
Streptomycetaceae and Promicromonosporaceae: Two Actinomycetes Families from Moroccan Oat Soils Enhancing Solubilization of Natural Phosphate

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Material and Methods
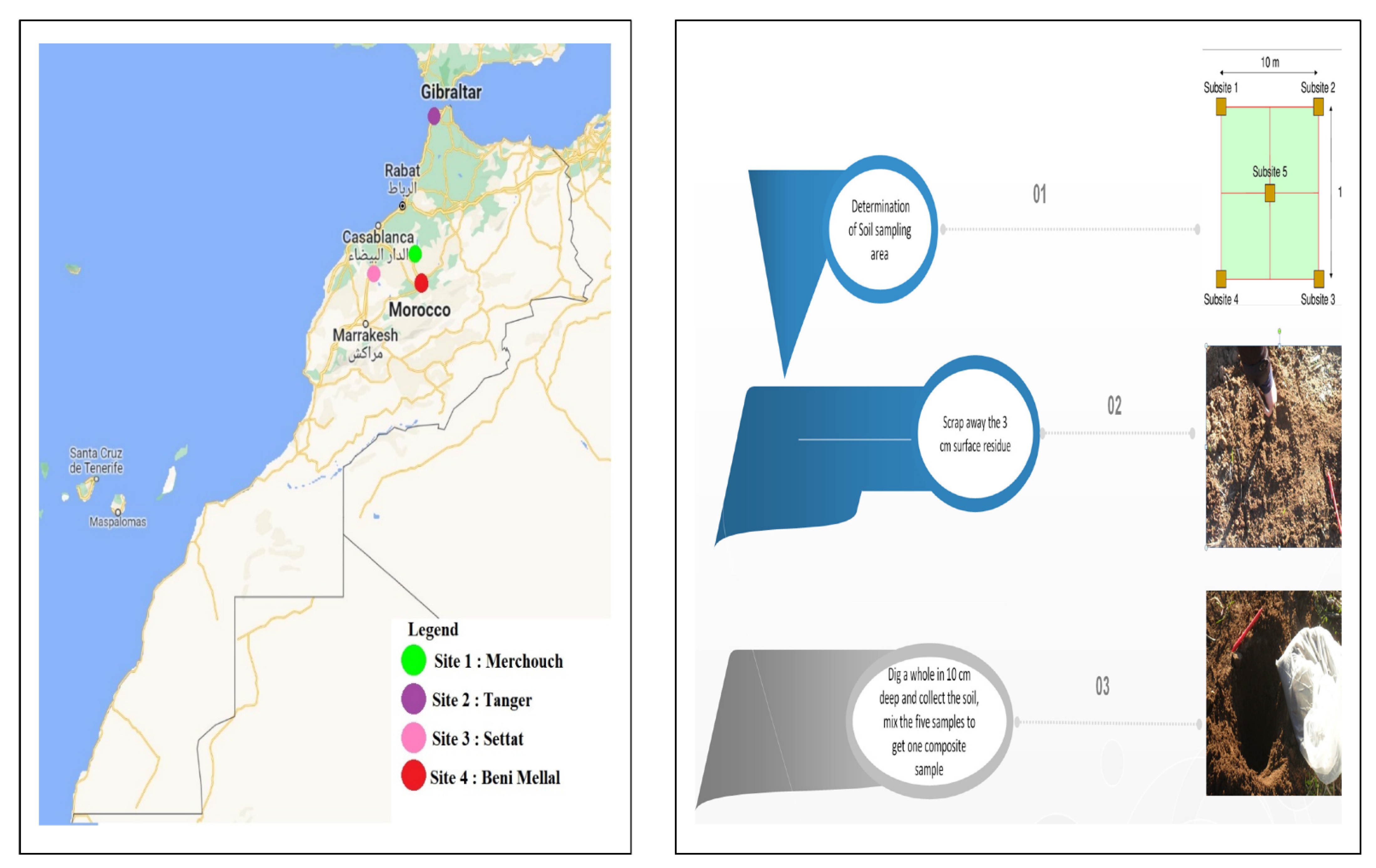
2.1. Study Area and Soil Sampling
2.2. Physicochemical Analysis
2.3. Isolation of Total Flora and Actinomycetes from Oat Soils
2.4. Screening for Actinomycetes Able to Use Rock Phosphate (RP) and Tricalcium Phosphate (TCP) as Unique P Source
2.5. Quantitative Estimation of the Amount Soluble P Released in the Growth Medium by the Selected Actinomycete Strains
2.6. Siderophore Production Test: CAS Agar Plate Technique
2.7. Morphological, Physiological, and Chemotaxonomic Characterization of the Selected Strains
2.8. Amplification and Sequencing of the 16S rDNA of the Selected Strains
2.9. Statistical Analysis
3. Results
3.1. Soil Properties and Distribution of Actinomycetes
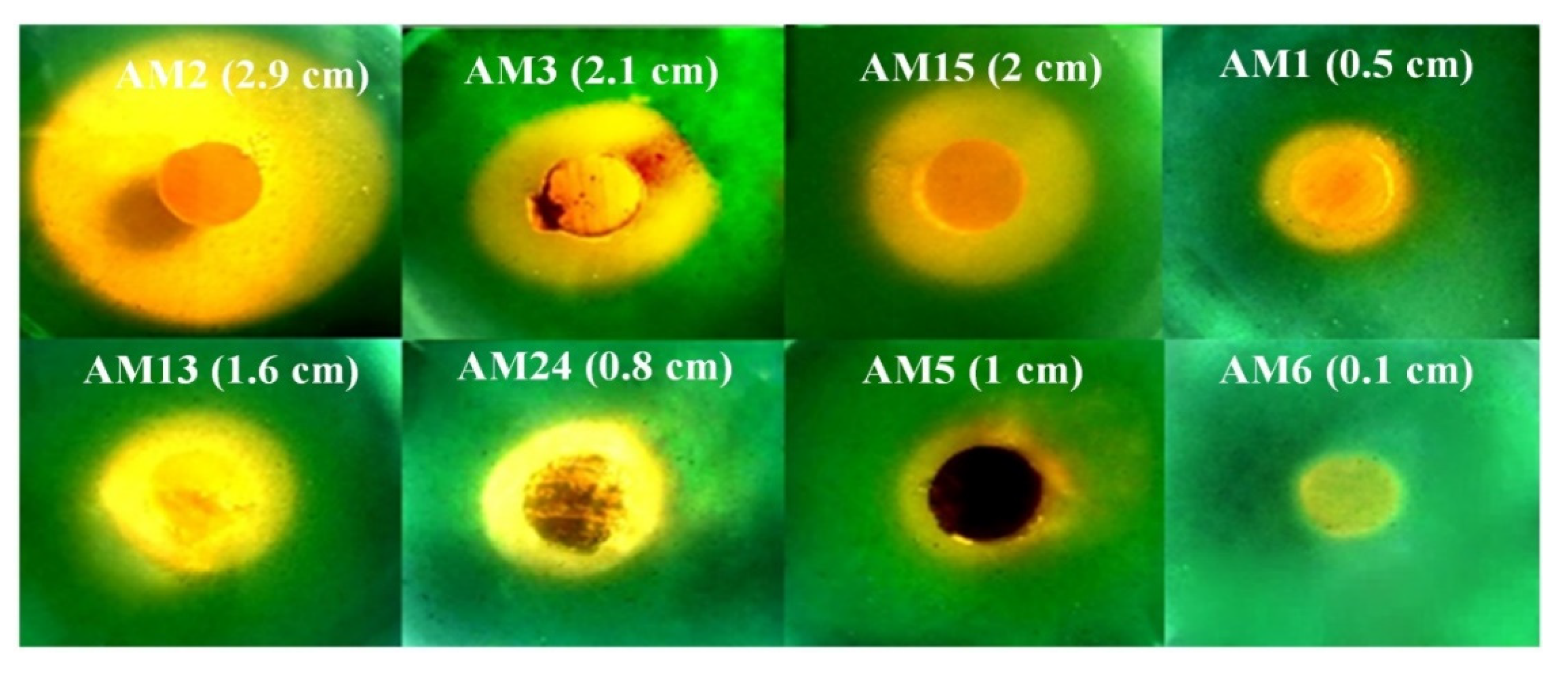
3.2. Isolation of Actinomycetes from Four Different Sites
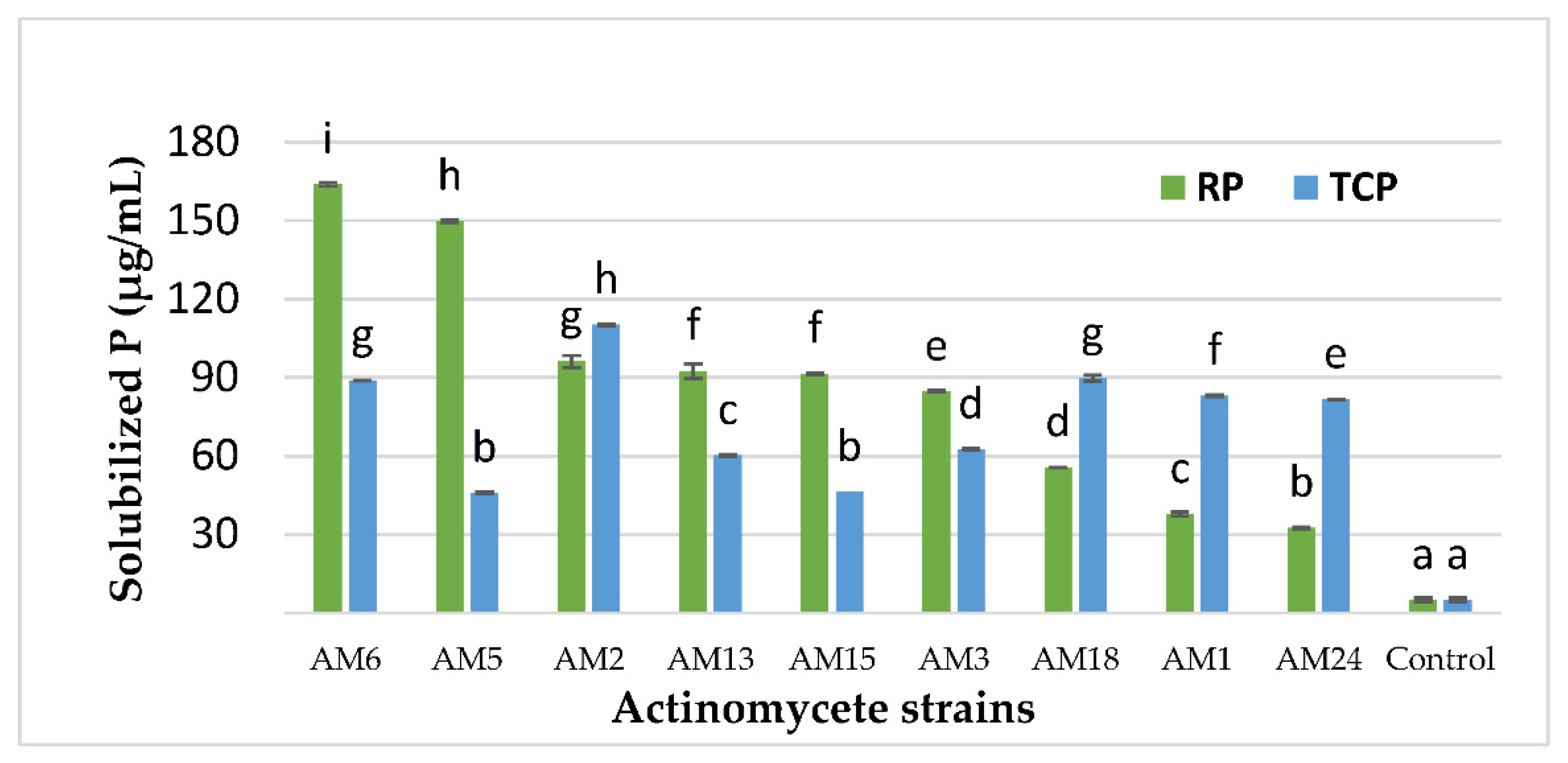
3.3. Quantitative Estimation of the P Concentration Solubilized by the Selected Actinomycetes
3.4. Investigation of the P Biosolubilization Mechanism
3.5. Biochemical and Taxonomical Characterization of the Selected Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). The Future of Food and Agriculture: Trends and Challenges; FAO: Rome, Italy, 2017; Volume 296, pp. 1–180. [Google Scholar]
- Kochhar, S.L. Economic Botany: A Comprehensive Study, 5th ed.; Cambridge University Press: Cambridge, UK, 2016; ISBN 9781316638224. [Google Scholar]
- Al Faïz, C.; Mohamed, C.; Allagui, M.B.; Sbeita, A. Fodder Oats in the Maghreb. In Fodder Oats: A World Overview; FAO: Rome, Italy, 2004; pp. 53–69. [Google Scholar]
- Schlörmann, W.; Zetzmann, S.; Wiege, B.; Haase, N.U.; Greiling, A.; Lorkowski, S. Impact of Different Roasting Conditions on Sensory Properties and Health-Related Compounds of Oat Products. Food Chem. 2020, 307, 125548. [Google Scholar] [CrossRef] [PubMed]
- Manzali, R.; Antari, A.E.; Douaik, A.; Taghouti, M.; Benchekroun, M.; Bouksaim, M.; Saidi, N. Profiling of Nutritional and Health-Related Compounds in Developed Hexaploid Oat Lines Derivative of Interspecific Crosses. Int. J. Celiac Dis. 2017, 5, 72–76. [Google Scholar] [CrossRef][Green Version]
