
Microbial Consortium Associated with Crustacean Shells Composting




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site Description and Sample Collection
2.2. DNA Extraction and Sequencing
2.3. Sequence Processing
2.4. Bioinformatics and Statistical Analysis
3. Results
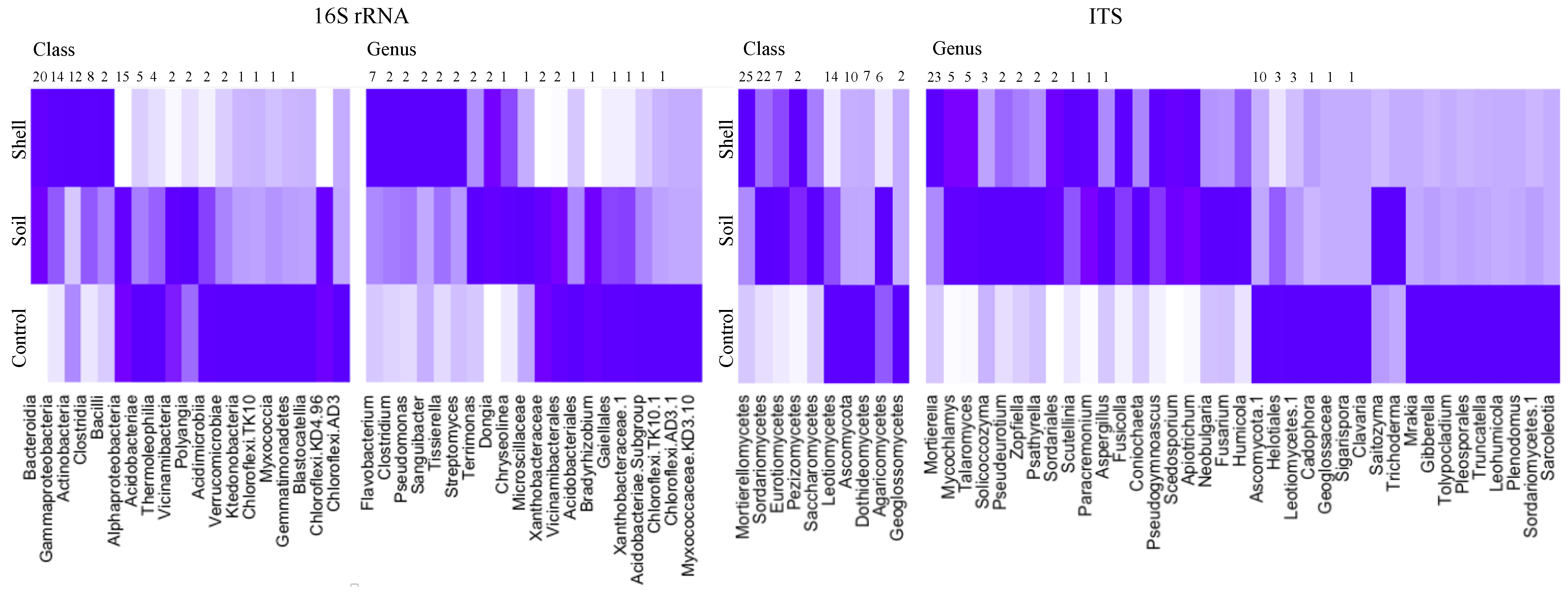
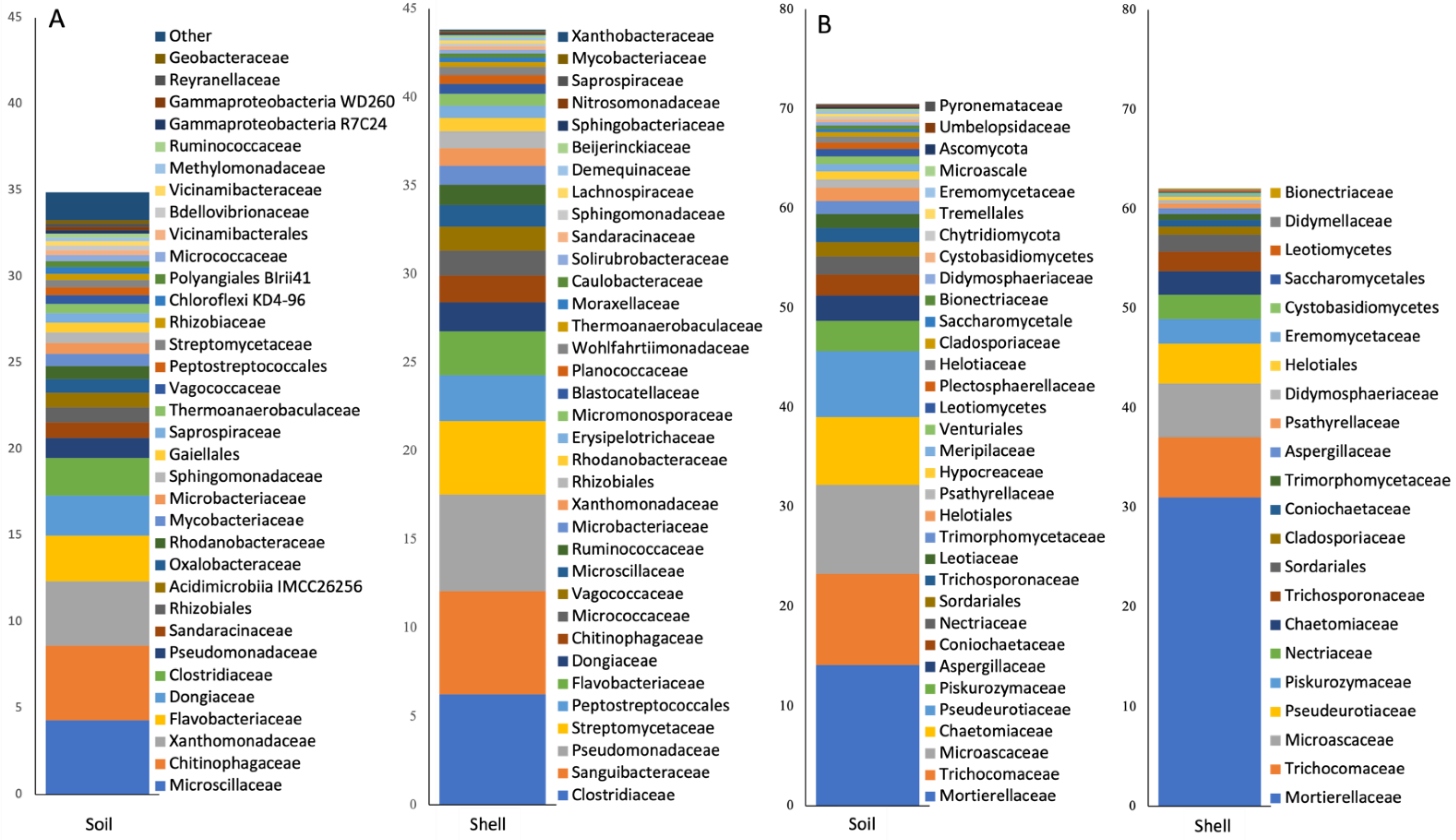
3.1. Taxonomic Compositing
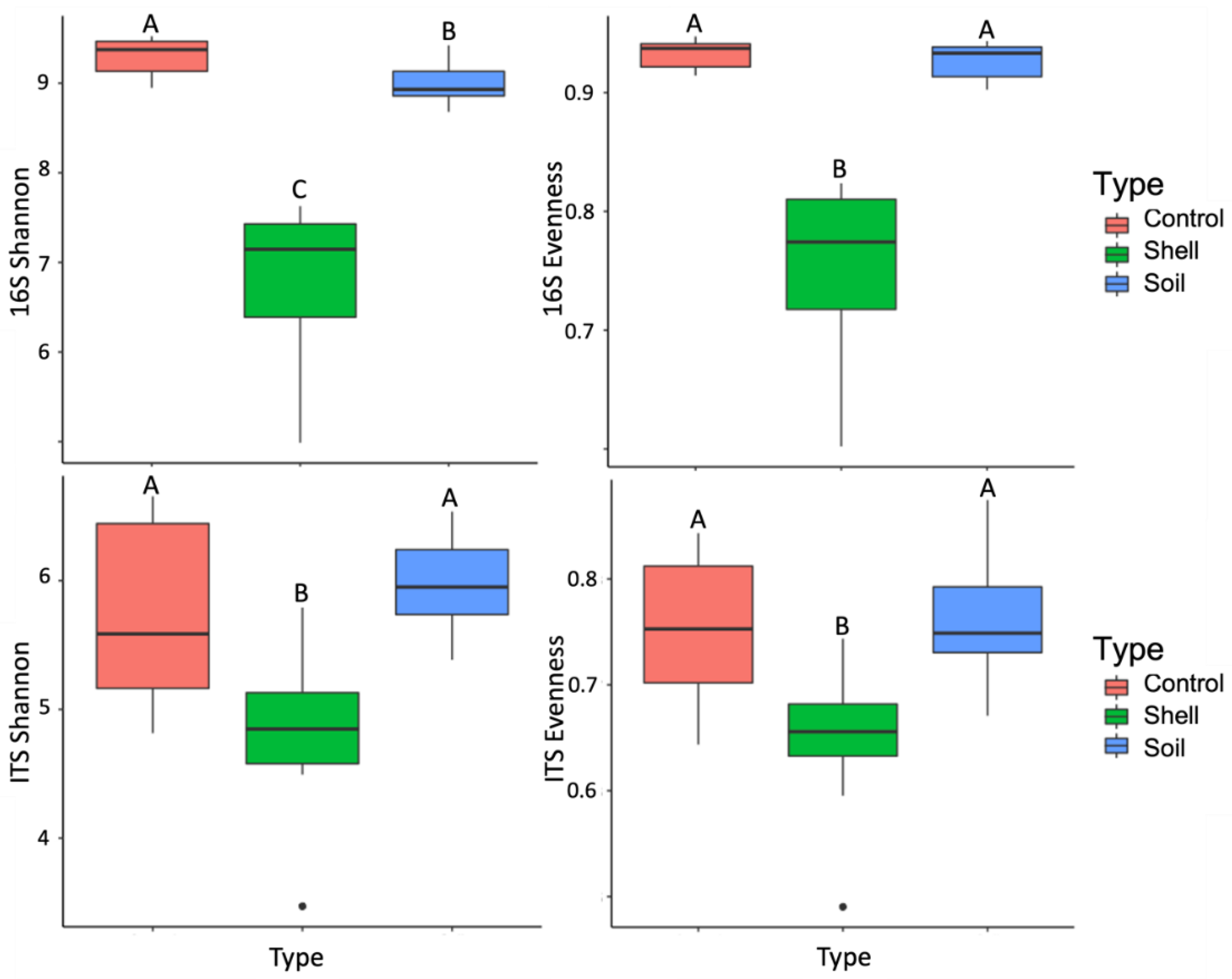
3.2. The Effect of Shell Proximity on the Diversity of Microbial Communities
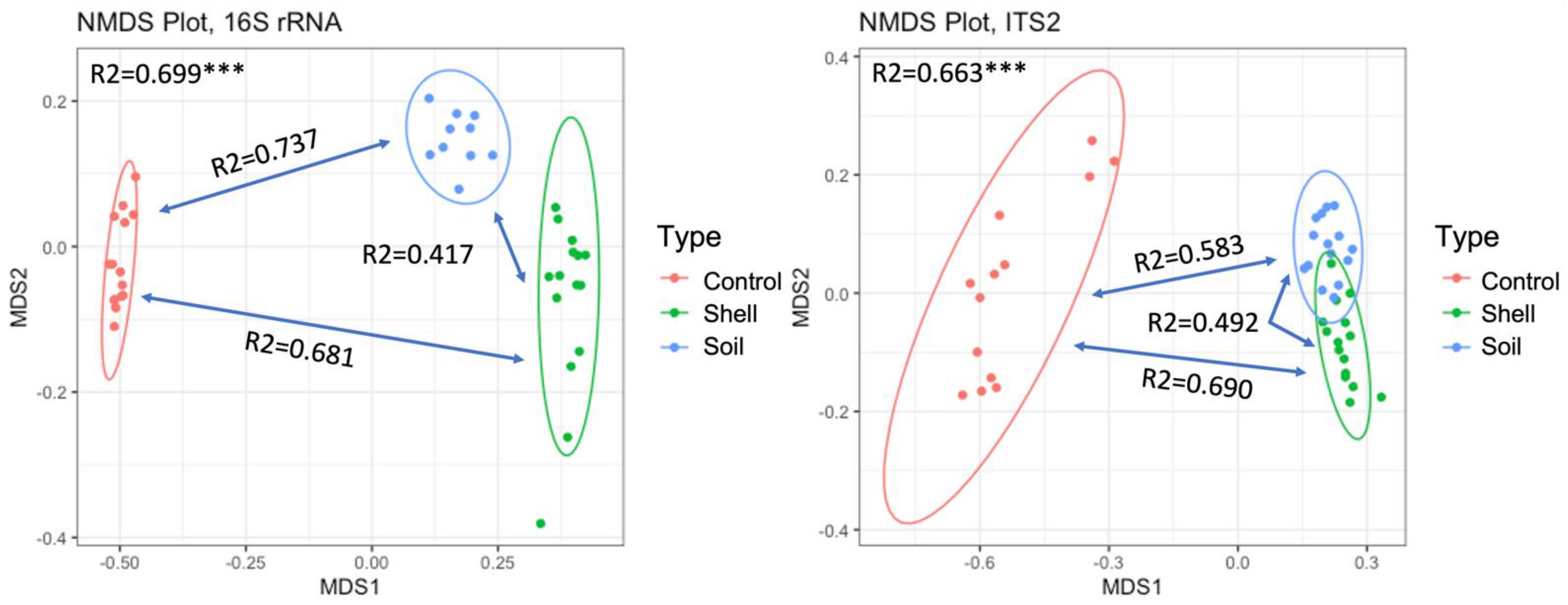
3.3. The Effect of Shell Proximity on the Composition of Microbial Communities
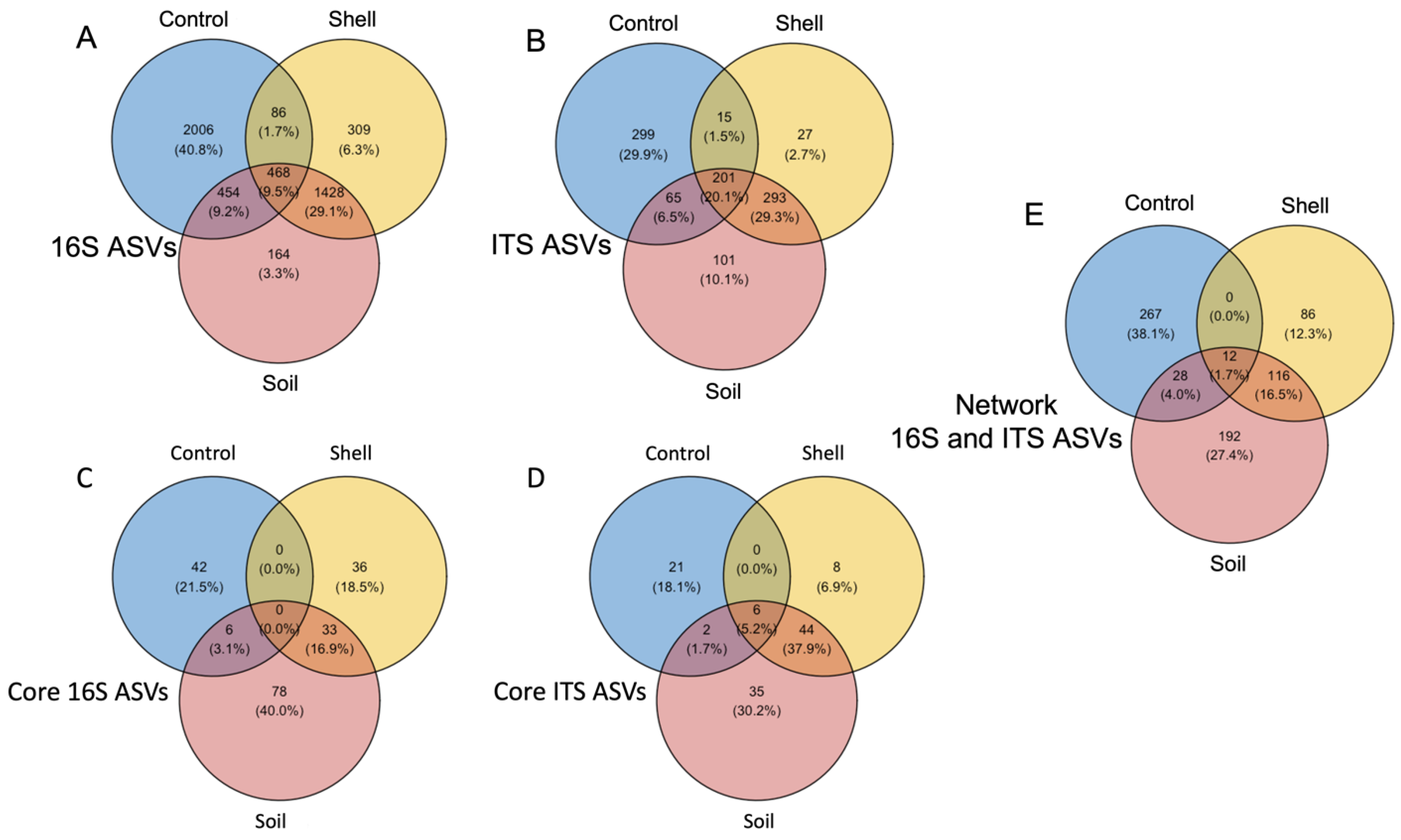
3.4. Core Microbiomes
3.5. Microbiome Cooperation
3.6. 16S and ITS2 ASVs with Strong Association with Shell Environment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, V.P.; Marques, N.S.S.; Maia, P.; Lima, M.A.B.; Franco, L.O.; Campos-Takaki, G.M. Seafood Waste as Attractive Source of Chitin and Chitosan Production and Their Applications. Int. J. Mol. Sci. 2020, 21, 4290. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Chen, X. Sustainability: Don’t waste seafood waste. Nature 2015, 524, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Barber, A.R.; Corbin, K.; Zhang, W. Lobster processing by-products as valuable bioresource of marine functional ingredients, nutraceuticals, and pharmaceuticals. Bioresour. Bioprocess. 2017, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Le, B.; Yang, S.H. Microbial chitinases: Properties, current state and biotechnological applications. World J. Microbiol. Biotechnol. 2019, 35, 144. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, V.; Bhatt, A.K.; Sharma, V.; Rathour, R.K.; Rana, N.; Bhatia, R.K.; Varjani, S.; Kumar, M.; Magdouli, S.; Yung, H.; et al. Advances in glucosamine production from waste biomass and microbial fermentation technology and its applications. Biomass Convers. Biorefin. 2021, 1–23. [Google Scholar] [CrossRef]
