
Antibiotic Resistance and Molecular Profiling of the Clinical Isolates of Staphylococcus aureus Causing Bovine Mastitis from India

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation of Bacteria
2.2. Biochemical and Molecular Characterization of S. aureus Isolates
2.3. Multilocus Sequence Typing (MLST)
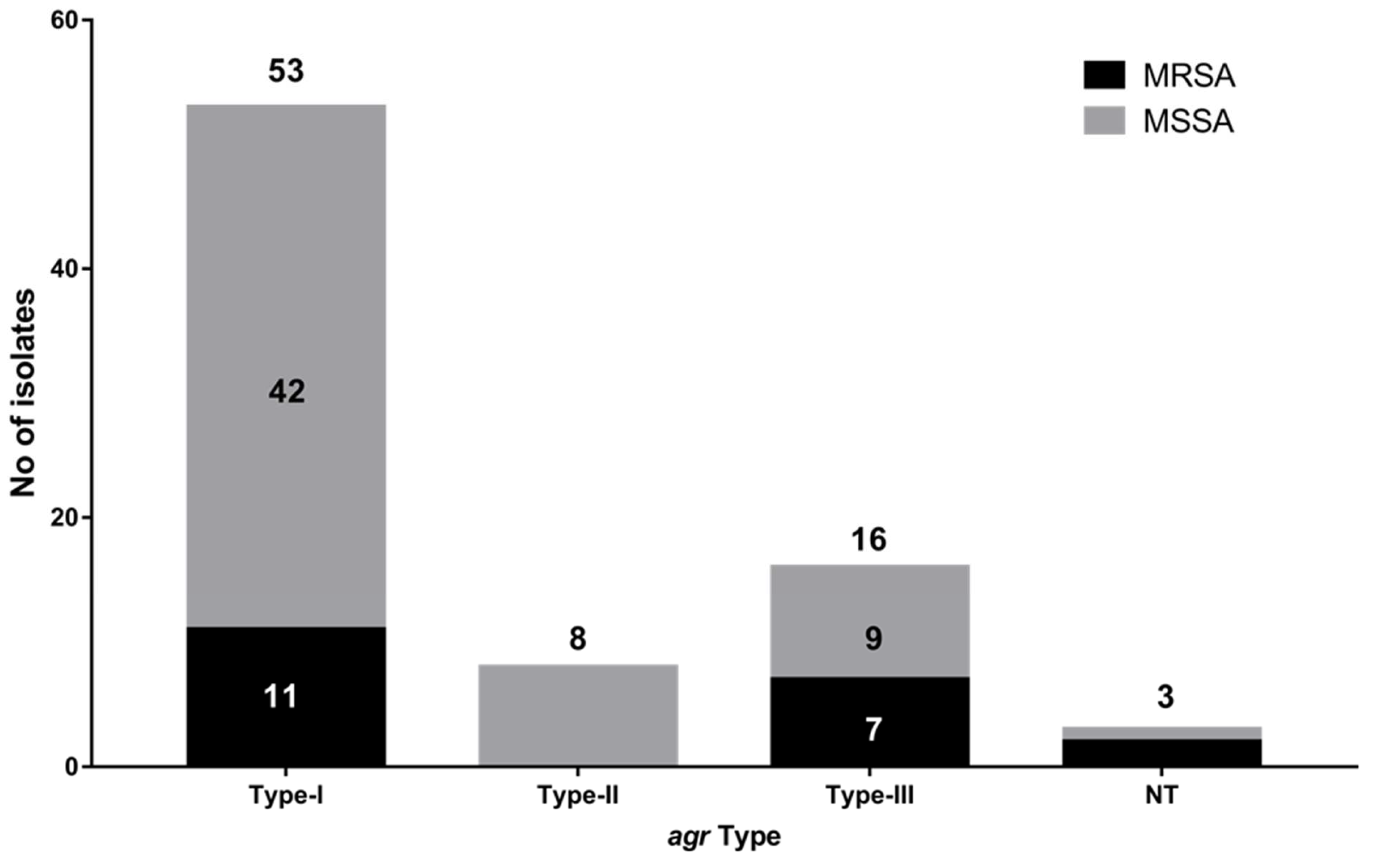
2.4. Accessory Gene Regulator (agr) Typing
2.5. Antibiotic Susceptibility Assay
2.6. Biofilm Formation
2.7. Virulence and Antibiotic Resistance Determinants Gene Profiling
2.8. eBUSRT Analysis
2.9. Statistical Analysis
3. Results
3.1. Antimicrobial Profiling of Clinical Isolates
3.2. Antibiotic Resistance Gene and SCCmec Typing
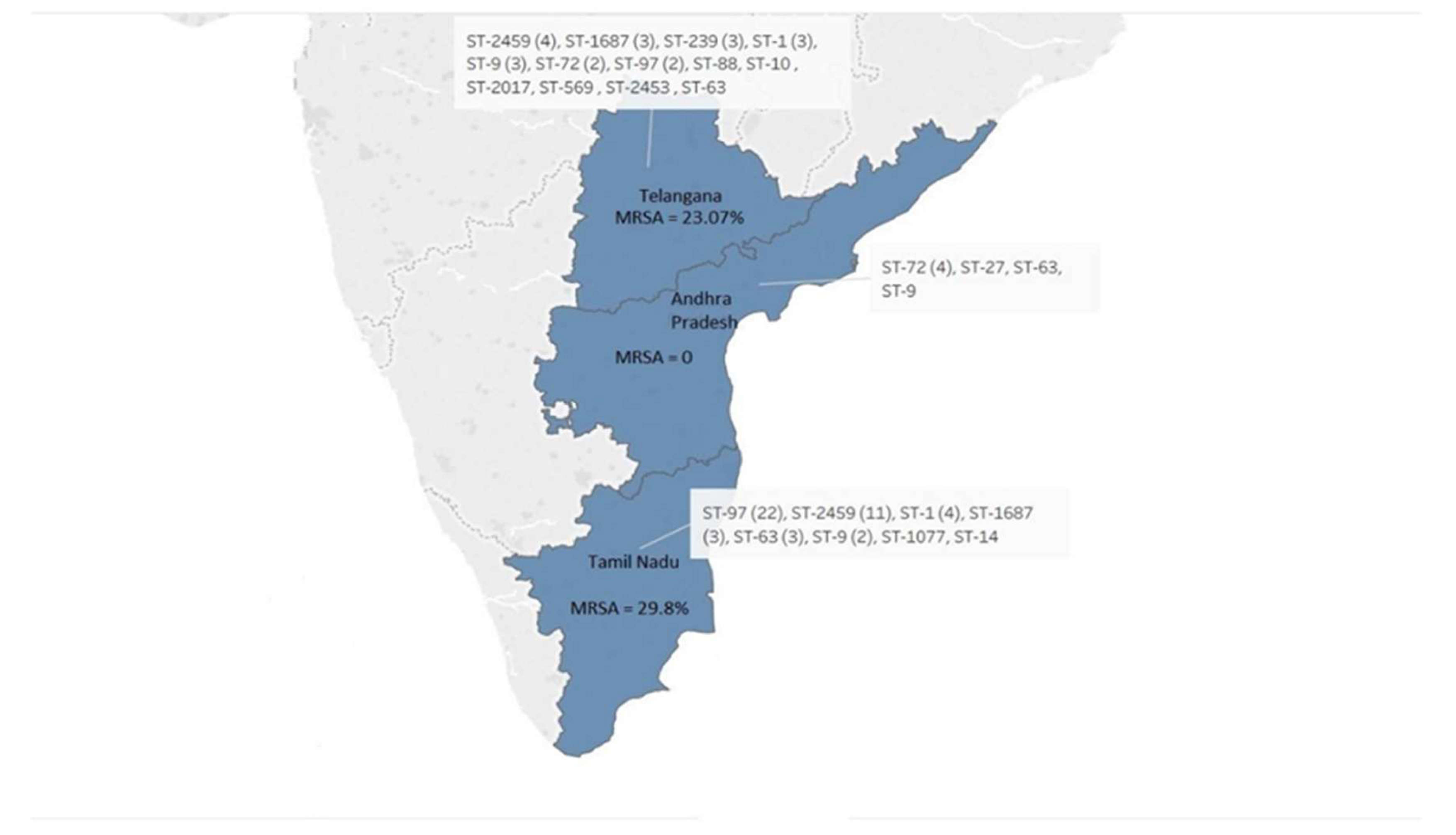
3.3. Molecular Typing of S. aureus Isolates
3.4. Biofilm Production, Virulence and Toxin Genes Profiling
3.5. Significance of Presence and Absence of Gene across S. aureus Isolates
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fluit, A.C. Livestock-associated Staphylococcus aureus. Clin. Microbiol. Infect. 2012, 18, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Mistry, H.; Sharma, P.; Mahato, S.; Saravanan, R.; Kumar, P.A.; Bhandari, V. Prevalence and Characterization of Oxacillin Susceptible mecA-Positive Clinical Isolates of Staphylococcus aureus Causing Bovine Mastitis in India. PLoS ONE 2016, 11, e0162256. [Google Scholar] [CrossRef] [PubMed]
