
The Parasite Load of Trypanosoma cruzi Modulates Feeding and Defecation Patterns of the Chagas Disease Vector Triatoma infestans

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Triatomine Insects
2.2. Mammal Hosts Infection
2.3. Triatomine Infection
2.4. Feeding and Defecation Activity
2.5. Quantification of Trypanosoma Cruzi DNA
2.6. Statistical Analyses
3. Results
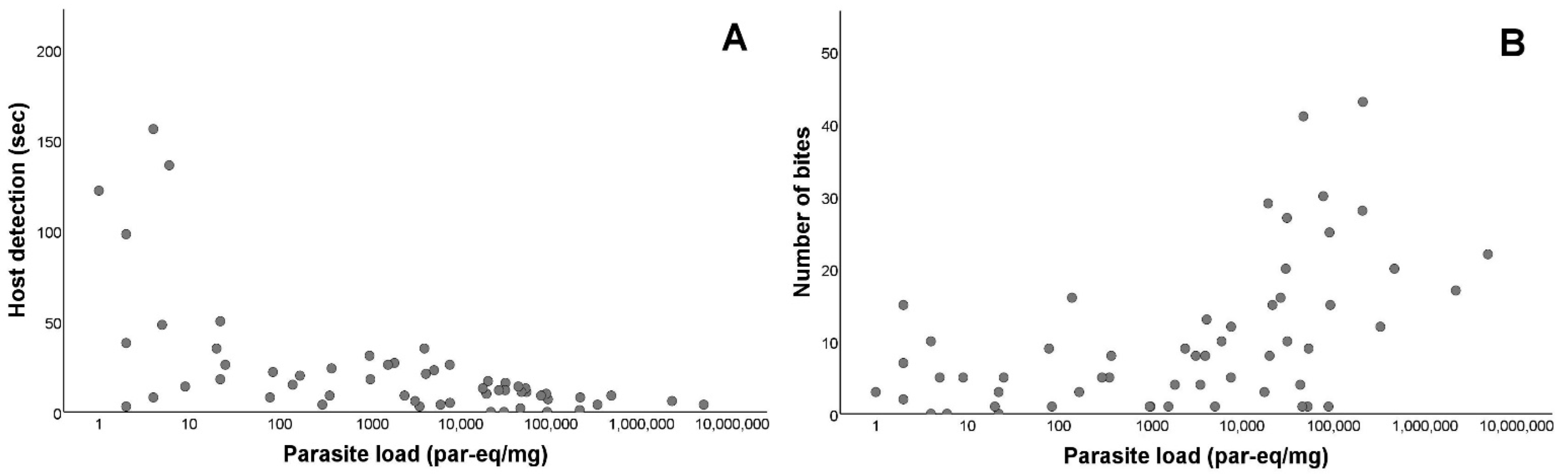