- Radwan, M.A.; Farrag, S.A.A.; Abu-Elamayem, M.M.; Ahmed, N.S. Extraction, characterization, and nematicidal activity of chitin and chitosan derived from shrimp shell wastes. Biol. Fertil. Soils 2021, 48, 463–468. [Google Scholar] [CrossRef]
- Ilangumaran, G.; Stratton, G.; Ravichandran, S.; Shukla, P.S.; Potin, P.; Asiedu, S.; Prithiviraj, B. Microbial Degradation of Lobster Shells to Extract Chitin Derivatives for Plant Disease Management. Front. Microbiol. 2017, 8, 781. [Google Scholar] [CrossRef]
- Andreo-Jimenez, B.; Schilder, M.T.; Nijhuis, E.H.; Te Beest, D.E.; Bloem, J.; Visser, J.H.M.; van Os, G.; Brolsma, K.; de Boer, W.; Postma, J. Chitin- and Keratin-Rich Soil Amendments Suppress Rhizoctonia solani Disease via Changes to the Soil Microbial Community. Appl. Environ. Microbiol. 2021, 87, e00318-21. [Google Scholar] [CrossRef]
- Randall, T.E.; Fernandez-Bayo, J.D.; Harrold, D.R.; Achmon, Y.; Hestmark, K.V.; Gordon, T.R.; Stapleton, J.J.; Simmons, C.W.; VanderGheynst, J.S. Changes of Fusarium oxysporum f.sp. lactucae levels and soil microbial community during soil biosolarization using chitin as soil amendment. PLoS ONE 2020, 15, e0232662. [Google Scholar] [CrossRef]
- Cretoiu, M.S.; Korthals, G.W.; Visser, J.H.; van Elsas, J.D. Chitin amendment increases soil suppressiveness toward plant pathogens and modulates the actinobacterial and oxalobacteraceal communities in an experimental agricultural field. Appl. Environ. Microbiol. 2013, 79, 5291–5301. [Google Scholar] [CrossRef] [Green Version]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.Á.; Rodríguez, A.T.; Alfonso, L.; Peniche, C. Chitin and its derivatives as biopolymers with potential agricultural applications. Biotecnol. Apl. 2010, 27, 270–276. [Google Scholar]
