelly, J.R.; Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G.; Hyland, N.P. Breaking down the barriers: The gut microbiome, intestinal permeability and stress-related psychiatric disorders. Front. Cell. Neurosci. 2015, 9, 392. [Google Scholar] [CrossRef] [Green Version]
- Dinan, T.G.; Cryan, J.F. The Microbiome-Gut-Brain Axis in Health and Disease. Gastroenterol. Clin. N. Am. 2017, 46, 77–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, C.R.; Osadchiy, V.; Kalani, A.; Mayer, E.A. The Brain-Gut-Microbiome Axis. Cell. Mol. Gastroenterol. Hepatol. 2018, 6, 133–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ameringen, M.; Turna, J.; Patterson, B.; Pipe, A.; Mao, R.Q.; Anglin, R.; Surette, M.G. The gut microbiome in psychiatry: A primer for clinicians. Depress. Anxiety 2019, 36, 1004–1025. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Kosciolek, T.; Eyler, L.T.; Knight, R.; Jeste, D.V. Overview and systematic review of studies of microbiome in schizophrenia and bipolar disorder. J. Psychiatr. Res. 2018, 99, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Groen, R.N.; de Clercq, N.C.; Nieuwdorp, M.; Hoenders, H.J.R.; Groen, A.K. Gut microbiota, metabolism and psychopathology: A critical review and novel perspectives. Crit. Rev. Clin. Lab. Sci. 2018, 55, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Dicks, L.M.T.; Hurn, D.; Hermanus, D. Gut Bacteria and Neuropsychiatric Disorders. Microorganisms 2021, 9, 2583. [Google Scholar] [CrossRef]
- Borre, Y.E.; O’Keeffe, G.W.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota and neurodevelopmental windows: Implications for brain disorders. Trends Mol. Med. 2014, 20, 509–518. [Google Scholar] [CrossRef]
- Alander, M.; Satokari, R.; Korpela, R.; Saxelin, M.; Vilpponen-Salmela, T.; Mattila-Sandholm, T.; von Wright, A. Persistence of colonization of human colonic mucosa by a probiotic strain, Lactobacillus rhamnosus GG, after oral consumption. Appl. Environ. Microbiol. 1999, 65, 351–354. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.H.; Balakrishnan, K.; Thiagarajah, K.; Mohd Ismail, N.I.; Yin, O.S. Beneficial Properties of Probiotics. Trop. Life Sci. Res. 2016, 27, 73–90. [Google Scholar] [CrossRef]
- Szeligowski, T.; Yun, A.L.; Lennox, B.R.; Burnet, P.W.J. The Gut Microbiome and Schizophrenia: The Current State of the Field and Clinical Applications. Front. Psychiatry 2020, 11, 156. [Google Scholar] [CrossRef] [Green Version]
- Derrien, M.; van Hylckama Vlieg, J.E. Fate, activity, and impact of ingested bacteria within the human gut microbiota. Trends Microbiol. 2015, 23, 354–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frei, R.; Akdis, M.; O’Mahony, L. Prebiotics, probiotics, synbiotics, and the immune system: Experimental data and clinical evidence. Curr. Opin. Gastroenterol. 2015, 31, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zommiti, M.; Feuilloley, M.G.J.; Connil, N. Update of Probiotics in Human World: A Nonstop Source of Benefactions till the End of Time. Microorganisms 2020, 8, 1907. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, M.; Lalonde, R.; Violle, N.; Javelot, H.; Desor, D.; Nejdi, A.; Bisson, J.F.; Rougeot, C.; Pichelin, M.; Cazaubiel, M.; et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br. J. Nutr. 2011, 105, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, Q.X.; Peters, C.; Ho, C.Y.X.; Lim, D.Y.; Yeo, W.S. A meta-analysis of the use of probiotics to alleviate depressive symptoms. J. Affect. Disord. 2018, 228, 13–19. [Google Scholar] [CrossRef]