- Hoque, M.N.; Das, Z.C.; Rahman, A.; Haider, M.G.; Islam, M.A. Molecular characterization of Staphylococcus aureus strains in bovine mastitis milk in Bangladesh. Int. J. Vet. Sci. Med. 2018, 6, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Haveri, M.; Roslof, A.; Rantala, L.; Pyorala, S. Virulence genes of bovine Staphylococcus aureus from persistent and nonpersistent intramammary infections with different clinical characteristics. J. Appl. Microbiol. 2007, 103, 993–1000. [Google Scholar] [CrossRef]
- Wang, D.; Wang, Z.; Yan, Z.; Wu, J.; Ali, T.; Li, J.; Lv, Y.; Han, B. Bovine mastitis Staphylococcus aureus: Antibiotic susceptibility profile, resistance genes and molecular typing of methicillin-resistant and methicillin-sensitive strains in China. Infect. Genet. Evol. 2015, 31, 9–16. [Google Scholar] [CrossRef]
- Van Loo, I.; Huijsdens, X.; Tiemersma, E.; de Neeling, A.J.; van de Sande-Bruinsma, N.; Beaujean, D.; Kluytmans, J. Emergence of Methicillin-Resistant Staphylococcus aureus of Animal Origin in Humans. Emerg. Infect. Dis. 2007, 13, 1834–1839. [Google Scholar] [CrossRef]
- Guardabassi, L.; Larsen, J.; Weese, J.S.; Butaye, P.; Battisti, A.; Kluytmans, J.; Lloyd, D.H.; Skov, R.L. Public health impact and antimicrobial selection of meticillin-resistant staphylococci in animals. J. Glob. Antimicrob. Resist. 2013, 1, 55–62. [Google Scholar] [CrossRef]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial biofilms: Development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb. Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef]
- Goyal, R.; Kerketta, P.; Kumar, P.; Rawat, M.; Viswas, K.N.; Agarwal, R.K. Genotypic and Phenotypic Characterization of Clinical Isolates of Staphylococcus aureus for Biofilm Formation Ability. Adv. Anim. Vet. Sci. 2014, 2, 233–238. [Google Scholar] [CrossRef]