3.1. Infection Status and Parasite Load Quantification
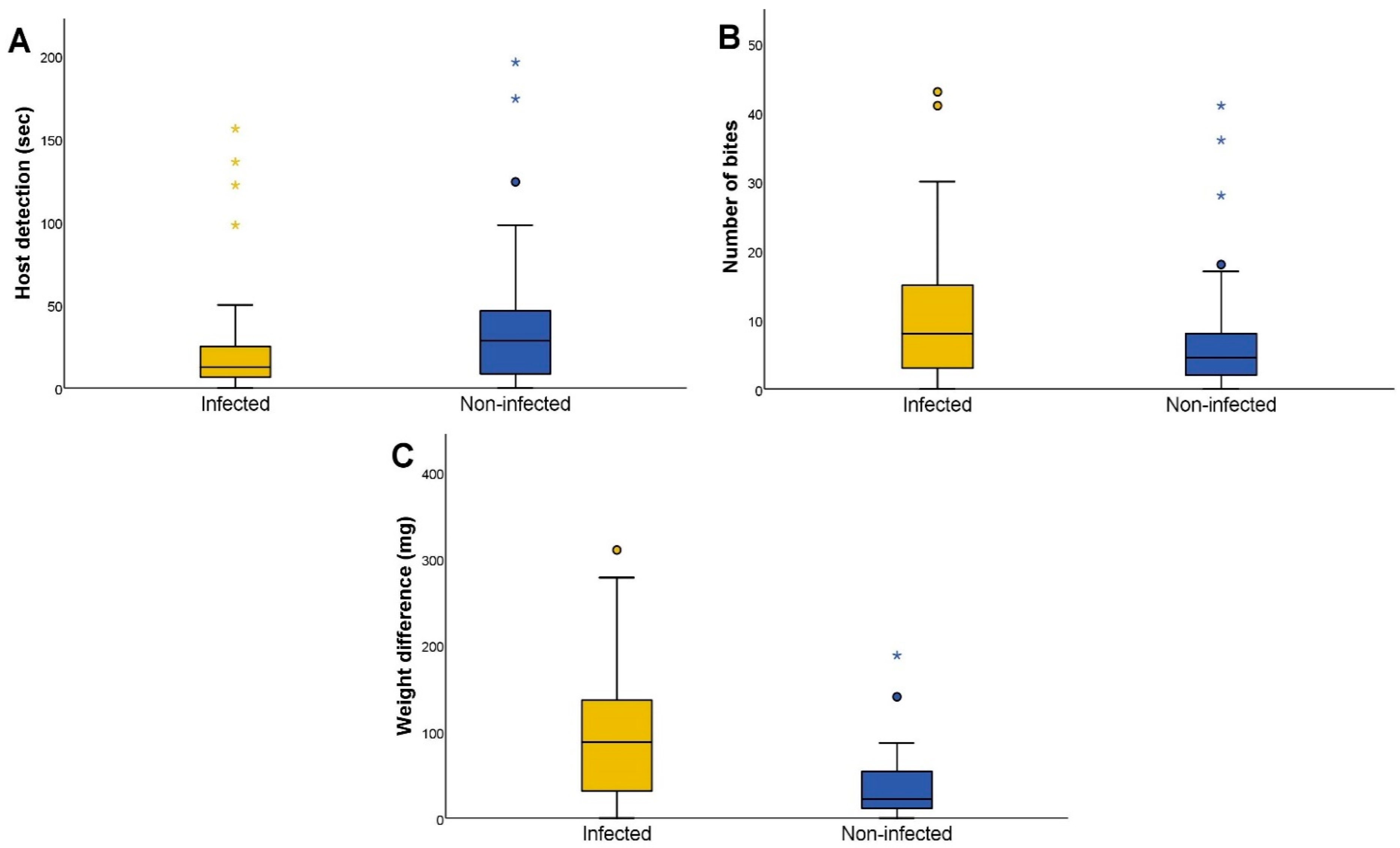
3.2. Feeding Activity of Triatoma Infestans
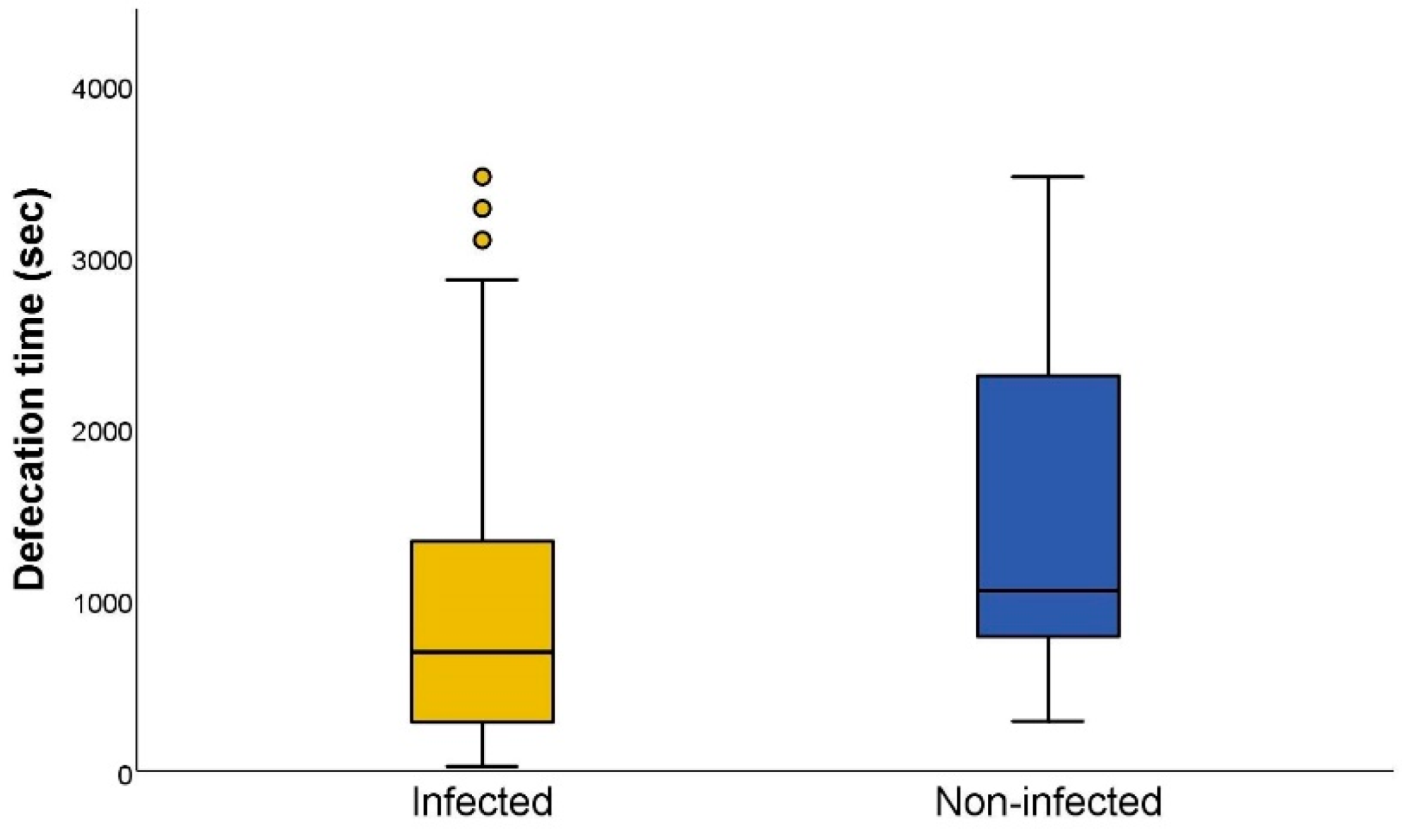
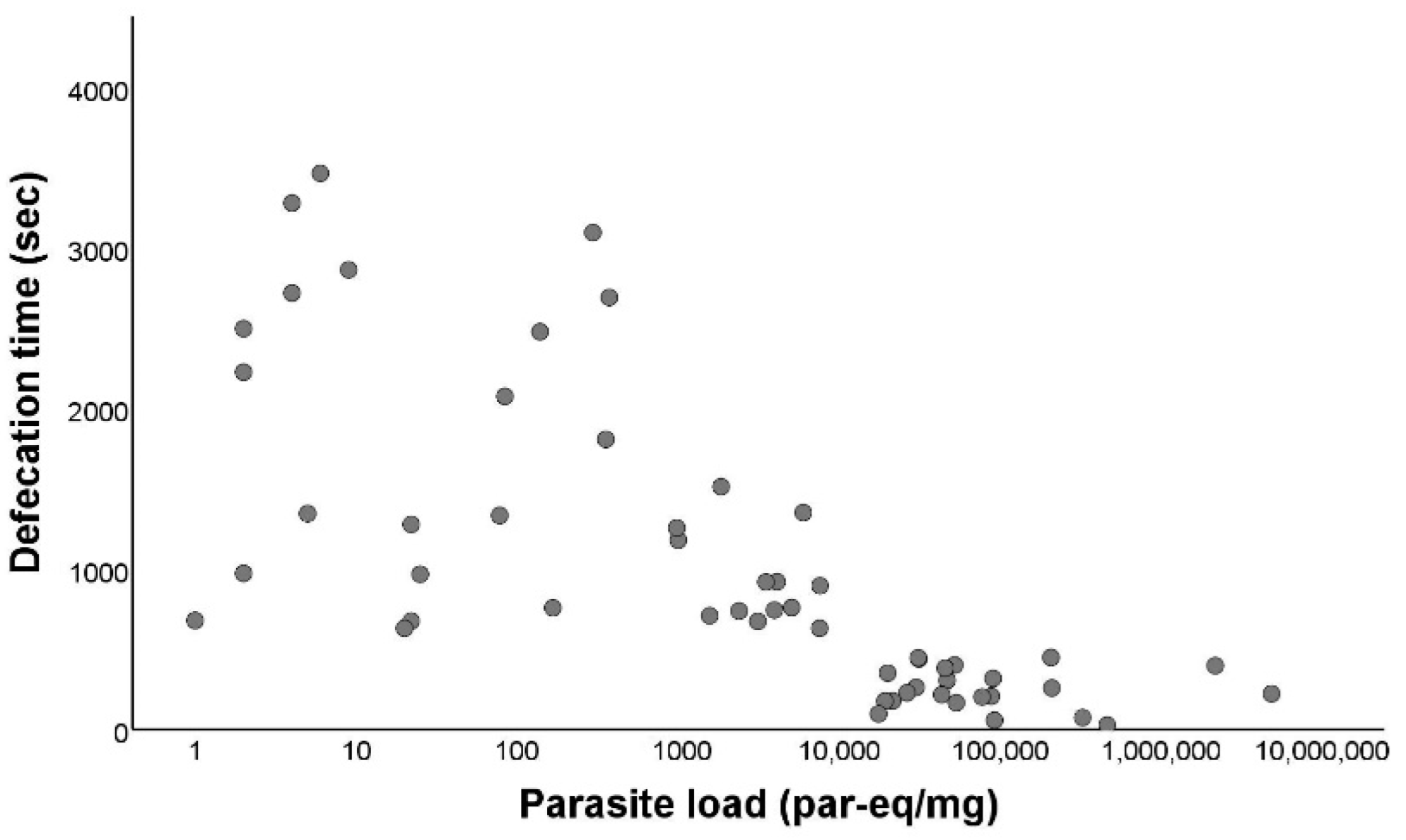
3.3. Defecation Activity of Triatoma Infestans
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poulin, R.; Thomas, F. Phenotypic variability induced by parasites: Parasitol. Today 1999, 15, 28–32. [Google Scholar] [CrossRef]
- Hurd, H. Manipulation of medically important insect vectors by their parasites. Annu. Rev. Entomol. 2003, 48, 141–161. [Google Scholar] [CrossRef] [PubMed]
- WHO. Chagas Disease (Also Known as American Trypanosomiasis). Available online: https://www.who.int/es/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 8 May 2021).