- Debode, J.; De Tender, C.; Soltaninejad, S.; Van Malderghem, C.; Haegeman, A.; Van der Linden, I.; Cottyn, B.; Heyndrickx, M.; Maes, M. Chitin Mixed in Potting Soil Alters Lettuce Growth, the Survival of Zoonotic Bacteria on the Leaves and Associated Rhizosphere Microbiology. Front. Microbiol. 2016, 7, 565. [Google Scholar] [CrossRef] [PubMed]
- Fabritius, H.O.; Sachs, C.; Triguero, P.R.; Roobe, D. Influence of structural principles on the mechanics of a biological fiber-based composite material with hierarchical organization: The exoskeleton of the lobster Homarus americanus. Adv. Mater. 2009, 21, 391–400. [Google Scholar] [CrossRef]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Castro, R.; Guerrero-Legarreta, I.; Borquez, R. Chitin extraction from Allopetrolisthes punctatus crab using lactic fermentation. Biotechnol. Rep. 2018, 20, e00287. [Google Scholar] [CrossRef]
- Gong, X.; Tian, W.; Bai, J.; Qiao, K.; Zhao, J.; Wang, L. Highly efficient deproteinization with an ammonifying bacteria Lysinibacillus fusiformis isolated from brewery spent diatomite. J. Biosci. Bioeng. 2019, 127, 326–332. [Google Scholar] [CrossRef]
- Abirami, S.; Nagarajan, D. Extraction of Chitin from Shrimp Shell Wastes by Using Bacillus licheniformis and Lactobacillus plantarum. Inter. J. Recent Res. Asp. 2018, 307–315. [Google Scholar]
- Beier, S.; Bertilsson, S. Bacterial chitin degradation-mechanisms and ecophysiological strategies. Front. Microbiol 2013, 4, 149. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, Y.; Ichino, T.; Saito, A. Transition of the Bacterial Community and Culturable Chitinolytic Bacteria in Chitin-treated Upland Soil: From Streptomyces to Methionine-auxotrophic Lysobacter and Other Genera. Microbes Environ. 2020, 35, ME19070. [Google Scholar] [CrossRef] [Green Version]