- Bączeka, N.; Jarmułowicz, A.; Wronkowska, M.; Haros, C.M. Assessment of the Glycaemic index, Content of Bioactive Compounds, and Their In Vitro Bioaccessibility in Oat-buckwheat Breads. Food Chem. 2020, 330, 127199. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; De, S.; Belkheir, A. Avena sativa (Oat), A Potential Neutraceutical and Therapeutic Agent: An Overview. Crit. Rev. Food Sci. Nutr. 2013, 53, 126–144. [Google Scholar] [CrossRef]
- Kumar, K. Functional Foods and Lifestyle Diseases EC. EC Nutr. 2017, 11, 216–217. [Google Scholar]
- Nuraini, Y.A.; Handayanto, E.; Syekhfani, S. The Potential of Phosphate Solubilizing Bacteria Isolated From Sugarcane Wastes for Solubilizing Phosphate. J. Degrad. Min. Lands Manag. 2014, 1, 175–182. [Google Scholar] [CrossRef][Green Version]
- Khan, M.M.A.; Haque, E.; Paul, N.C.; Khaleque, M.A.; Al-Garni, S.M.S.; Islam, M.T. Enhancement of Growth and Grain Yield of Rice in Nutrient Deficient Soils by Rice Probiotic Bacteria. Rice Sci. 2017, 24, 264–273. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate Solubilizing Microbes: Sustainable Approach for Managing Phosphorus Deficiency in Agricultural Soils. Springerplus 2013, 2, 587. [Google Scholar] [CrossRef]
- Roychowdury, D.; Paul, M.; Kumar, B.S. Isolation Identification and Partial Characterization of Nitrogen Fixing Bacteria from Soil and Then the Production of Biofertilizer. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 808–815. [Google Scholar]
- Sagervanshi, A.; Priyanka, K.; Anju, N.; Ashwani, K. Isolation and Characterization of Phosphate Solubilizing Bacteria from Agriculture Soil. Int. J. Life Sci. Pharma Res. 2012, 23, 256–266. [Google Scholar]
- Azaroual, S.E.; Hazzoumi, Z.; El Mernissi, N.; Aasfar, A.; Kadmiri, M.I.; Bouizgarne, B. Role of Inorganic Phosphate Solubilizing Bacilli Isolated from Moroccan Phosphate Rock Mine and Rhizosphere Soils in Wheat (Triticum aestivum L.) Phosphorus Uptake. Curr. Microbiol. 2020, 77, 2391–2404. [Google Scholar] [CrossRef] [PubMed]
- Jena, J.; Palai, J.B.; Dash, G.K. Nutrient Managment for Sustainable Oat Production—A Review. Forage Res. 2019, 45, 165–172. [Google Scholar]
- Goldstein, A.H.; Rogers, R.D. Biomediated Continuous Release Phosphate Fertilizer. U.S. Patent 5912398, 1 January 1999. [Google Scholar]