- Ikawaty, R.; Brouwer, E.C.; Duijkeren, E.V.; Mevius, D.; Verhoef, J.; Fluit, A.C. Virulence Factors of Genotyped Bovine Mastitis Staphylococcus aureus Isolates in The Netherlands. Int. J. Dairy Sci. 2010, 5, 60–70. [Google Scholar] [CrossRef]
- Buzzola, F.R.; Alvarez, L.P.; Tuchscherr, L.P.; Barbagelata, M.S.; Lattar, S.M.; Calvinho, L.; Sordelli, D.O. Differential abilities of capsulated and noncapsulated Staphylococcus aureus isolates from diverse agr groups to invade mammary epithelial cells. Infect. Immun. 2007, 75, 886–891. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Lu, H.; Wang, X.; Gao, Q.; Dai, Y.; Shang, J.; Li, M. Molecular Characteristics of Staphylococcus aureus Causing Bovine Mastitis between 2014 and 2015. Front. Cell. Infect. Microbiol. 2017, 7, 127. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Michael, G. Nucleic Acid Techniques in Bacterial Systematics; Wiley: Hoboken, NJ, USA, 1991. [Google Scholar]
- Gilot, P.; Lina, G.; Cochard, T.; Poutrel, B. Analysis of the genetic variability of genes encoding the RNA III-activating components Agr and TRAP in a population of Staphylococcus aureus strains isolated from cows with mastitis. J. Clin. Microbiol. 2002, 40, 4060–4067. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, M.P. CLSI Performance Standards for Antimicrobial Susceptibility Testing. CLSI Suppl. M100 2018, 58. Available online: https://file.qums.ac.ir/repository/mmrc/CLSI-2018-M100-S28.pdf (accessed on 10 December 2021).