- Ng, Q.X.; Soh, A.Y.S.; Venkatanarayanan, N.; Ho, C.Y.X.; Lim, D.Y.; Yeo, W.S. A Systematic Review of the Effect of Probiotic Supplementation on Schizophrenia Symptoms. Neuropsychobiology 2019, 78, 1–6. [Google Scholar] [CrossRef]
- Alagiakrishnan, K.; Halverson, T. Microbial Therapeutics in Neurocognitive and Psychiatric Disorders. J. Clin. Med. Res. 2021, 13, 439–459. [Google Scholar] [CrossRef]
- Desai, V.; Kozyrskyj, A.L.; Lau, S.; Sanni, O.; Dennett, L.; Walter, J.; Ospina, M.B. Effectiveness of Probiotic, Prebiotic, and Synbiotic Supplementation to Improve Perinatal Mental Health in Mothers: A Systematic Review and Meta-Analysis. Front. Psychiatry 2021, 12, 622181. [Google Scholar] [CrossRef]
- Chudzik, A.; Orzyłowska, A.; Rola, R.; Stanisz, G.J. Probiotics, Prebiotics and Postbiotics on Mitigation of Depression Symptoms: Modulation of the Brain-Gut-Microbiome Axis. Biomolecules 2021, 11, 1000. [Google Scholar] [CrossRef]
- Le Morvan de Sequeira, C.; Hengstberger, C.; Enck, P.; Mack, I. Effect of Probiotics on Psychiatric Symptoms and Central Nervous System Functions in Human Health and Disease: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 621. [Google Scholar] [CrossRef]
- Simpson, C.A.; Diaz-Arteche, C.; Eliby, D.; Schwartz, O.S.; Simmons, J.G.; Cowan, C.S.M. The gut microbiota in anxiety and depression—A systematic review. Clin. Psychol. Rev. 2021, 83, 101943. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.C.; Oliveira, J.; Silva, S.; Madeira, N.; Pereira, C.M.F.; Cruz, M.T. Inflammation in Bipolar Disorder (BD): Identification of new therapeutic targets. Pharmacol. Res. 2021, 163, 105325. [Google Scholar] [CrossRef] [PubMed]
- Munawar, N.; Ahsan, K.; Muhammad, K.; Ahmad, A.; Anwar, M.A.; Shah, I.; Al Ameri, A.K.; Al Mughairbi, F. Hidden Role of Gut Microbiome Dysbiosis in Schizophrenia: Antipsychotics or Psychobiotics as Therapeutics? Int. J. Mol. Sci. 2021, 22, 7671. [Google Scholar] [CrossRef] [PubMed]
- Minichino, A.; Brondino, N.; Solmi, M.; Del Giovane, C.; Fusar-Poli, P.; Burnet, P.; Cipriani, A.; Lennox, B.R. The gut-microbiome as a target for the treatment of schizophrenia: A systematic review and meta-analysis of randomised controlled trials of add-on strategies. Schizophr. Res. 2021, 234, 58–70. [Google Scholar] [CrossRef]
- Alnæs, D.; Kaufmann, T.; van der Meer, D.; Córdova-Palomera, A.; Rokicki, J.; Moberget, T.; Bettella, F.; Agartz, I.; Barch, D.M.; Bertolino, A.; et al. Brain Heterogeneity in Schizophrenia and Its Association With Polygenic Risk. JAMA Psychiatry 2019, 76, 739–748. [Google Scholar] [CrossRef]
- Fusar-Poli, P.; McGorry, P.D.; Kane, J.M. Improving outcomes of first-episode psychosis: An overview. World Psychiatry 2017, 16, 251–265. [Google Scholar] [CrossRef]