- Macías, F.A.; Marín, D.; Oliveros-bastidas, A.; Varela, R.M.; Simonet, A.M.; Molinillo, J.M.G. Allelopathy as a New Strategy for Sustainable Ecosystems Development. Biol. Sci. Space 2003, 17, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Aallam, Y.; Dhiba, D.; Lemriss, S.; Souiri, A.; Karray, F.; El Rasafi, T.; Saïdi, N.; Haddioui, A.; El Kabbaj, S.; Virolle, M.J.; et al. Isolation and Characterization of Phosphate Solubilizing Streptomyces sp. Endemic from Sugar Beet Fields of the Beni-Mellal Region in Morocco. Microorganisms 2021, 9, 914. [Google Scholar] [CrossRef]
- Aallam, Y.; El Maliki, B.; Dhiba, D.; Lemriss, S.; Souiri, A.; Haddioui, A.; Tarkka, M.; Hamdali, H. Multiple Potential Plant Growth Promotion Activities of Endemic Streptomyces spp. from Moroccan Sugar Beet Fields with Their Inhibitory Activities against Fusarium spp. Microorganisms 2021, 9, 1429. [Google Scholar] [CrossRef]
- Aallam, Y.; Dhiba, D.; El Rasafi, T.; Lemriss, S.; Haddioui, A.; Tarkka, M.; Hamdali, H. Growth Promotion and Protection Against Root Rot of Sugar Beet (Beta vulgaris L.) by Two Rock Phosphate and Potassium Solubilizing Streptomyces spp. under Greenhouse Conditions. Plant Soil 2022, 472, 407–420. [Google Scholar] [CrossRef]
- Bargaz, A.; Lyamlouli, K.; Chtouki, M.; Zeroual, Y.; Dhiba, D. Soil Microbial Resources for Improving Fertilizers Efficiency in an Integrated Plant Nutrient Management System. Front. Microbiol. 2018, 9, 1606. [Google Scholar] [CrossRef]
- Hamdali, H.; Bouizgarne, B.; Hafidi, M.; Lebrihi, A.; Virolle, M.J.; Ouhdouch, Y. Screening for Rock Phosphate Solubilizing Actinomycetes from Moroccan Phosphate Mines. Appl. Soil Ecol. 2008, 38, 12–19. [Google Scholar] [CrossRef]
- Hamdali, H.; Moursalou, K.; Tchangbedji, G.; Ouhdouch, Y.; Hafidi, M. Isolation and Characterization of Rock Phosphate Solubilizing Actinobacteria from a Togolese Phosphate Mine. Afr. J. Biotechnol. 2012, 11, 312–320. [Google Scholar] [CrossRef]
- Hamdali, H.; Lebrihi, A.; Monje, M.C.; Benharref, A.; Hafidi, M.; Ouhdouch, Y.; Virolle, M.J. A Molecule of the Viridomycin Family Originating from a Streptomyces griseus—Related Strain Has the Ability to Solubilize Rock Phosphate and to Inhibit Microbial Growth. Antibiotics 2021, 10, 72. [Google Scholar] [CrossRef]
- Zeroual, Y.; Chadghan, R.; Hakam, A.; Kossir, A. Biotechnology & Biomaterials Biosolubilization of Mineral Insoluble Phosphates by Immobilized Fungi (Aspergillus niger) in Fluidized Bed Bioreactor. J. Biotechnol. Biomater. 2012, S6, 4. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- Khan, A.A.; Jilani, G.; Akhtar, M.S.; Naqvi, S.M.S.; Rasheed, M. Phosphorus Solubilizing Bacteria: Occurrence, Mechanisms and their Role in Crop Production. J. Agric. Biol. Sci. 2009, 1, 48–58. [Google Scholar]