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Xu, Z.; Liang, Y.; Lin, S.; Chen, D.; Li, B.; Li, L.; Deng, Y. Crystal Violet and XTT Assays on Staphylococcus aureus Biofilm Quantification. Curr. Microbiol. 2016, 73, 474–482. [Google Scholar] [CrossRef]
- Swift, M.L. GraphPad Prism, Data Analysis, and Scientific Graphing. J. Chem. Inf. Comput. Sci. 1997, 37, 411–412. [Google Scholar] [CrossRef]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus Biofilm: An Emerging Battleground in Microbial Communities. Antimicrob. Resist. Infect. 2019, 8, 76. [Google Scholar] [CrossRef]
- Stępień-Pyśniak, D.; Hauschild, T.; Kosikowska, U.; Dec, M.; Urban-Chmiel, R. Biofilm Formation Capacity and Presence of Virulence Factors among Commensal Enterococcus Spp. from Wild Birds. Sci. Rep. 2019, 9, 11204. [Google Scholar] [CrossRef]
- Cuteri, V.; Mezzasoma, P.; Valente, C. Application of biomolecular methods to Staphylococcus aureus strains from dairy cows. Vet. Res. Commun. 2003, 27 (Suppl. 1), 335–338. [Google Scholar] [CrossRef] [PubMed]
- Prashanth, K.; Rao, K.R.; Reddy, P.V.; Saranathan, R.; Makki, A.R. Genotypic Characterization of Staphylococcus aureus Obtained from Humans and Bovine Mastitis Samples in India. J. Glob. Infect. Dis. 2011, 3, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Turutoglu, H.; Ercelik, S.; Ozturk, D. Antibiotic resistance of Staphylococcus aureus and coagulase-negative staphylococci isolated from bovine mastitis. Bull.-Vet. Inst. Pulawy 2006, 50, 41. [Google Scholar]
- Elhassan, M.M.; Ozbak, H.A.; Hemeg, H.A.; Elmekki, M.A.; Ahmed, L.M. Absence of the MecA Gene in Methicillin Resistant Staphylococcus Aureus Isolated from Different Clinical Specimens in Shendi City, Sudan. Biomed. Res. Int. 2015, 2015, 895860. [Google Scholar] [CrossRef] [PubMed]
- Ariza-Miguel, J.; Hernández, M.; Fernández-Natal, I.; Rodríguez-Lázaro, D. Methicillin-Resistant Staphylococcus Aureus Harboring MecC in Livestock in Spain. J. Clin. Microbiol. 2014, 52, 4067–4069. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.D.; Velu, D.; Bhuvana, M.; Krithiga, N.; Banerjee, A.; Shome, R.; Rahman, H.; Ghosh, S.K.; Shome, B.R. Staphylococcus aureus spa type t267, clonal ancestor of bovine subclinical mastitis in India. J. Appl. Microbiol. 2013, 114, 1604–1615. [Google Scholar] [CrossRef]
- Ote, I.; Taminiau, B.; Duprez, J.N.; Dizier, I.; Mainil, J.G. Genotypic characterization by polymerase chain reaction of Staphylococcus aureus isolates associated with bovine mastitis. Vet. Microbiol. 2011, 153, 285–292. [Google Scholar] [CrossRef]
- Soares, B.S.; Melo, D.A.; Motta, C.C.; Marques, V.F.; Barreto, N.B.; Coelho, S.M.O.; Coelho, I.S.; Souza, M.M.S. Characterization of virulence and antibiotic profile and agr typing of Staphylococcus aureus from milk of subclinical mastitis bovine in State of Rio de Janeiro. Arq. Bras. Med. Veterinária Zootec. 2017, 69, 843–850. [Google Scholar] [CrossRef][Green Version]