- Severance, E.G.; Gressitt, K.L.; Stallings, C.R.; Origoni, A.E.; Khushalani, S.; Leweke, F.M.; Dickerson, F.B.; Yolken, R.H. Discordant patterns of bacterial translocation markers and implications for innate immune imbalances in schizophrenia. Schizophr. Res. 2013, 148, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.; Berk, M.; Dodd, S.; Bechter, K.; Altamura, A.C.; Dell’osso, B.; Kanba, S.; Monji, A.; Fatemi, S.H.; Buckley, P.; et al. Immuno-inflammatory, oxidative and nitrosative stress, and neuroprogressive pathways in the etiology, course and treatment of schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 42, 1–4. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Kamada, N.; Seo, S.U.; Chen, G.Y.; Núñez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef]
- Marazziti, D.; Buccianelli, B.; Palermo, S.; Parra, E.; Arone, A.; Beatino, M.F.; Massa, L.; Carpita, B.; Barberi, F.M.; Mucci, F. The Microbiota/Microbiome and the Gut-Brain Axis: How Much Do They Matter in Psychiatry? Life 2021, 11, 760. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Guo, R.; Wang, W.; Ju, Y.; Wang, Q.; Ma, Q.; Sun, Q.; Fan, Y.; Xie, Y.; Yang, Z.; et al. Transplantation of microbiota from drug-free patients with schizophrenia causes schizophrenia-like abnormal behaviors and dysregulated kynurenine metabolism in mice. Mol. Psychiatry 2020, 25, 2905–2918. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, S.; Schwieler, L.; Imbeault, S.; Engberg, G. The kynurenine pathway in schizophrenia and bipolar disorder. Neuropharmacology 2017, 112, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Zeng, B.; Liu, M.; Chen, J.; Pan, J.; Han, Y.; Liu, Y.; Cheng, K.; Zhou, C.; Wang, H.; et al. The gut microbiome from patients with schizophrenia modulates the glutamate-glutamine-GABA cycle and schizophrenia-relevant behaviors in mice. Sci. Adv. 2019, 5, eaau8317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stan, A.D.; Ghose, S.; Zhao, C.; Hulsey, K.; Mihalakos, P.; Yanagi, M.; Morris, S.U.; Bartko, J.J.; Choi, C.; Tamminga, C.A. Magnetic resonance spectroscopy and tissue protein concentrations together suggest lower glutamate signaling in dentate gyrus in schizophrenia. Mol. Psychiatry 2015, 20, 433–439. [Google Scholar] [CrossRef]
- McCutcheon, R.A.; Krystal, J.H.; Howes, O.D. Dopamine and glutamate in schizophrenia: Biology, symptoms and treatment. World Psychiatry 2020, 19, 15–33. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.; Hathaway, H.; Kosciolek, T.; Knight, R.; Jeste, D.V. Gut microbiome in serious mental illnesses: A systematic review and critical evaluation. Schizophr. Res. 2021, 234, 24–40. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Xu, J.; Li, Z.; Huang, Y.; Yuan, Y.; Wang, J.; Zhang, M.; Hu, S.; Liang, Y. Analysis of gut microbiota diversity and auxiliary diagnosis as a biomarker in patients with schizophrenia: A cross-sectional study. Schizophr. Res. 2018, 197, 470–477. [Google Scholar] [CrossRef]
- Li, S.; Song, J.; Ke, P.; Kong, L.; Lei, B.; Zhou, J.; Huang, Y.; Li, H.; Li, G.; Chen, J.; et al. The gut microbiome is associated with brain structure and function in schizophrenia. Sci. Rep. 2021, 11, 9743. [Google Scholar] [CrossRef]
- Ma, X.; Asif, H.; Dai, L.; He, Y.; Zheng, W.; Wang, D.; Ren, H.; Tang, J.; Li, C.; Jin, K.; et al. Alteration of the gut microbiome in first-episode drug-naïve and chronic medicated schizophrenia correlate with regional brain volumes. J. Psychiatr. Res. 2020, 123, 136–144. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Phillips, R.; Sarnyai, Z. The Gut Microbiome in Psychosis From Mice to Men: A Systematic Review of Preclinical and Clinical Studies. Front. Psychiatry 2020, 11, 799. [Google Scholar] [CrossRef] [PubMed]