- Farhat, M.B.; Boukhris, I.; Chouayekh, H. Mineral phosphate solubilization by Streptomyces sp. CTM396 involves the Excretion of Gluconic Acid and is Stimulated by Humic Acids. FEMS Microbiol. Lett. 2015, 362, fnv008. [Google Scholar] [CrossRef]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Microbial Pathogenesis Actinomycetes Benefaction Role in Soil and Plant Health. Microb. Pathog. 2017, 111, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Chouyia, F.E.; Romano, I.; Fechtali, T.; Fagnano, M.; Fiorentino, N.; Visconti, D.; Idbella, M.; Ventorino, V.; Pepe, O. P-Solubilizing Streptomyces roseocinereus ms1b15 with Multiple Plant Growth-Promoting Traits Enhance Barley Development and Regulate Rhizosphere Microbial Population. Front. Plant Sci. 2020, 11, 1137. [Google Scholar] [CrossRef]
- Hamdali, H.; Hafidi, M.; Virolle, M.J.; Ouhdouch, Y. Rock Phosphate-Solubilizing Actinomycetes: Screening for Plant Growth-promoting Activities. World J. Microbiol. Biotechnol. 2008, 24, 2565–2575. [Google Scholar] [CrossRef]
- Hamdali, H.; Hafidi, M.; Virolle, M.J.; Ouhdouch, Y. Growth Promotion and Protection against Damping-off of Wheat by Two Rock Phosphate Slubilizing Actinomycetes in a P-deficient Soil Under Greenhouse Conditions. Appl. Soil Ecol. 2008, 40, 510–517. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and Interactions in Plant Growth Promotion. Appl. Microbiol. Biotechnol. 2018, 103, 1179–1188. [Google Scholar] [CrossRef]
- Nandimath, A.P.; Karad, D.D.; Gupta, S.G.; Kharat, A.S. Consortium Inoculum of Five Thermo-tolerant Phosphate Solubilizing Actinomycetes for Multipurpose Biofertilizer Preparation. Iran. J. Microbiol. 2017, 9, 295–304. [Google Scholar]
- Wei, J.; He, L.; Niu, G. Regulation of Antibiotic Biosynthesis in Actinomycetes: Perspectives and challenges. Synth. Syst. Biotechnol. 2018, 3, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Ait Barka, E.; Vatsa, P.; Sanchez, L.; Gaveau-vaillant, N.; Jacquard, C.; Klenk, H.; Clément, C.; Ouhdouch, Y.; Wezel, P.V. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, S.; Shahid, I.; Mehnaz, S.; Malik, K.A. Assessment of Two Carrier Materials for Phosphate Solubilizing Biofertilizers and Their Effect on Growth of Wheat (Triticum aestivum L.). Microbiol. Res. 2017, 205, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Tarkka, M.T.; Grams, T.E.E.; Angay, O.; Kurth, F.; Fleischmann, F.; Ruess, L.; Schädler, M.; Scheu, S.; Schrey, S.D.; Buscot, F.; et al. Ectomycorrhizal Fungus Supports Endogenous Rhythmic Growth and Corresponding Resource Allocation in Oak During Various Below—And Aboveground Biotic Interactions. Sci. Rep. 2021, 11, 23680. [Google Scholar] [CrossRef]
- Yeomans, J.C.; Bremner, J.M. Communications in Soil Science and Plant Analysis A Rapid and Precise Method for Routine Determination of Organic Carbon in Soil. Commun. Soil Sci. Plant Anal. 2013, 19, 1467–1476. [Google Scholar] [CrossRef]
- Olsen, S.R.; Sommers, L.E. Methods of Soil Analysis, Part 2; American Society of Agronomy: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Qin, S.; Miao, Q.; Feng, W.; Wang, Y.; Zhu, X.; Xing, K.; Jiang, J.-H. Biodiversity and Plant Growth Promoting Traits of Culturable Endophytic Actinobacteria Associated with Jatropha curcas L. growing in Panxi dry-hot valley soil. Appl. Soil Ecol. 2015, 93, 47–55. [Google Scholar] [CrossRef]
- Shirling, E.B.; Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal Chemical Assay for the Detection and Determination of Siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acids Techniques in Bacterial Systematics; Wiley: Chichester, UK, 1991; pp. 115–147. [Google Scholar]
- Flandrois, J.P.; Perrière, G.; Gouy, M. leBIBIQBPP: A Set of Databases and a Webtool for Automatic Phylogenetic Analysis of Prokaryotic Sequences. BMC Bioinform. 2015, 16, 251. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Phylogenies And The Comparative Method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Kimura, M.A. Simple Method for Estimating Evolutionary Rates of Base Substitutions Through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis Across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Soumare, A.; Boubekri, K.; Lyamlouli, K.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L. From Isolation of Phosphate Solubilizing Microbes to Their Formulation and Use as Biofertilizers: Status and Needs. Front. Bioeng. Biotechnol. 2020, 7, 425. [Google Scholar] [CrossRef] [PubMed]
- Suleman, M.; Yasmin, S.; Rasul, M.; Yahya, M.; Atta, B.M.; Mirza, M.S. Phosphate Solubilizing Bacteria With Glucose Dehydrogenase Gene for Phosphorus Uptake and Beneficial Efects on Wheat. PLoS ONE 2018, 13, e0204408. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.; Naeem, M.A.; Shahid, M. Inoculation of pqqE Gene Inhabiting Pantoea and Pseudomonas strains Improves the Growth and Grain Yield of Wheat With a Reduced Amount of Chemical Fertilizer. J. Appl. Microbiol. 2020, 129, 575–589. [Google Scholar] [CrossRef]