- Jacquiod, S.; Franqueville, L.; Cecillon, S.; Vogel, T.M.; Simonet, P. Soil bacterial community shifts after chitin enrichment: An integrative metagenomic approach. PLoS ONE 2013, 8, e79699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieczorek, A.S.; Schmidt, O.; Chatzinotas, A.; von Bergen, M.; Gorissen, A.; Kolb, S. Ecological Functions of Agricultural Soil Bacteria and Microeukaryotes in Chitin Degradation: A Case Study. Front. Microbiol. 2019, 10, 1293. [Google Scholar] [CrossRef] [PubMed]
- Baty, A.M., 3rd; Eastburn, C.C.; Diwu, Z.; Techkarnjanaruk, S.; Goodman, A.E.; Geesey, G.G. Differentiation of chitinase-active and non-chitinase-active subpopulations of a marine bacterium during chitin degradation. Appl. Environ. Microbiol. 2000, 66, 3566–3573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baty, A.M., 3rd; Eastburn, C.C.; Techkarnjanaruk, S.; Goodman, A.E.; Geesey, G.G. Spatial and temporal variations in chitinolytic gene expression and bacterial biomass production during chitin degradation. Appl. Environ. Microbiol. 2000, 66, 3574–3585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klassen, G.; Locke, A. A biological synopsis of the European green crab, Carcinus maenas. Can. Manuscr. Rep. Fish. Aquat. Sci. 2007, 2818, 1–82. [Google Scholar] [CrossRef]
- Skonberg, D.I.; Perkins, B.L. Nutrient composition of green crab (Carcinus maenus) leg meat and claw meat. Food Chem. 2002, 77, 401–404. [Google Scholar] [CrossRef]
- Hirano, S. Chitin biotechnology applications. Biotechnol. Ann. Rev. 1996, 2, 237–258. [Google Scholar] [CrossRef]
- Maksimov, I.V.; Abizgil’dina, R.R.; Pusenkova, L.I. Plant growth promoting rhizobacteria as alternative to chemical crop protectors from pathogens (review). Appl. Biochem. Microbiol. 2011, 47, 333–345. [Google Scholar] [CrossRef]