- Nocera, A.; Nasrallah, H.A. The Association of the Gut Microbiota with Clinical Features in Schizophrenia. Behav. Sci. 2022, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Kosciolek, T.; Maldonado, Y.; Daly, R.E.; Martin, A.S.; McDonald, D.; Knight, R.; Jeste, D.V. Differences in gut microbiome composition between persons with chronic schizophrenia and healthy comparison subjects. Schizophr. Res. 2019, 204, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ni, J.; Qing, H. Gut Microbiota: Critical Controller and Intervention Target in Brain Aging and Cognitive Impairment. Front. Aging Neurosci. 2021, 13, 671142. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.L.; Xu, J.; Rong, X.Y.; Wang, F.; Wang, H.J.; Zhao, C. Gut microbiota alterations and health status in aging adults: From correlation to causation. Aging Med. 2021, 4, 206–213. [Google Scholar] [CrossRef]
- Nosarti, C.; Reichenberg, A.; Murray, R.M.; Cnattingius, S.; Lambe, M.P.; Yin, L.; MacCabe, J.; Rifkin, L.; Hultman, C.M. Preterm birth and psychiatric disorders in young adult life. Arch. Gen. Psychiatry 2012, 69, E1–E8. [Google Scholar] [CrossRef] [Green Version]
- Tamburini, S.; Shen, N.; Wu, H.C.; Clemente, J.C. The microbiome in early life: Implications for health outcomes. Nat. Med. 2016, 22, 713–722. [Google Scholar] [CrossRef] [Green Version]
- Strassnig, M.; Brar, J.S.; Ganguli, R. Dietary intake of patients with schizophrenia. Psychiatry 2005, 2, 31–35. [Google Scholar]
- Hartz, S.M.; Pato, C.N.; Medeiros, H.; Cavazos-Rehg, P.; Sobell, J.L.; Knowles, J.A.; Bierut, L.J.; Pato, M.T. Comorbidity of severe psychotic disorders with measures of substance use. JAMA Psychiatry 2014, 71, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Vancampfort, D.; Stubbs, B.; Sienaert, P.; Wyckaert, S.; De Hert, M.; Soundy, A.; Probst, M. A comparison of physical fitness in patients with bipolar disorder, schizophrenia and healthy controls. Disabil. Rehabil. 2016, 38, 2047–2051. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, E.; Maukonen, J.; Hyytiäinen, T.; Kieseppä, T.; Orešič, M.; Sabunciyan, S.; Mantere, O.; Saarela, M.; Yolken, R.; Suvisaari, J. Analysis of microbiota in first episode psychosis identifies preliminary associations with symptom severity and treatment response. Schizophr. Res. 2018, 192, 398–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyndt Jørgensen, B.; Krych, L.; Pedersen, T.B.; Plath, N.; Redrobe, J.P.; Hansen, A.K.; Nielsen, D.S.; Pedersen, C.S.; Larsen, C.; Sørensen, D.B. Investigating the long-term effect of subchronic phencyclidine-treatment on novel object recognition and the association between the gut microbiota and behavior in the animal model of schizophrenia. Physiol. Behav. 2015, 141, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Dunphy-Doherty, F.; O’Mahony, S.M.; Peterson, V.L.; O’Sullivan, O.; Crispie, F.; Cotter, P.D.; Wigmore, P.; King, M.V.; Cryan, J.F.; Fone, K.C.F. Post-weaning social isolation of rats leads to long-term disruption of the gut microbiota-immune-brain axis. Brain Behav. Immun. 2018, 68, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Ghaderi, A.; Banafshe, H.R.; Mirhosseini, N.; Moradi, M.; Karimi, M.A.; Mehrzad, F.; Bahmani, F.; Asemi, Z. Clinical and metabolic response to vitamin D plus probiotic in schizophrenia patients. BMC Psychiatry 2019, 19, 77. [Google Scholar] [CrossRef] [Green Version]