- Alam, M.; Dharni, S.; Abdul-Khaliq; Srivastava, S.K.; Samad, A.; Gupta, M.K. A Promising Strain of Streptomyces sp. with Agricultural Traits for Growth Promotion and Disease Management. Indian J. Exp. Biol. 2012, 50, 559–568. [Google Scholar]
- Xue, P.; Carrillo, Y.; Pino, V.; Minasny, B.; Mcbratney, A.B. Soil Properties Drive Microbial Community Structure in a Large Scale Transect in South Eastern. Sci. Rep. 2018, 8, 11725. [Google Scholar] [CrossRef]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. Soil Biology & Biochemistry The Influence of Soil Properties on the Structure of Bacterial and Fungal Communities Across Land-use Types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Mccarthy, A.J.; Williams, S.T. Actinomycetes as Agents of Biodegradation in the Environment—A review. Gene 1992, 115, 189–192. [Google Scholar] [CrossRef]
- Seshachala, U.; Tallapragada, P. Phosphate Solubilizers from the Rhizosphere of Piper nigrum L. in karnataka, India. Chil. J. Agric. Res. 2012, 72, 397–403. [Google Scholar] [CrossRef]
- Tian, J.; Ge, F.; Zhang, D.; Deng, S.; Liu, X. Roles of Phosphate Solubilizing Microorganisms from. Biology 2021, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Feng, K.; Pei, Z.; Meng, F.; Sun, J. Multivariate Analysis Combined with GIS to Source Identification of Heavy Metals in Soils Around an Abandoned Industrial Area, Eastern China. Ecotoxicology 2016, 25, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Mardad, I.; Serrano, A.; Soukri, A. Solubilization of Inorganic Phosphate and Production of Organic Acids by Bacteria Isolated from a Moroccan Mineral Phosphate Deposit. Afr. J. Microbiol. Res. 2013, 7, 626–635. [Google Scholar] [CrossRef]
- Puentel, M.E.; Li, C.Y.; Bashan, Y. Microbial Populations and Activities in the Rhizoplane of Rock-Weathering Desert Plants. II. Growth Promotion of Cactus Seedlings. Plant Soil 2004, 6, 643–650. [Google Scholar]