- Comeau, A.M.; Vincent, W.F.; Bernier, L.; Lovejoy, C. Novel chytrid lineages dominate fungal sequences in diverse marine and freshwater habitats. Sci. Rep. 2016, 6, 30120. [Google Scholar] [CrossRef]
- Yurgel, S.N.; Douglas, G.M.; Comeau, A.M.; Mammoliti, M.; Dusault, A.; Percival, D.; Langille, M.G. Variation in Bacterial and Eukaryotic Communities Associated with Natural and Managed Wild Blueberry Habitats. Phytobioms J. 2017, 1, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Comeau, A.M.; Douglas, G.M.; Langille, M.G. Microbiome Helper: A Custom and Streamlined Workflow for Microbiome Research. mSystems 2017, 2, e00127-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Amir, A.; McDonald, D.; Navas-Molina, J.A.; Kopylova, E.; Morton, J.T.; Zech Xu, Z.; Kightley, E.P.; Thompson, L.R.; Hyde, E.R.; Gonzalez, A.; et al. Deblur Rapidly Resolves Single-Nucleotide Community Sequence Patterns. mSystems 2017, 2, e00191-16. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.R.; Shin, J.; Guevarra, R.; Lee, J.H.; Kim, D.W.; Seol, K.H.; Lee, J.H.; Kim, H.B.; Isaacson, R. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package; R Package Version 2.4-0; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://CRAN.R-project.org/package=vegan (accessed on 5 March 2022).
- Fernandes, A.D.; Reid, J.N.; Macklaim, J.M.; McMurrough, T.A.; Edgell, D.R.; Gloor, G.B. Unifying the analysis of high-throughput sequencing datasets: Characterizing RNA-seq, 16S rRNA gene sequencing and selective growth experiments by compositional data analysis. Microbiome 2014, 2, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Schwager, E.; Weingart, G.; Bielski, C.; Huttenhower, C. CCREPE: Compositionality Corrected by PErmutation and REnormalization. 2020. Available online: bioconductor.org (accessed on 20 April 2022).