- Zhang, L.; Gao, J.; Barkema, H.W.; Ali, T.; Liu, G.; Deng, Y.; Naushad, S.; Kastelic, J.P.; Han, B. Virulence gene profiles: Alpha-hemolysin and clonal diversity in Staphylococcus aureus isolates from bovine clinical mastitis in China. BMC Vet. Res. 2018, 14, 63. [Google Scholar] [CrossRef]
- Cucarella, C.; Tormo, M.A.; Ubeda, C.; Trotonda, M.P.; Monzon, M.; Peris, C.; Amorena, B.; Lasa, I.; Penades, J.R. Role of biofilm-associated protein bap in the pathogenesis of bovine Staphylococcus aureus. Infect. Immun. 2004, 72, 2177–2185. [Google Scholar] [CrossRef]
- Zecconi, A.; Cesaris, L.; Liandris, E.; Dapra, V.; Piccinini, R. Role of several Staphylococcus aureus virulence factors on the inflammatory response in bovine mammary gland. Microb. Pathog. 2006, 40, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, N.; Sharma, V.; Shrivastav, A.; Nayak, A.; Rai, A.K. Prevalence and characterization of Panton-Valentine leukocidin-positive Staphylococcus aureus in bovine milk in Jabalpur district of Madhya Pradesh, India. Vet. World 2018, 11, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Kulangara, V.; Nair, N.; Sivasailam, A.; Sasidharan, S.; Kollannur, J.D.; Syam, R. Genotypic and phenotypic beta-lactam resistance and presence of PVL gene in Staphylococci from dry bovine udder. PLoS ONE 2017, 12, e0187277. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Kock, M.M.; Ehlers, M.M. Molecular Characterization of Staphylococcus aureus Isolated from Bovine Mastitis and Close Human Contacts in South African Dairy Herds: Genetic Diversity and Inter-Species Host Transmission. Front. Microbiol. 2017, 8, 511. [Google Scholar] [CrossRef]
- Burke, F.M.; McCormack, N.; Rindi, S.; Speziale, P.; Foster, T.J. Fibronectin-binding protein B variation in Staphylococcus aureus. BMC Microbiol. 2010, 10, 160. [Google Scholar] [CrossRef]
- Kot, B.; Szweda, P.; Frankowska-Maciejewska, A.; Piechota, M.; Wolska, K. Virulence gene profiles in Staphylococcus aureus isolated from cows with subclinical mastitis in eastern Poland. J. Dairy Res. 2016, 83, 228–235. [Google Scholar] [CrossRef]
- Ashraf, S.; Cheng, J.; Zhao, X. Clumping factor A of Staphylococcus aureus interacts with AnnexinA2 on mammary epithelial cells. Sci. Rep. 2017, 7, 40608. [Google Scholar] [CrossRef]
- Ahangari, Z.; Ghorbanpoor, M.; Shapouri, M.R.S.; Gharibi, D.; Ghazvini, K. Methicillin resistance and selective genetic determinants of Staphylococcus aureus isolates with bovine mastitis milk origin. Iran. J. Microbiol. 2017, 9, 152–159. [Google Scholar]
- Wang, W.; Lin, X.; Jiang, T.; Peng, Z.; Xu, J.; Yi, L.; Li, F.; Fanning, S.; Baloch, Z. Prevalence and Characterization of Staphylococcus aureus Cultured From Raw Milk Taken From Dairy Cows With Mastitis in Beijing, China. Front. Microbiol. 2018, 9, 1123. [Google Scholar] [CrossRef]