- Noireau, F.; Diosque, P.; Jansen, A.M. Trypanosoma cruzi: Adaptation to its vectors and its hosts. Veter. Res. 2009, 40, 26. [Google Scholar] [CrossRef]
- Guarneri, A.A.; Schaub, G.A. Interaction of triatomines with their bacterial microbiota and trypanosomes. In Triatominae-The Biology of Chagas Disease Vectors; Springer: Berlin/Heidelberg, Germany, 2021; pp. 345–386. [Google Scholar] [CrossRef]
- Lazzari, C.R. Orientation towards hosts in haematophagous insects: An integrative perspective. Adv. Insect Physiol. 2009, 37, 1–58. [Google Scholar] [CrossRef]
- Lazzari, C.R. The behaviour of kissing bugs. In Triatominae-The Biology of Chagas Disease Vectors; Springer: Berlin/Heidelberg, Germany, 2021; pp. 215–238. [Google Scholar] [CrossRef]
- Barrozo, R.B.; Manrique, G.; Lazzari, C.R. The role of water vapour in the orientation behaviour of the blood-sucking bug Triatoma infestans (Hemiptera, Reduviidae). J. Insect. Physiol. 2003, 49, 315–321. [Google Scholar] [CrossRef]
- Barrozo, R.B.; Lazzari, C.R. The response of the blood-sucking bug Triatoma infestans to carbon dioxide and other host odours. Chem. Senses 2004, 29, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Barrozo, R.B.; Reisenman, C.E.; Guerenstein, P.; Lazzari, C.R.; Lorenzo, M.G. An inside look at the sensory biology of triatomines. J. Insect. Physiol. 2017, 97, 3–19. [Google Scholar] [CrossRef]
- Araújo, R.F.; José Mendonça, V.; Rosa, J.A.; Matos, J.F.; Lima, S.C.; de Araújo Figueiredo, M.A. Description of a newly discovered Triatoma infestans (Hemiptera: Reduviidae) Foci in Ibipeba, State of Bahia, Brazil. Rev. Soc. Bras. Med. Trop. 2014, 47, 513–516. [Google Scholar] [CrossRef]
- Zeledon, R.; Beard, C.B.; Dias, J.P.; Leiby, D.A.; Dorn, P.; Coura, J.R. Chapter 2—Triatomine vectors. In An Appraisal of the Status of Chagas Disease in the United States; Elsevier: Walthm, MA, USA, 2012; pp. 5–32. [Google Scholar] [CrossRef]
- Fernandes, M.C.; Andrews, N.W. Host cell invasion by Trypanosoma cruzi: A unique strategy that promotes persistence. FEMS Microbiol. Rev. 2012, 36, 734–747. [Google Scholar] [CrossRef]
- Xavier, S.C.; Roque, A.L.; Bilac, D.; de Araújo, V.A.; da Costa Neto, S.F.; Lorosa, E.S.; da Silva, L.F.; Jansen, A.M. Distantiae transmission of Trypanosoma cruzi: A new epidemiological feature of acute Chagas disease in Brazil. PLoS Negl. Trop. Dis. 2014, 8, e2878. [Google Scholar] [CrossRef]
- Pereyra, N.; Lobbia, P.A.; Mougabure-Cueto, G. Effects of the infection with Trypanosoma cruzi on the feeding and excretion/defecation patterns of Triatoma infestans. Bull. Entomol. Res. 2020, 110, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Zeledón, R.; Alvarado, R.; Jirón, L.F. Observations on the feeding and defecation patterns of three triatomine species (Hemiptera: Reduviidae). Acta Trop. 1977, 34, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Marliére, N.P.; Lorenzo, M.G.; Guarneri, A.A. Trypanosoma cruzi-infected Rhodnius prolixus endure increased predation facilitating parasite transmission to mammal hosts. PLoS Negl. Trop. Dis. 2021, 15, e0009570. [Google Scholar] [CrossRef] [PubMed]
- Botto-Mahan, C.; Cattan, P.E.; Medel, R. Chagas disease parasite induces behavioural changes in the kissing bug Mepraia spinolai. Acta Trop. 2006, 98, 219–223. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Mandel, S. Natural infections and behavior of Trypanosoma rangeli and Trypanosoma cruzi in the vector Rhodnius prolixus in Colombia. J. Parasitol. 1969, 55, 846–852. [Google Scholar] [CrossRef]
- Fellet, M.R.; Lorenzo, M.G.; Elliot, S.L.; Carrasco, D.; Guarneri, A.A. Effects of infection by Trypanosoma cruzi and Trypanosoma rangeli on the reproductive performance of the vector Rhodnius prolixus. PLoS ONE 2014, 9, e105255. [Google Scholar] [CrossRef]
- Marliére, N.P.; Latorre-Estivalis, J.M.; Lorenzo, M.G.; Carrasco, D.; Alves-Silva, J.; Rodrigues, J.e.O.; Ferreira, L.e.L.; Lara, L.e.M.; Lowenberger, C.; Guarneri, A.A. Trypanosomes modify the behavior of their insect hosts: Effects on locomotion and on the expression of a related gene. PLoS Negl. Trop. Dis. 2015, 9, e0003973. [Google Scholar] [CrossRef]