- Okubo, R.; Koga, M.; Katsumata, N.; Odamaki, T.; Matsuyama, S.; Oka, M.; Narita, H.; Hashimoto, N.; Kusumi, I.; Xiao, J.; et al. Effect of bifidobacterium breve A-1 on anxiety and depressive symptoms in schizophrenia: A proof-of-concept study. J. Affect. Disord. 2019, 245, 377–385. [Google Scholar] [CrossRef]
- He, Y.; Kosciolek, T.; Tang, J.; Zhou, Y.; Li, Z.; Ma, X.; Zhu, Q.; Yuan, N.; Yuan, L.; Li, C.; et al. Gut microbiome and magnetic resonance spectroscopy study of subjects at ultra-high risk for psychosis may support the membrane hypothesis. Eur. Psychiatry 2018, 53, 37–45. [Google Scholar] [CrossRef]
- Yuan, X.; Zhang, P.; Wang, Y.; Liu, Y.; Li, X.; Kumar, B.U.; Hei, G.; Lv, L.; Huang, X.F.; Fan, X.; et al. Changes in metabolism and microbiota after 24-week risperidone treatment in drug naïve, normal weight patients with first episode schizophrenia. Schizophr. Res. 2018, 201, 299–306. [Google Scholar] [CrossRef]
- Zhang, X.; Pan, L.Y.; Zhang, Z.; Zhou, Y.Y.; Jiang, H.Y.; Ruan, B. Analysis of gut mycobiota in first-episode, drug-naïve Chinese patients with schizophrenia: A pilot study. Behav. Brain Res. 2020, 379, 112374. [Google Scholar] [CrossRef]
- Zhu, F.; Ju, Y.; Wang, W.; Wang, Q.; Guo, R.; Ma, Q.; Sun, Q.; Fan, Y.; Xie, Y.; Yang, Z.; et al. Metagenome-wide association of gut microbiome features for schizophrenia. Nat. Commun. 2020, 11, 1612. [Google Scholar] [CrossRef] [Green Version]
- Buckley, P.F.; Correll, C.U.; Miller, A.L. First-episode psychosis: A window of opportunity for best practices. CNS Spectr. 2007, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.; Armet, A.M.; Finlay, B.B.; Shanahan, F. Establishing or Exaggerating Causality for the Gut Microbiome: Lessons from Human Microbiota-Associated Rodents. Cell 2020, 180, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Mayneris-Perxachs, J.; Castells-Nobau, A.; Arnoriaga-Rodríguez, M.; Martin, M.; de la Vega-Correa, L.; Zapata, C.; Burokas, A.; Blasco, G.; Coll, C.; Escrichs, A.; et al. Microbiota alterations in proline metabolism impact depression. Cell Metab. 2022, 34, 681–701.e10. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Microbiome Changes | Pathophysiological Mechanisms | |
|---|---|---|
| Schizophrenia |
|
|
| First episode of psychosis (FEP) |
|
|
| First Author/Year | Type of Study | Findings and Comments |
|---|---|---|
| Zhu, Guo, 2020 [42] | Animal (mice) | FMT from schizophrenic patients into mice treated with antibiotics resulted in psychomotor hyperactivity and impaired learning and memory. Elevation of the kynurenine–kynurenic acid pathway of tryptophan degradation in both the central and peripheral nervous systems. Elevated basal extracellular dopamine in prefrontal cortex and 5-HT in hippocampus. |
| Zheng, 2019 [44] | Animal (mice) and Human (schizophrenic vs. controls) | Altered amino acid and lipid metabolisms, along with disruptions in the glutamate–glutamine–GABA cycle and decreased brain glutamate. Veillonellaceae OTU191 had a negative correlation with PANNS scores, whereas Bacteroidaceae OTU172, Streptococcaceae OTU834 and two Lachnospiraceae OTUs (477 and 629) had a positive correlation with PANSS scores. |