- Rodríguez, H.; Fraga, R. Phosphate Solubilizing Bacteria and their Role in Plant Growth Promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, G.; Wei, Y.; Dong, Y.; Hou, L. Isolation and Screening of Multifunctional Phosphate Solubilizing Bacteria and its Growth-Promoting Effect on Chinese Fir Seedlings. Sci. Rep. 2021, 11, 9081. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Zaidi, A.; Ahmad, E. Mechanism of Phosphate Solubilization and Physiological Functions of Phosphate-Solubilizing Microorganisms. In Phosphate Solubilizing Microorganisms; Khan, M., Zaidi, A., Musarrat, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Gong, Y.; Bai, J.; Yang, H.; Zhang, W.; Xiong, Y.; Ding, P.; Qin, S. Phylogenetic Diversity and Investigation of Plant Growth-Promoting Traits of Actinobacteria in Coastal Salt Marsh Plant Rhizospheres from Jiangsu, China. Syst. Appl. Microbiol. 2018, 41, 516–527. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | ||||||
| RP | TCP | RP | TCP | RP | TCP | RP | TCP | RP | TCP | |
| AM2 | 6.88 ± 0.01 | 6.55 ± 0.21 | 6.93 ± 0.04 | 6.75 ± 0.21 | 7.09 ± 0.01 | 7.05 ± 0.06 | 7.35 ± 0.07 | 7.24 ± 0.01 | 7.47 ± 0.04 | 7.60 ± 0.28 |
| AM5 | 6.80 ± 0.01 | 6.80 ± 0.01 | 6.86 ± 0.01 | 6.93 ± 0.13 | 6.80 ± 0.00 | 7.16 ± 0.04 | 7.12 ± 0.04 | 7.5 ± 0.01 | 7.62 ± 0.04 | 7.55 ± 0.07 |
| AM6 | 6.815 ± 0.01 | 6.81 ± 0.01 | 7.19 ± 0.01 | 7.22 ± 0.02 | 7.10 ± 0.13 | 7.25 ± 0.02 | 6.90 ± 0.00 | 7.46 ± 0.06 | 7.23 ± 0.01 | 7.52 ± 0.02 |
| AM13 | 6.65 ± 0.07 | 6.82 ± 0.02 | 6.89 ± 0.01 | 6.90 ± 0.17 | 6.91 ± 0.01 | 7.06 ± 0.05 | 7.07 ± 0.01 | 7.10 ± 0.02 | 7.71 ± 0.01 | 7.59 ± 0.14 |
| AM15 | 6.03 ± 0.02 | 6.82 ± 0.04 | 6.85 ± 0.07 | 6.80 ± 0.00 | 7.43 ± 0.04 | 7.46 ± 0.06 | 7.43 ± 0.11 | 7.45 ± 0.06 | 7.49 ± 0.04 | 7.5 ± 0.06 |
| AM1 | 6.8 ± 0.01 | 6.75 ± 0.07 | 6.88 ± 0.02 | 6.8 ± 0.01 | 7.1 ± 0.02 | 7.23 ± 0.14 | 7.3 ± 0.01 | 7.7 ± 0.03 | 7.35 ± 0.01 | 7.81 ± 0.01 |
| AM3 | 6 ± 0.01 | 6.9 ± 0.01 | 6.5 ± 0.05 | 7.08 ± 0.01 | 6.59 ± 0.00 | 7.43 ± 0.11 | 6.8 ± 0.02 | 7.6 ± 0.01 | 6.86 ± 0.14 | 7.8 ± 0.01 |
| AM24 | 7.01 ± 0.01 | 6.5 ± 0.05 | 7.14 ± 0.01 | 6.67 ± 0.02 | 7.28 ± 0.13 | 7.12 ± 0.00 | 7.45 ± 0.04 | 7.32 ± 0.01 | 7.7 ± 0.01 | 7.56 ± 0.017 |
| Strains | AM2 | AM5 | AM6 | AM13 | AM15 | AM1 | AM3 | AM24 |
|---|---|---|---|---|---|---|---|---|
| Origin | Site 3 | Site 2 | Site 1 | Site 4 | Site 1 | Site 3 | Site 4 | |
| Aerial spore mass | Yellow | White | Gray | White | Pink | Yellow | White | Brown |
| Soluble pigment | - | Green | - | Beige | - | - | - | - |
| Colony reverse | Yellow | Brown | Brown | Beige | White | White | White | Brown |
| C. source utilization | ||||||||
| Glucose | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ |
| Fructose | +++ | +++ | +++ | +++ | ++ | + | ++ | + |
| Maltose | +++ | +++ | +++ | +++ | ++ | ++ | + | + |
| Lactose | +++ | +++ | +++ | +++ | ++ | + | ++ | + |
| Amidon | +++ | +++ | +++ | +++ | ++ | + | + | + |
| Mannitol | +++ | +++ | +++ | +++ | ++ | + | + | ++ |
| Sorbitol | +++ | +++ | +++ | +++ | + | ++ | ++ | + |
| Citrate | +++ | +++ | +++ | +++ | + | + | + | + |
| Salinity resistance % (maximum of resistance to NaCl on g/L) | 9.5 | 9.5 | 9.5 | 9.5 | 7.5 | 7.5 | 2.5 | 9.5 |
| pH resistance | 5–13 | 5–7 | 5–10 | 5–13 | 5–13 | 5–13 | 7–10 | 5–13 |
| Strains | 16S rRNA Identification | Accession Number |
|---|---|---|
| AM2 | Streptomyces griseoflavus | MW854007 |
| AM5 | Streptomycessampsonii | MW854325 |
| AM6 | Streptomyces anulatus | MW854030 |
| AM15 | Streptomyces aegyptia | MT505706 |
| AM13 | Promicromonospora alba | MT505707 |
| AM1 | Streptomyces exfoliates | OK501973 |
| AM3 | Streptomyces pratensis | OK501976 |
| AM24 | Streptomyces griseoincarnatus | OK501975 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bousselham, M.; Lemriss, S.; Dhiba, D.; Aallam, Y.; Souiri, A.; Abbas, Y.; Saïdi, N.; Boukcim, H.; Hamdali, H. Streptomycetaceae and Promicromonosporaceae: Two Actinomycetes Families from Moroccan Oat Soils Enhancing Solubilization of Natural Phosphate. Microorganisms 2022, 10, 1116. https://doi.org/10.3390/microorganisms10061116
Bousselham M, Lemriss S, Dhiba D, Aallam Y, Souiri A, Abbas Y, Saïdi N, Boukcim H, Hamdali H. Streptomycetaceae and Promicromonosporaceae: Two Actinomycetes Families from Moroccan Oat Soils Enhancing Solubilization of Natural Phosphate. Microorganisms. 2022; 10(6):1116. https://doi.org/10.3390/microorganisms10061116
Chicago/Turabian StyleBousselham, Meriam, Sanaa Lemriss, Driss Dhiba, Yassine Aallam, Amal Souiri, Younes Abbas, Nezha Saïdi, Hassan Boukcim, and Hanane Hamdali. 2022. "Streptomycetaceae and Promicromonosporaceae: Two Actinomycetes Families from Moroccan Oat Soils Enhancing Solubilization of Natural Phosphate" Microorganisms 10, no. 6: 1116. https://doi.org/10.3390/microorganisms10061116
APA StyleBousselham, M., Lemriss, S., Dhiba, D., Aallam, Y., Souiri, A., Abbas, Y., Saïdi, N., Boukcim, H., & Hamdali, H. (2022). Streptomycetaceae and Promicromonosporaceae: Two Actinomycetes Families from Moroccan Oat Soils Enhancing Solubilization of Natural Phosphate. Microorganisms, 10(6), 1116. https://doi.org/10.3390/microorganisms10061116