- Gogoi-Tiwari, J.; Waryah, C.B.; Eto, K.Y.; Tau, M.; Wells, K.; Costantino, P.; Tiwari, H.K.; Isloor, S.; Hegde, N.; Mukkur, T. Relative distribution of virulence-associated factors among Australian bovine Staphylococcus aureus isolates: Potential relevance to development of an effective bovine mastitis vaccine. Virulence 2015, 6, 419–423. [Google Scholar] [CrossRef]
- Klein, R.C.; Fabres-Klein, M.H.; Brito, M.A.; Fietto, L.G.; Ribon Ade, O. Staphylococcus aureus of bovine origin: Genetic diversity, prevalence and the expression of adhesin-encoding genes. Vet. Microbiol. 2012, 160, 183–188. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Primer | Sequence (5′-3′) | Size (Base Pairs) | Annealing Temperature (°C) |
|---|---|---|---|---|
| 1 | bap For | CCCTATATCGAAGGTGTAGAATTGCAC | 971 | 55 |
| 2 | bap Rev | GCTGTTGAAGTTAATACTGTACCTGC | ||
| 3 | clfA For | ATTGGCGTGGCTTCAGTGCT | 292 | 52 |
| 4 | clfA Rev | CGTTTCTTCCGTAGTTGCATTTG | ||
| 5 | clfB For | ACATCAGTAATAGTAGGGGGCAAC | 205 | 52 |
| 6 | clfB Rev | TTCGCACTGTTTGTGTTTGCAC | ||
| 7 | fnbA For | CATAAATTGGGAGCAGCATCA | 127 | 52 |
| 8 | fnbA Rev | ATCAGCAGCTGAATTCCCATT | ||
| 9 | fnbB For | GTAACAGCTAATGGTCGAATTGATACT | 524 | 52 |
| 10 | fnbB Rev | CAAGTTCGATAGGAGTACTATGTTC | ||
| 11 | icaA For | TATACCTTTCTTCGATGTCG | 561 | 48 |
| 12 | icaA Rev | CTTTCGTTATAACAGGCAAG | ||
| 13 | icaD For | AAACGTAAGAGAGGTGG | 381 | 47 |
| 14 | icaD Rev | GGCAATATGATCAAGATAC | ||
| 15 | cna For | TTACACCAGACGGAGCAACA | 498 | 55 |
| 16 | cna Rev | ATGACCCATAGCCTTGTGGA | ||
| 17 | adsA For | GATGACGTTAGAACGCGTGA | 499 | 55 |
| 18 | adsA Rev | CTCTAGGGCCACCGAACATA | ||
| 19 | sbi For | GTTGGGGCAGCAACAATTAC | 497 | 55 |
| 20 | sbi Rev | GCTGCTGATTTATCGTGTGG | ||
| 21 | scn For | ACTTGCGGGAACTTTAGCAA | 319 | 55 |
| 22 | scn Rev | TTTTAGTGCTTCGTCAATTTCG | ||
| 23 | α-hls For | GAAAATGGCATGCACAAAAA | 485 | 55 |
| 24 | α-hls Rev | CCATATACCGGGTTCCAAGA | ||
| 25 | β-hls For | GTGCCAAAGCCGAATCTAAG | 504 | 55 |
| 26 | β-hls Rev | TTTTTCGATCATGTCCAGCA | ||
| 27 | δ-hls For | TAATTAAGGAAGGAGTGATTTCAATG | 100 | 55 |
| 28 | δ-hls Rev | TTTTTAGTGAATTTGTTCACTGTGTC | ||
| 29 | ¥-hls For | AGAAGATATCGGCCAAGGTG | 497 | 55 |
| 30 | ¥-hls Rev | CTTGACCATTCGGTGTAACG | ||
| 31 | lukMF For | CAACTTTGTCGCTAGGTCTAA | 730 | 48 |
| 32 | lukMF Rev | CGCTCGTATCGCCTGAATCTT | ||
| 33 | luk F For | TGCAGCTCAACATATCACACC | 507 | 55 |
| 34 | luk F Rev | GCTTCAACATCCCAACCAAT | ||
| 35 | luk S For | TGAGGTGGCCTTTCCAATAC | 492 | 55 |
| 36 | luk S Rev | CATCCATATTTCTGCCATACG |
| FILE | Van MIC µg/mL | Lin MIC µg/mL | Oxa MIC µg/mL | VAN | LIN | OXA | CX 30 | CD 2 | E 15 | GEN 10 | TE 30 | TEI 30 | RIF 5 | Res/Sus | MRSA/MSSA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SA-1 | 2 | 2 | <0.5 | S | S | S | S | R | S | S | R | S | S | R | MSSA |
| SA-2 | 2 | 2 | <0.5 | S | S | S | S | R | R | R | R | S | S | MDR | MSSA |