| Li, 2021 [49] | Human (schizophrenic vs. controls) | Relative abundance of Ruminococcus and Roseburia was significantly reduced at the genus level, while the abundance of Veillonella was significantly increased in schizophrenic patients compared to the controls. |
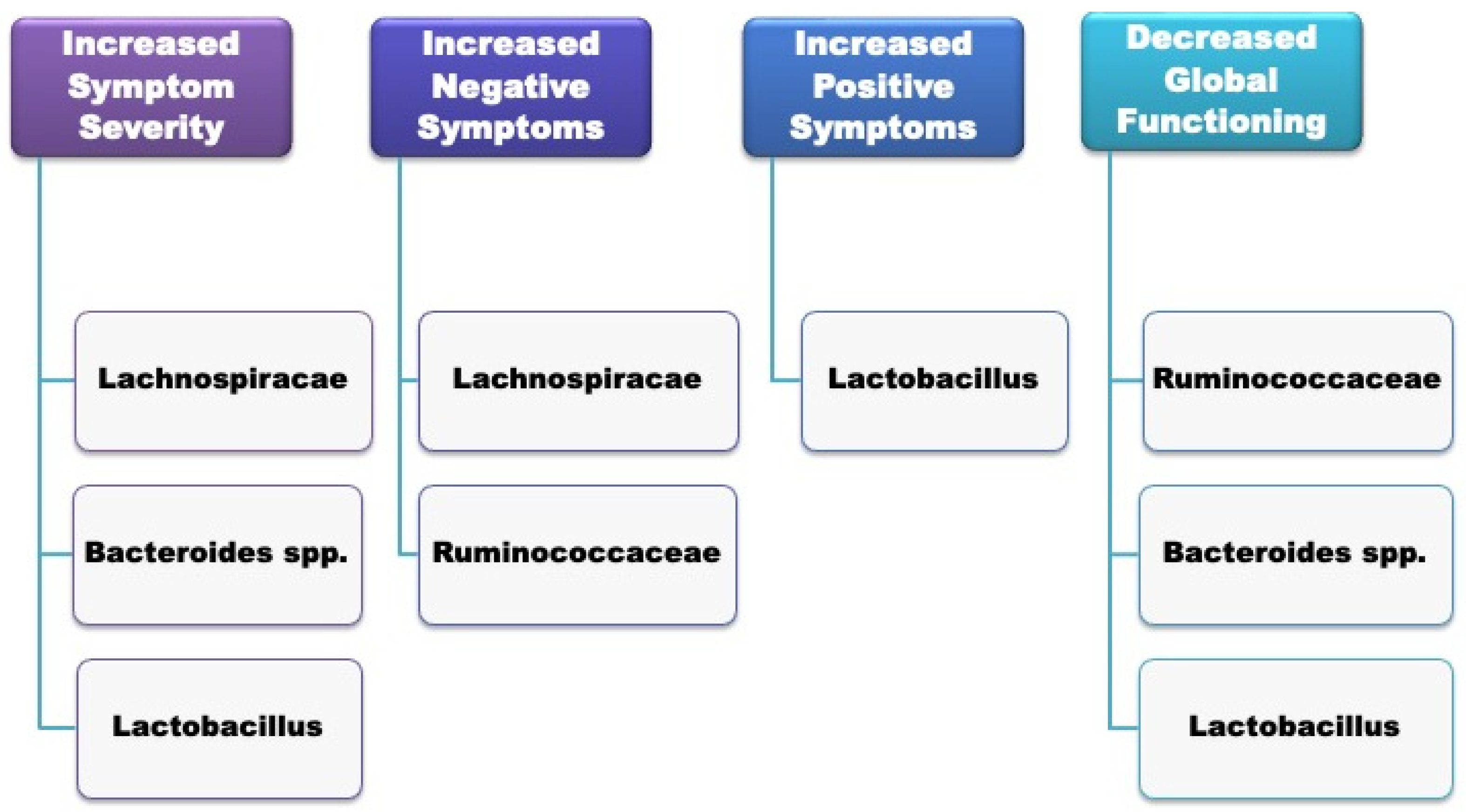
| Nguyen, 2019 [53] | Human (schizophrenic vs. controls) | Proteobacteria were relatively lower in schizophrenic subjects compared to the controls. At the genus level, Anaerococcus was relatively higher in schizophrenic subjects, whereas Haemophilus, Sutterella and Clostridium were reduced. In schizophrenic patients, the abundance of Ruminococcaceae was associated with reduced severity of negative symptoms. Bacteroides was correlated with more severe depressive symptoms. |
| Shen, 2018 [48] | Human (schizophrenic vs. controls) | Abundance of Proteobacteria (at the phylum level) was significantly higher in schizophrenic subjects. At the genus level, the relative abundance of Succinivibrio, Megasphaera, Collinsella, Clostridium, Klebsiella and Methanobrevibacter was significantly increased, while the abundance of Blautia, Coprococcus and Roseburia was reduced. Numerous metabolic pathways were significantly different between the healthy controls and schizophrenic subjects, such as vitamin B6 and fatty acid. |
| Ma, 2020 [50] | Human (FEP vs. schizophrenic vs. controls) | Both first-episode psychotic (FSCZ) patients, and chronically antipsychotic-treated schizophrenic subjects (TSCZ) had marked changes in gut microbiome composition in certain taxa, including Christensenellaceae, Enterobacteriaceae, Pasteurellaceae, Turicibacteraceae at the family level and Escherichia at the genus level. Major disturbances in the gut microbiome composition in TSCZ compared to FSCZ patients (eg. Enterococcaceae and Lactobacillaceae). Certain schizophrenia-related microbiota correlated with the right middle frontal gyrus volume, which was abnormal in schizophrenic subjects. |
| Pyndt Jørgensen, 2015 [63] | Animal (rats) | Hyperactivity linked to an elevation in Lachnospiraceae and Clostridiaceae. At the genus level, it was related to increased Roseburia, Clostridium and Odoribacter. |
| Dunphy-Doherty, 2018 [64] | Animal (rats) | Socially isolated rats had altered microbiota composition with elevated Actinobacteria and reduced Clostridia class compared to controls. Differences were also observed at the genus level. Positive correlations were seen between microbiota and hippocampal IL-6 and IL-10, conditioned freezing and open-field exploration. Adverse early life stress resulting from continuous social isolation increased ‘anxiety-like’ behaviour and impaired associative learning and memory that went along with alterations to gut microbiota, reduced hippocampal IL-6 and IL-10 and neurogenesis. |
| He, 2018 [67] | Human (high-risk (HR) subjects vs. ultra-high-risk (UHR) subjects vs. health controls (HC)) | Increased Clostridiales, Lactobacillales and Bacteroidales were noted in the faecal samples of UHR subjects compared to the other two groups. Increased production of short-chain fatty acids (SCFAs), as indicated by changes in microbiota composition, which can lead to the activation of microglia and disruption of membrane metabolism. |