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- De Caceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Swiontek Brzezinska, M.; Jankiewicz, U.; Burkowska, A.; Walczak, M. Chitinolytic microorganisms and their possible application in environmental protection. Curr. Microbiol. 2014, 68, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.L.; Xiong, J.; Xu, T.; Zhang, C.G.; Zhang, R.Y.; Zhang, Q.; Wu, S.; Qiu, G.Z. Purification and characterization of extracellular chitinase from a novel strain Aspergillus fumigatus CS-01. J. Cent. South Univ. Technol. 2009, 16, 552–557. [Google Scholar] [CrossRef]
- Han, S.J.; Park, H.; Lee, S.G.; Lee, H.K.; Yim, J.H. Optimization of cold-active chitinase production from the Antarctic bacterium, Sanguibacter antarcticus KOPRI 21702. Appl. Microbiol. Biotechnol. 2011, 89, 613–621. [Google Scholar] [CrossRef]
- Starke, R.; Morais, D.; Vetrovsky, T.; Lopez Mondejar, R.; Baldrian, P.; Brabcova, V. Feeding on fungi: Genomic and proteomic analysis of the enzymatic machinery of bacteria decomposing fungal biomass. Environ. Microbiol. 2020, 22, 4604–4619. [Google Scholar] [CrossRef]
- Someya, N.; Ikeda, S.; Morohoshi, T.; Noguchi Tsujimoto, M.; Yoshida, T.; Sawada, H.; Ikeda, T.; Tsuchiya, K. Diversity of culturable chitinolytic bacteria from rhizospheres of agronomic plants in Japan. Microbes Environ. 2011, 26, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Larsbrink, J.; Zhu, Y.; Kharade, S.S.; Kwiatkowski, K.J.; Eijsink, V.G.; Koropatkin, N.M.; McBride, M.J.; Pope, P.B. A polysaccharide utilization locus from Flavobacterium johnsoniae enables conversion of recalcitrant chitin. Biotechnol. Biofuels 2016, 9, 260. [Google Scholar] [CrossRef] [Green Version]
- Kharade, S.S.; McBride, M.J. Flavobacterium johnsoniae chitinase ChiA is required for chitin utilization and is secreted by the type IX secretion system. J. Bacteriol. 2014, 196, 961–970. [Google Scholar] [CrossRef] [Green Version]
- Wegner, C.E.; Liesack, W. Microbial community dynamics during the early stages of plant polymer breakdown in paddy soil. Environ. Microbiol. 2016, 18, 2825–2842. [Google Scholar] [CrossRef] [PubMed]
- Harkin, C.; Bruck, W.M.; Lynch, C. Isolation & identification of bacteria for the treatment of brown crab (Cancer pagurus) waste to produce chitinous material. J. Appl. Microbiol. 2015, 118, 954–965. [Google Scholar] [CrossRef] [PubMed]
- Ghorbel-Bellaaj, O.; Hmidet, N.; Jellouli, K.; Younes, I.; Maalej, H.; Hachicha, R.; Nasri, M. Shrimp waste fermentation with Pseudomonas aeruginosa A2: Optimization of chitin extraction conditions through Plackett-Burman and response surface methodology approaches. Int. J. Biol. Macromol. 2011, 48, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; He, R.; Zhou, X.; Gu, Q.; Xia, Z.; Liang, M.; Zhou, J.; Lin, B.; Zou, C. Dynamic changes in fermentation profiles and bacterial community composition during sugarcane top silage fermentation: A preliminary study. Bioresour Technol. 2019, 285, 121315. [Google Scholar] [CrossRef]
- Ziganshina, E.E.; Bagmanova, A.R.; Khilyas, I.V.; Ziganshin, A.M. Assessment of a biogas-generating microbial community in a pilot-scale anaerobic reactor. J. Biosci. Bioeng. 2014, 117, 730–736. [Google Scholar] [CrossRef]
- Pang, L.; He, Y.; Liu, X.; Li, J.; Yang, P. The role of a newly isolated strain Corynebacterium pollutisoli SPH6 in waste activated sludge alkaline fermentation. Chemosphere 2020, 241, 125072. [Google Scholar] [CrossRef]
- Stiles, M.E.; Holzapfel, W.H. Lactic acid bacteria of foods and their current taxonomy. Int. J. Food Microbiol. 1997, 36, 1–29. [Google Scholar] [CrossRef]
- Johnson, J.M.; Ludwig, A.; Furch, A.C.U.; Mithofer, A.; Scholz, S.; Reichelt, M.; Oelmuller, R. The Beneficial Root-Colonizing Fungus Mortierella hyalina Promotes the Aerial Growth of Arabidopsis and Activates Calcium-Dependent Responses That Restrict Alternaria brassicae-Induced Disease Development in Roots. Mol. Plant-Microbe Interact. 2019, 32, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Ozimek, E. Mortierella Species as the Plant Growth-Promoting Fungi Present in the Agricultural Soils. Agriculture 2021, 11, 7. [Google Scholar] [CrossRef]
- Gomez Exposito, R.; de Bruijn, I.; Postma, J.; Raaijmakers, J.M. Current Insights into the Role of Rhizosphere Bacteria in Disease Suppressive Soils. Front. Microbiol. 2017, 8, 2529. [Google Scholar] [CrossRef]