- Pérez, G.; Muñoz-San Martín, C.; Chacón, F.; Bacigalupo, A.; Cattan, P.E.; Solís, R. Modification of the daily activity pattern of the diurnal triatomine Mepraia spinolai (Hemiptera: Reduviidae) Induced by Trypanosoma cruzi (Trypanosomatida: Trypanosomatidae) Infection. J. Med. Entomol. 2021, 58, 2474–2478. [Google Scholar] [CrossRef]
- Ramírez-González, M.G.; Flores-Villegas, A.L.; Salazar-Schettino, P.M.; Gutiérrez-Cabrera, A.E.; Rojas-Ortega, E.; Córdoba-Aguilar, A. Zombie bugs? Manipulation of kissing bug behavior by the parasite Trypanosoma cruzi. Acta Trop. 2019, 200, 105177. [Google Scholar] [CrossRef]
- Chacón, F.; Muñoz-San Martín, C.; Bacigalupo, A.; Álvarez-Duhart, B.; Solís, R.; Cattan, P.E. Trypanosoma cruzi parasite load modulates the circadian activity pattern of Triatoma infestans. Insects 2022, 13, 76. [Google Scholar] [CrossRef]
- Cordero-Montoya, G.; Flores-Villegas, A.L.; Salazar-Schettino, P.M.; Vences-Blanco, M.O.; Rocha-Ortega, M.; Gutiérrez-Cabrera, A.E.; Rojas-Ortega, E.; Córdoba-Aguilar, A. The cost of being a killer’s accomplice: Trypanosoma cruzi impairs the fitness of kissing bugs. Parasitol. Res. 2019, 118, 2523–2529. [Google Scholar] [CrossRef] [PubMed]
- Paranaiba, L.F.; Soares, R.P.; Guarneri, A.A. Triatoma infestans susceptibility to different Trypanosoma cruzi strains: Parasite development and early escape from anterior midgut. Parasitology 2021, 148, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Cummings, K.L.; Tarleton, R.L. Rapid quantitation of Trypanosoma cruzi in host tissue by real-time PCR. Mol. Biochem. Parasitol. 2003, 129, 53–59. [Google Scholar] [CrossRef]
- Piron, M.; Fisa, R.; Casamitjana, N.; López-Chejade, P.; Puig, L.; Vergés, M.; Gascón, J.; Gómez i Prat, J.; Portús, M.; Sauleda, S. Development of a real-time PCR assay for Trypanosoma cruzi detection in blood samples. Acta Trop. 2007, 103, 195–200. [Google Scholar] [CrossRef]
- Elias, M.C.; Vargas, N.; Tomazi, L.; Pedroso, A.; Zingales, B.; Schenkman, S.; Briones, M.R. Comparative analysis of genomic sequences suggests that Trypanosoma cruzi CL Brener contains two sets of non-intercalated repeats of satellite DNA that correspond to T. cruzi I and T. cruzi II types. Mol. Biochem. Parasitol. 2005, 140, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Elias, M.C.; Vargas, N.S.; Zingales, B.; Schenkman, S. Organization of satellite DNA in the genome of Trypanosoma cruzi. Mol. Biochem. Parasitol. 2003, 129, 1–9. [Google Scholar] [CrossRef]
- Dias, F.D.A.; Guerra, B.; Vieira, L.R.; Perdomo, H.D.; Gandara, A.C.P.; Amaral, R.J.V.D.; Vollú, R.E.; Gomes, S.A.O.; Lara, F.A.; Sorgine, M.H.F.; et al. Monitoring of the parasite load in the digestive tract of Rhodnius prolixus by combined qPCR analysis and imaging techniques provides new insights into the trypanosome life cycle. PLoS Negl. Trop. Dis. 2015, 9, e0004186. [Google Scholar] [CrossRef]
- Nogueira, N.P.; Saraiva, F.M.; Sultano, P.E.; Cunha, P.R.; Laranja, G.A.; Justo, G.A.; Sabino, K.C.; Coelho, M.G.; Rossini, A.; Atella, G.C.; et al. Proliferation and differentiation of Trypanosoma cruzi inside its vector have a new trigger: Redox status. PLoS ONE 2015, 10, e0116712. [Google Scholar] [CrossRef]
- Saavedra, M.; Zulantay, I.; Apt, W.; Castillo, J.; Araya, E.; Martínez, G.; Rodríguez, J. Quantification by real-time PCR of Trypanosoma cruzi DNA in samples of Triatoma infestans used in xenodiagnosis of chronic Chagas disease patients. Parasit. Vectors 2016, 9, 382. [Google Scholar] [CrossRef]
- Noireau, F.; Bastrenta, B.; Catala, S.; Dujardin, J.P.; Panzera, F.; Torres, M.; Perez, R.; Galvao, C.; Jurberg, J. Sylvatic population of Triatoma infestans from the Bolivian Chaco: From field collection to characterization. Mem. Inst. Oswaldo Cruz 2000, 95 (Suppl. 1), 119–122. [Google Scholar] [CrossRef]
- PAHO. Información General: Enfermedad de Chagas. Available online: https://www3.paho.org/hq/index.php?option=com_content&view=article&id=5856:2011-informacion-general-enfermedad-chagas&Itemid=40370%E2%9F%A8=fr (accessed on 7 March 2022).