| Yuan, 2018 [68] | Human (FEP vs. healthy controls (HC)) | FEP subjects had significantly reduced numbers of faecal Bifidobacterium spp., Escherichia coli and Lactobacillus spp. Significantly higher numbers of faecal Clostridium coccoides group in the patient group. After 24-week risperidone treatment, significant increases were noted in body weight, BMI, fasting blood glucose, triglycerides, LDL and a major elevation in the numbers of fecal Bifidobacterium spp. and E. coli. Additionally, significant decreases in the numbers of faecal Clostridium coccoides group and Lactobacillus spp. |
| Zhang, 2018 [69] | Human (schizophrenic (SC) vs. healthy controls (HC)) | Increased abundance of harmful bacterial (Proteobacteria) and decreased short-chain fatty acid (SCFA)-producing bacteria, such as the Faecalibacterium and Lachnospiraceae genera in schizophrenic subjects. Relative reduction in alpha diversity and altered composition in the gut mycobiota. Higher levels of Chaetomium and a lower level of Trichoderma in the schizophrenic group. |
| Zhu, Ju, 2020 [70] | Human (medication-free psychotic vs. controls) and animals (mice) | Psychotic subjects had a higher alpha diversity and higher beta diversity. Differences in short-chain fatty acid synthesis, tryptophan metabolism and synthesis/degradation of neurotransmitters associated with schizophrenia. FMT of a schizophrenia-enriched bacterium, Streptococcus vestibularis, induced deficits in social behaviours and altered neurotransmitter levels in peripheral tissues. |
| Schwarz, 2018 [62] | Human (FEP vs. healthy controls) | Elevated Lactobacillus group bacteria in FEP-subjects that significantly correlated with severity along different symptom domains. FEP subjects with the highest microbiome alterations showed a poorer response after up to 12 months of treatment. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsamakis, K.; Galinaki, S.; Alevyzakis, E.; Hortis, I.; Tsiptsios, D.; Kollintza, E.; Kympouropoulos, S.; Triantafyllou, K.; Smyrnis, N.; Rizos, E. Gut Microbiome: A Brief Review on Its Role in Schizophrenia and First Episode of Psychosis. Microorganisms 2022, 10, 1121. https://doi.org/10.3390/microorganisms10061121
Tsamakis K, Galinaki S, Alevyzakis E, Hortis I, Tsiptsios D, Kollintza E, Kympouropoulos S, Triantafyllou K, Smyrnis N, Rizos E. Gut Microbiome: A Brief Review on Its Role in Schizophrenia and First Episode of Psychosis. Microorganisms. 2022; 10(6):1121. https://doi.org/10.3390/microorganisms10061121
Chicago/Turabian StyleTsamakis, Konstantinos, Sofia Galinaki, Evangelos Alevyzakis, Ioannis Hortis, Dimitrios Tsiptsios, Evangelia Kollintza, Stylianos Kympouropoulos, Konstantinos Triantafyllou, Nikolaos Smyrnis, and Emmanouil Rizos. 2022. "Gut Microbiome: A Brief Review on Its Role in Schizophrenia and First Episode of Psychosis" Microorganisms 10, no. 6: 1121. https://doi.org/10.3390/microorganisms10061121
APA StyleTsamakis, K., Galinaki, S., Alevyzakis, E., Hortis, I., Tsiptsios, D., Kollintza, E., Kympouropoulos, S., Triantafyllou, K., Smyrnis, N., & Rizos, E. (2022). Gut Microbiome: A Brief Review on Its Role in Schizophrenia and First Episode of Psychosis. Microorganisms, 10(6), 1121. https://doi.org/10.3390/microorganisms10061121