- Duo-Chuan, L.I.; Chen, S.; Jing, L.U. Purification and partial characterization of two chitinases from the mycoparasitic fungus Talaromyces flavus. Mycopathologia 2005, 159, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Yang, Q.; Hu, S.; Zhang, J.D.; Ma, J. Cloning and characterization of a novel chitinase gene (chi46) from Chaetomium globosum and identification of its biological activity. Appl. Microbiol. Biotechnol. 2008, 80, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, K.; Bentley, I.S.; Tighe, E.; McGleenan, E.; Gaffney, D.; Walsh, G. Recombinant production and characterisation of two chitinases from Rasamsonia emersonii, and assessment of their potential industrial applicability. Appl. Microbiol. Biotechnol. 2021, 105, 7769–7783. [Google Scholar] [CrossRef] [PubMed]
- Zang, J.; Xu, Y.; Xia, W.; Yu, D.; Gao, P.; Jiang, Q.; Yang, F. Dynamics and diversity of microbial community succession during fermentation of Suan yu, a Chinese traditional fermented fish, determined by high throughput sequencing. Food Res. Int. 2018, 111, 565–573. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genera/Lowest Annotation | Total * | Control | Soil | Shell | Taxonomy |
|---|---|---|---|---|---|
| 16S ASVs | |||||
| Sanguibacter | 3.095 | 0.000 | 0.272 | 5.820 | Actinobacteriota Actinobacteria Micrococcales Sanguibacteraceae |
| Pseudomonas, 4 ** | 3.140 | 0.006 | 1.738 | 5.456 | Proteobacteria Gammaproteobacteria Pseudomonadales Pseudomonadaceae |
| Streptomyces, 2 | 2.247 | 0.000 | 0.428 | 4.156 | Actinobacteriota Actinobacteria Streptomycetales Streptomycetaceae |
| Flavobacterium | 1.380 | 0.000 | 0.557 | 2.462 | Bacteroidota Bacteroidia Flavobacteriales Flavobacteriaceae |
| Tissierella, 4 | 1.165 | 0.000 | 0.641 | 2.026 | Firmicutes Clostridia Peptostreptococcales-Tissierellales |
| Dongia, 2 | 1.182 | 0.000 | 1.980 | 1.651 | Proteobacteria Alphaproteobacteria Dongiales Dongiaceae |
| Vagococcus | 0.792 | 0.000 | 0.499 | 1.358 | Firmicutes Bacilli Lactobacillales Vagococcaceae |
| Ruminococcaceae | 0.628 | 0.000 | 0.212 | 1.132 | Firmicutes Clostridia Oscillospirales |
| Chryseolinea | 0.935 | 0.000 | 2.219 | 1.108 | Bacteroidota Bacteroidia Cytophagales Microscillaceae |
| Micrococcaceae | 0.516 | 0.000 | 0.331 | 0.884 | Actinobacteriota Actinobacteria Micrococcales |
| Clostridium, 2 | 0.475 | 0.000 | 0.655 | 0.706 | Firmicutes Clostridia Clostridiales Clostridiaceae |
| Erysipelothrix | 0.410 | 0.000 | 0.275 | 0.697 | Firmicutes Bacilli Erysipelotrichales Erysipelotrichaceae |
| Stenotrophomonas | 0.346 | 0.000 | 0.068 | 0.640 | Proteobacteria Gammaproteobacteria Xanthomonadales Xanthomonadaceae |
| Microbacteriaceae, 2 | 0.357 | 0.002 | 0.144 | 0.635 | Actinobacteriota Actinobacteria Micrococcales |
| Terrimonas | 0.406 | 0.000 | 0.672 | 0.570 | Bacteroidota Bacteroidia Chitinophagales Chitinophagaceae |
| Chitinophagaceae | 0.334 | 0.000 | 0.225 | 0.569 | Bacteroidota Bacteroidia Chitinophagales |
| Longispora | 0.300 | 0.000 | 0.156 | 0.525 | Actinobacteriota Actinobacteria Micromonosporales Micromonosporaceae |
| Rhizobiales, 2 | 0.336 | 0.000 | 0.464 | 0.499 | Proteobacteria Alphaproteobacteria |
| Ignatzschineria, 2 | 0.279 | 0.000 | 0.144 | 0.488 | Proteobacteria Gammaproteobacteria Cardiobacteriales Wohlfahrtiimonadaceae |
| Leifsonia, 2 | 0.291 | 0.000 | 0.307 | 0.461 | Actinobacteriota Actinobacteria Micrococcales Microbacteriaceae |
| Dokdonella | 0.2 | 0.000 | 0.089 | 0.429 | Proteobacteria Gammaproteobacteria Xanthomonadales Rhodanobacteraceae |
| Peptostreptococcus | 0.245 | 0.000 | 0.141 | 0.424 | Firmicutes Clostridia Peptostreptococcales-Tissierellales Peptostreptococcaceae |
| Rhodanobacteraceae | 0.174 | 0.000 | 0.056 | 0.316 | Proteobacteria Gammaproteobacteria Xanthomonadales |
| Pseudarthrobacter | 0.180 | 0.000 | 0.138 | 0.301 | Actinobacteriota Actinobacteria Micrococcales Micrococcaceae |
| Lysinibacillus | 0.151 | 0.000 | 0.003 | 0.286 | Firmicutes Bacilli Bacillales Planococcaceae |
| Blastocatellaceae | 0.159 | 0.000 | 0.103 | 0.273 | Acidobacteriota Blastocatellia Blastocatellales |
| Lysobacter | 0.156 | 0.000 | 0.082 | 0.272 | Proteobacteria Gammaproteobacteria Xanthomonadales Xanthomonadaceae |
| Bryobacter | 0.164 | 0.000 | 0.154 | 0.267 | Acidobacteriota Acidobacteriae Bryobacterales Bryobacteraceae |
| Subgroup 10 | 0.195 | 0.000 | 0.372 | 0.259 | Acidobacteriota Thermoanaerobaculia Thermoanaerobaculales Thermoanaerobaculaceae |
| Psychrobacter | 0.148 | 0.000 | 0.077 | 0.258 | Proteobacteria Gammaproteobacteria Pseudomonadales Moraxellaceae |
| Shinella | 0.140 | 0.000 | 0.050 | 0.251 | Proteobacteria Alphaproteobacteria Rhizobiales Rhizobiaceae |
| Flavihumibacter | 0.230 | 0.000 | 0.622 | 0.249 | Bacteroidota Bacteroidia Chitinophagales Chitinophagaceae |
| Phenylobacterium, 2 | 0.132 | 0.000 | 0.050 | 0.237 | Proteobacteria Alphaproteobacteria Caulobacterales Caulobacteraceae |
| Paeniglutamicibacter | 0.129 | 0.000 | 0.054 | 0.230 | Actinobacteriota Actinobacteria Micrococcales Micrococcaceae |
| Psychrobacillus | 0.113 | 0.000 | 0.012 | 0.212 | Firmicutes Bacilli Bacillales Planococcaceae |