- Dias, J.C.; Silveira, A.C.; Schofield, C.J. The impact of Chagas disease control in Latin America: A review. Mem. Inst. Oswaldo Cruz 2002, 97, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Abad-Franch, F.; Diotaiuti, L.; Gurgel-Gonçalves, R.; Gürtler, R.E. Certifying the interruption of Chagas disease transmission by native vectors: Cui bono? Mem. Inst. Oswaldo Cruz 2013, 108, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.; Botto-Mahan, C.; Ortiz, S.; Acuña, M.; Cattan, P.E.; Solari, A. Trypanosoma cruzi detection in blood by xenodiagnosis and polymerase chain reaction in the wild rodent Octodon degus. Am. J. Trop. Med. Hyg. 2007, 76, 324–326. [Google Scholar] [CrossRef]
- Botto-Mahan, C.; Campos, R.; Acuna-Retamar, M.; Coronado, X.; Cattan, P.E.; Solari, A. Temporal variation of Trypanosoma cruzi infection in native mammals in Chile. Vector Borne Zoonotic Dis. 2010, 10, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Botto-Mahan, C.; Bacigalupo, A.; Correa, J.P.; Oda, E.; Solari, A. Field assessment of Trypanosoma cruzi infection and host survival in the native rodent Octodon degus. Acta Trop. 2012, 122, 164–167. [Google Scholar] [CrossRef]
- Bacigalupo, A.; Torres-Pérez, F.; Segovia, V.; García, A.; Correa, J.P.; Moreno, L.; Arroyo, P.; Cattan, P.E. Sylvatic foci of the Chagas disease vector Triatoma infestans in Chile: Description of a new focus and challenges for control programs. Mem. Inst. Oswaldo Cruz 2010, 105, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Yefi-Quinteros, E.; Muñoz-San Martín, C.; Bacigalupo, A.; Correa, J.P.; Cattan, P.E. Trypanosoma cruzi load in synanthropic rodents from rural areas in Chile. Parasit. Vectors 2018, 11, 171. [Google Scholar] [CrossRef]
- Valenzuela, L.; Sepúlveda, S.; Ponce, I.; Galanti, N.; Cabrera, G. The overexpression of TcAP1 endonuclease confers resistance to infective Trypanosoma cruzi trypomastigotes against oxidative DNA damage. J. Cell Biochem. 2018, 119, 5985–5995. [Google Scholar] [CrossRef]
- Brewer, M.; Garay, M.; Gorla, D.; Murua, F.; Favot, R. Caracterización de los estadios ninfales del genero Triatoma Laporte 1833 I. Triatoma infestans Klug, 1834 (Hemiptera, Reduviidae). Rev. Soc. Entomol. Argent. 1981, 40, 91–102. [Google Scholar]
- Erickson, R.L.; Terzi, M.C.; Jaber, S.M.; Hankenson, F.C.; McKinstry-Wu, A.; Kelz, M.B.; Marx, J.O. Intraperitoneal Continuous-Rate Infusion for the Maintenance of Anesthesia in Laboratory Mice (Mus musculus). J. Am. Assoc. Lab. Anim. Sci. 2016, 55, 548–557. [Google Scholar]
- Paim, R.M.; Pereira, M.H.; Di Ponzio, R.; Rodrigues, J.O.; Guarneri, A.A.; Gontijo, N.F.; Araújo, R.N. Validation of reference genes for expression analysis in the salivary gland and the intestine of Rhodnius prolixus (Hemiptera, Reduviidae) under different experimental conditions by quantitative real-time PCR. BMC Res. Notes 2012, 5, 128. [Google Scholar] [CrossRef] [PubMed]
- Rojo, G.; Sandoval-Rodríguez, A.; López, A.; Ortiz, S.; Correa, J.P.; Saavedra, M.; Botto-Mahan, C.; Cattan, P.E.; Solari, A. Within-host temporal fluctuations of Trypanosoma cruzi discrete typing units: The case of the wild reservoir rodent Octodon degus. Parasites Vectors 2017, 10, 380. [Google Scholar] [CrossRef] [PubMed]
- Jenni, L.; Molyneux, D.H.; Livesey, J.L.; Galun, R. Feeding behaviour of tsetse flies infected with salivarian trypanosomes. Nature 1980, 283, 383–385. [Google Scholar] [CrossRef] [PubMed]
- Beach, R.; Kiilu, G.; Leeuwenburg, J. Modification of sand fly biting behavior by Leishmania leads to increased parasite transmission. Am. J. Trop. Med. Hyg. 1985, 34, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Takano-Lee, M.; Edman, J.D. Lack of manipulation of Rhodnius prolixus (Hemiptera: Reduviidae) vector competence by Trypanosoma cruzi. J. Med. Entomol. 2002, 39, 44–51. [Google Scholar] [CrossRef]