| Solirubrobacteraceae | 0.156 | 0.000 | 0.328 | 0.198 | Actinobacteriota Thermoleophilia Solirubrobacterales |
| Sandaracinus | 0.106 | 0.000 | 0.035 | 0.192 | Myxococcota Polyangia Polyangiales Sandaracinaceae |
| Methylomonadaceae | 0.130 | 0.000 | 0.219 | 0.182 | Proteobacteria Gammaproteobacteria Methylococcales |
| Lachnospiraceae | 0.102 | 0.000 | 0.068 | 0.173 | Firmicutes Clostridia Lachnospirales |
| Allocatelliglobosispora | 0.079 | 0.000 | 0.003 | 0.149 | Actinobacteriota Actinobacteria Micromonosporales Micromonosporaceae |
| Romboutsia | 0.099 | 0.000 | 0.141 | 0.147 | Firmicutes Clostridia Peptostreptococcales-Tissierellales Peptostreptococcaceae |
| Bosea | 0.076 | 0.000 | 0.048 | 0.130 | Proteobacteria Alphaproteobacteria Rhizobiales Beijerinckiaceae |
| Flavitalea | 0.072 | 0.000 | 0.041 | 0.125 | Bacteroidota Bacteroidia Chitinophagales Chitinophagaceae |
| Sphingomonadaceae | 0.087 | 0.000 | 0.145 | 0.122 | Proteobacteria Alphaproteobacteria Sphingomonadales Sphingomonadaceae |
| Microscillaceae | 0.107 | 0.000 | 0.316 | 0.108 | Bacteroidota Bacteroidia Cytophagales |
| Pedobacter | 0.057 | 0.000 | 0.061 | 0.090 | Bacteroidota Bacteroidia Sphingobacteriales Sphingobacteriaceae |
| ITS2 ASVs | |||||
| Mortierella, 5 | 16.213 | 0.804 | 9.626 | 30.505 | Mortierellomycota Mortierellomycetes Mortierellales Mortierellaceae |
| Talaromyces, 8 | 4.836 | 0.006 | 6.995 | 5.054 | Ascomycota Eurotiomycetes Eurotiales Trichocomaceae |
| Mycochlamys, 2 | 4.784 | 0.003 | 7.923 | 3.990 | Ascomycota Sordariomycetes Microascales Microascaceae |
| Pseudeurotium | 2.998 | 0.028 | 5.280 | 2.169 | Ascomycota Leotiomycetes Thelebolales Pseudeurotiaceae |
| Fusicolla, 2 | 1.204 | 0.020 | 1.082 | 1.914 | Ascomycota Sordariomycetes Hypocreales Nectriaceae |
| Sordariales | 1.157 | 0.006 | 1.195 | 1.691 | Ascomycota Sordariomycetes |
| Pseudogymnoascus | 1.017 | 0.028 | 0.944 | 1.583 | Ascomycota Leotiomycetes Thelebolales Pseudeurotiaceae |
| Zopfiella | 2.651 | 0.000 | 5.046 | 1.549 | Ascomycota Sordariomycetes Sordariales Chaetomiaceae |
| Apiotrichum | 0.724 | 0.006 | 0.708 | 1.096 | Basidiomycota Tremellomycetes Trichosporonales Trichosporonaceae |
| Scedosporium | 0.456 | 0.000 | 0.237 | 0.905 | Ascomycota Sordariomycetes Microascales Microascaceae |
| Trichosporonaceae | 0.365 | 0.003 | 0.156 | 0.756 | Basidiomycota Tremellomycetes Trichosporonales |
| Coniochaeta, 2 | 1.118 | 0.026 | 2.129 | 0.641 | Ascomycota Sordariomycetes Coniochaetales Coniochaetaceae |
| Aspergillus, 3 | 1.250 | 0.001 | 2.569 | 0.538 | Ascomycota Eurotiomycetes Eurotiales Aspergillaceae |
| Coprinopsis | 0.542 | 0.000 | 0.827 | 0.525 | Basidiomycota Agaricomycetes Agaricales Psathyrellaceae |
| Humicola | 0.604 | 0.000 | 1.049 | 0.456 | Ascomycota Sordariomycetes Sordariales Chaetomiaceae |
| Rasamsonia | 0.443 | 0.000 | 0.695 | 0.408 | Ascomycota Eurotiomycetes Eurotiales Trichocomaceae |
| Paracremonium | 0.407 | 0.000 | 0.616 | 0.399 | Ascomycota Sordariomycetes Hypocreales Nectriaceae |
| Parascedosporium | 0.308 | 0.000 | 0.467 | 0.300 | Ascomycota Sordariomycetes Microascales Microascaceae |
| Cephalotrichum, 2 | 0.212 | 0.000 | 0.278 | 0.252 | Ascomycota Sordariomycetes Microascales Microascaceae |
| Thermomyces | 0.421 | 0.000 | 0.810 | 0.237 | Ascomycota Eurotiomycetes Eurotiales Trichocomaceae |
| Pseudeurotiaceae | 0.254 | 0.000 | 0.407 | 0.226 | Ascomycota Leotiomycetes Thelebolales |
| Anguillospora | 0.329 | 0.021 | 0.584 | 0.224 | Ascomycota Leotiomycetes Helotiales Helotiaceae |
| Arthrographis | 0.181 | 0.001 | 0.249 | 0.201 | Ascomycota Dothideomycetes Eremomycetaceae |
| Cystobasidiomycetes | 0.191 | 0.000 | 0.295 | 0.182 | Basidiomycota |
| Candida | 0.204 | 0.008 | 0.343 | 0.162 | Ascomycota Saccharomycetes Saccharomycetales |
| Leotiomycetes | 0.275 | 0.000 | 0.529 | 0.155 | Ascomycota |
| Sugiyamaella | 0.344 | 0.007 | 0.707 | 0.144 | Ascomycota Saccharomycetes Saccharomycetales Trichomonascaceae |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yurgel, S.N.; Nadeem, M.; Cheema, M. Microbial Consortium Associated with Crustacean Shells Composting. Microorganisms 2022, 10, 1033. https://doi.org/10.3390/microorganisms10051033
Yurgel SN, Nadeem M, Cheema M. Microbial Consortium Associated with Crustacean Shells Composting. Microorganisms. 2022; 10(5):1033. https://doi.org/10.3390/microorganisms10051033
Chicago/Turabian StyleYurgel, Svetlana N., Muhammad Nadeem, and Mumtaz Cheema. 2022. "Microbial Consortium Associated with Crustacean Shells Composting" Microorganisms 10, no. 5: 1033. https://doi.org/10.3390/microorganisms10051033
APA StyleYurgel, S. N., Nadeem, M., & Cheema, M. (2022). Microbial Consortium Associated with Crustacean Shells Composting. Microorganisms, 10(5), 1033. https://doi.org/10.3390/microorganisms10051033