- Garcia, E.S.; Mello, C.B.; Azambuja, P.; Ribeiro, J.M.C. Rhodnius prolixus: Salivary antihemostatic components decrease with Trypanosoma rangeli infection. Exp. Parasitol. 1994, 78, 287–293. [Google Scholar] [CrossRef]
- Estay-Olea, D.; Correa, J.P.; de Bona, S.; Bacigalupo, A.; Quiroga, N.; San Juan, E.; Solari, A.; Botto-Mahan, C. Trypanosoma cruzi could affect wild triatomine approaching behaviour to humans by altering vector nutritional status: A field test. Acta Trop. 2020, 210, 105574. [Google Scholar] [CrossRef]
- Schaub, G.A.; Lösch, P. Parasite/host-interrelationships of the trypanosomatids Trypanosoma cruzi and Blastocrithidia triatomae and the reduviid bug Triatoma infestans: Influence of starvation of the bug. Ann. Trop. Med. Parasitol. 1989, 83, 215–223. [Google Scholar] [CrossRef]
- Sabelis, M.W.; Schippers, P. Variable wind directions and anemotactic strategies of searching for an odour plume. Oecologia 1984, 63, 225–228. [Google Scholar] [CrossRef]
- Taneja, J.; Guerin, P.M. Ammonia attracts the haematophagous bug Triatoma infestans: Behavioural and neurophysiological data on nymphs. J. Comp. Physiol. 1997, 181, 21–34. [Google Scholar] [CrossRef]
- Reisenman, C.E. Hunger is the best spice: Effects of starvation in the antennal responses of the blood-sucking bug Rhodnius prolixus. J. Insect. Physiol. 2014, 71, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Lazzari, C.R.; Núñez, J. The response to radiant heat and the estimation of the temperature of distant sources in Triatoma infestans. J. Insect. Physiol. 1989, 35, 525–529. [Google Scholar] [CrossRef]
- Botto-Mahan, C. Trypanosoma cruzi induces life-history trait changes in the wild kissing bug Mepraia spinolai: Implications for parasite transmission. Vector Borne Zoonotic Dis. 2009, 9, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Verly, T.; Costa, S.; Lima, N.; Mallet, J.; Odêncio, F.; Pereira, M.; Moreira, C.J.C.; Britto, C.; Pavan, M.G. Vector competence and feeding-excretion behavior of Triatoma rubrovaria (Blanchard, 1843) (Hemiptera: Reduviidae) infected with Trypanosoma cruzi TcVI. PLoS Negl. Trop. Dis. 2020, 14, e0008712. [Google Scholar] [CrossRef]
- Lent, H.; Wygodzinsky, P.W. Revision of the Triatominae (Hemiptera, Reduviidae), and their significance as vectors of Chagas’ disease. Bull. Am. Mus. Nat. Hist. 1979, 163, 130–138. [Google Scholar]
- Gürtler, R.E.; Fernández, M.D.P.; Cecere, M.C.; Cohen, J.E. Body size and hosts of Triatoma infestans populations affect the size of bloodmeal contents and female fecundity in rural northwestern Argentina. PLoS Negl. Trop. Dis. 2017, 11, e0006097. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, J.E. Vital statistics of Triatominae (Hemiptera: Reduviidae) under laboratory conditions. I. Triatoma infestans Klug. J. Med. Entomol. 1972, 9, 351–370. [Google Scholar] [CrossRef] [PubMed]
- Schenone, H.; Rojas Mercado, A.; Castillo, D. Estudio comparativo de la sensibilidad y mortalidad de las ninfas III y IV de Triatoma infestans usadas en el xenodiagnóstico de pacientes crónicos. Bol. Chil. Parasitol. 2000, 55, 14–17. [Google Scholar] [CrossRef]
- Trumper, E.V.; Gorla, D.E. Density-dependent timing of defaecation by Triatoma infestans. Trans. R. Soc. Trop. Med. Hyg. 1991, 85, 800–802. [Google Scholar] [CrossRef]
- Crocco, L.B.; Catalá, S.S. Feeding and defaecation patterns in Triatoma sordida. Mem. Inst. Oswaldo Cruz 1996, 91, 409–413. [Google Scholar] [CrossRef]
- Rodríguez, C.S.; Carrizo, S.A.; Crocco, L.B. Comparison of feeding and defecation patterns between fifth-instar nymphs of Triatoma patagonica (Del Ponte, 1929) and Triatoma infestans (Klug, 1934) under laboratory conditions. Rev. Soc. Bras. Med. Trop. 2008, 41, 330–333. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Piesman, J.; Sherlock, I.A. Factors controlling the volume of feces produced by triatomine vectors of Chagas’ disease. Acta Trop. 1983, 40, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Sterkel, M.; Oliveira, J.H.M.; Bottino-Rojas, V.; Paiva-Silva, G.O.; Oliveira, P.L. The Dose Makes the Poison: Nutritional overload determines the life traits of blood-feeding arthropods. Trends Parasitol. 2017, 33, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Melo, R.d.F.P.; Guarneri, A.A.; Silber, A.M. The Influence of environmental cues on the development of Trypanosoma cruzi in triatominae Vector. Front Cell Infect. Microbiol. 2020, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.B.B.; Alevi, K.C.C.; Imperador, C.H.L.; Madeira, F.F.; Azeredo-Oliveira, M.T.V.d. Parasite-vector interaction of Chagas disease: A mini-review. Am. J. Trop. Med. Hyg. 2018, 98, 653–655. [Google Scholar] [CrossRef]
- Ihle-Soto, C.; Costoya, E.; Correa, J.P.; Bacigalupo, A.; Cornejo-Villar, B.; Estadella, V.; Solari, A.; Ortiz, S.; Hernández, H.J.; Botto-Mahan, C.; et al. Spatio-temporal characterization of Trypanosoma cruzi infection and discrete typing units infecting hosts and vectors from non-domestic foci of Chile. PLoS Negl. Trop. Dis. 2019, 13, e0007170. [Google Scholar] [CrossRef]
- Poulin, R. Parasite manipulation of host behavior: An update and frequently asked questions. Adv. Study Behav. 2010, 41, 151–186. [Google Scholar] [CrossRef]
- Goddard, J. Infectious Diseases and Arthropods, 3rd ed.; Humana Press: Cham, Switzerland, 2018; pp. 33–34. [Google Scholar] [CrossRef]
- Libersat, F.; Delago, A.; Gal, R. Manipulation of host behavior by parasitic insects and insect parasites. Annu. Rev. Entomol. 2009, 54, 189–207. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infection Status | N 1 | Units | Median | Q1 2 | Q3 3 | |
|---|---|---|---|---|---|---|
| Host Detection * | Infected | 56 | Seconds | 12.5 | 6.2 | 25.5 |
| Non-infected | 52 | Seconds | 28.5 | 8.3 | 47.3 | |
| First Approach | Infected | 55 | Seconds | 9.0 | 6.0 | 27.0 |
| Non-infected | 50 | Seconds | 12.0 | 6.8 | 27.1 | |
| Number of Bites * | Infected | 56 | Events | 8.0 | 3.0 | 15.0 |
| Non-infected | 52 | Events | 4.5 | 2.0 | 8.0 | |
| Feeding Time | Infected | 55 | Seconds | 872.0 | 557.0 | 1187.0 |
| Non-infected | 50 | Seconds | 711.0 | 441.8 | 1051.0 | |
| Weight Difference *** | Infected | 56 | Milligrams | 88.0 | 30.0 | 138.3 |
| Non-infected | 52 | Milligrams | 22.1 | 11.0 | 54.3 |
| Infection Status | N 1 | Units | Median | Q1 2 | Q3 3 | |
|---|---|---|---|---|---|---|
| Dejection Time *** | Infected | 55 | Seconds | 675.0 | 262.0 | 1277.0 |
| Non-infected | 50 | Seconds | 1045.5 | 765.0 | 2323.5 | |
| Distance of the Dejection | Infected | 55 | Centimeters | 7.10 | 3.8 | 9.6 |
| Non-infected | 50 | Centimeters | 8.1 | 4.8 | 9.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chacón, F.; Bacigalupo, A.; Álvarez-Duhart, B.; Cattan, P.E.; Solís, R.; Muñoz-San Martín, C. The Parasite Load of Trypanosoma cruzi Modulates Feeding and Defecation Patterns of the Chagas Disease Vector Triatoma infestans. Microorganisms 2022, 10, 1003. https://doi.org/10.3390/microorganisms10051003
Chacón F, Bacigalupo A, Álvarez-Duhart B, Cattan PE, Solís R, Muñoz-San Martín C. The Parasite Load of Trypanosoma cruzi Modulates Feeding and Defecation Patterns of the Chagas Disease Vector Triatoma infestans. Microorganisms. 2022; 10(5):1003. https://doi.org/10.3390/microorganisms10051003
Chicago/Turabian StyleChacón, Francisco, Antonella Bacigalupo, Bárbara Álvarez-Duhart, Pedro E. Cattan, Rigoberto Solís, and Catalina Muñoz-San Martín. 2022. "The Parasite Load of Trypanosoma cruzi Modulates Feeding and Defecation Patterns of the Chagas Disease Vector Triatoma infestans" Microorganisms 10, no. 5: 1003. https://doi.org/10.3390/microorganisms10051003
APA StyleChacón, F., Bacigalupo, A., Álvarez-Duhart, B., Cattan, P. E., Solís, R., & Muñoz-San Martín, C. (2022). The Parasite Load of Trypanosoma cruzi Modulates Feeding and Defecation Patterns of the Chagas Disease Vector Triatoma infestans. Microorganisms, 10(5), 1003. https://doi.org/10.3390/microorganisms10051003