| SA-3 | 2 | 2 | <0.5 | S | S | S | S | R | R | R | R | S | S | MDR | MSSA |
| SA-4 | 2 | 2 | <0.5 | S | S | S | S | R | S | S | R | S | S | R | MSSA |
| SA-5 | 1 | 2 | <0.5 | S | S | S | S | R | S | S | S | S | S | R | MSSA |
| SA-6 | 2 | 2 | 0.5 | S | S | S | S | R | S | R | S | S | R | MDR | MSSA |
| SA-7 | 1 | 2 | 0.5 | S | S | S | S | R | S | S | S | S | S | R | MSSA |
| SA-8 | 0.5 | 4 | <0.5 | S | S | S | S | R | R | R | R | S | S | MDR | MSSA |
| SA-9 | 1 | 2 | <0.5 | S | S | S | S | R | R | R | R | S | R | MDR | MSSA |
| SA-10 | 2 | 2 | 1.25 | S | S | S | S | S | R | S | S | S | S | R | MSSA |
| SA-11 | 1 | 2 | <0.5 | S | S | S | S | S | R | S | S | S | R | R | MSSA |
| SA-12 | 1 | 4 | <0.5 | S | S | S | S | S | S | S | S | S | S | S | MSSA |
| SA-13 | 1 | 2 | <0.5 | S | S | S | S | S | S | S | S | S | S | S | MSSA |
| SA-14 | 2 | 2 | <0.5 | S | S | S | S | S | R | R | R | S | S | MDR | MSSA |
| SA-15 | 2 | 2 | <0.5 | S | S | S | S | S | R | R | R | S | S | MDR | MSSA |
| SA-16 | 1 | 2 | <0.5 | S | S | S | S | S | S | S | S | S | S | S | MSSA |
| SA-17 | 1 | 2 | <0.5 | S | S | S | S | S | S | S | R | S | S | R | MSSA |
| SA-18 | 0.5 | 2 | 1.25 | S | S | S | S | R | R | S | S | S | R | MDR | MSSA |
| SA-19 | 1 | 2 | <0.5 | S | S | S | S | R | R | S | S | S | R | MDR | MSSA |
| SA-20 | 1 | 2 | 0.63 | S | S | S | S | S | R | S | R | S | S | R | MSSA |
| SA-21 | 1 | 2 | 0.63 | S | S | S | S | R | R | S | R | S | S | MDR | MSSA |
| SA-22 | 1 | 2 | 0.63 | S | S | S | S | S | R | S | R | S | S | R | MSSA |
| SA-23 | 2 | 4 | 8 | S | S | R | R | S | S | S | R | S | S | MDR | MRSA |
| SA-24 | 1 | 2 | 8 | S | S | R | R | S | S | R | S | S | R | MDR | MRSA |
| SA-25 | 1 | 2 | 0.5 | S | S | R | R | S | S | R | R | S | S | MDR | MRSA |
| SA-26 | 1 | 4 | 0.5 | S | S | S | S | S | S | R | S | R | S | R | MSSA |
| SA-27 | 1 | 2 | 0.5 | S | S | R | R | S | S | R | R | S | S | MDR | MRSA |
| SA-28 | 1 | 4 | 0.5 | S | S | R | R | S | S | R | R | R | S | MDR | MRSA |
| SA-29 | 1 | 4 | 0.5 | S | S | R | R | S | S | R | R | S | S | MDR | MRSA |
| SA-30 | 1 | 4 | 0.25 | S | S | S | S | S | S | R | S | S | S | R | MSSA |
| SA-31 | 2 | 4 | 0.25 | S | S | S | S | S | S | R | S | S | S | R | MSSA |
| SA-32 | 1 | 4 | 0.25 | S | S | S | S | S | S | S | S | R | S | R | MSSA |
| SA-33 | 2 | 4 | 0.25 | S | S | S | S | S | S | R | S | R | R | MDR | MSSA |
| SA-34 | 1 | 2 | 0.13 | S | S | S | S | S | S | R | S | R | S | R | MSSA |
| SA-35 | 2 | 4 | 0.5 | S | S | S | S | S | S | R | S | R | R | MDR | MSSA |
| SA-36 | 1 | 4 | 0.25 | S | S | S | S | S | S | R | S | S | S | R | MSSA |
| SA-37 | 1 | 4 | 0.25 | S | S | S | S | R | S | R | S | R | S | MDR | MSSA |
| SA-38 | 2 | 4 | 8 | S | S | R | R | S | S | R | S | R | R | MDR | MRSA |
| SA-39 | 1 | 4 | 0.5 | S | S | S | S | R | S | R | R | R | S | MDR | MSSA |
| SA-40 | 1 | 2 | 0.25 | S | S | S | S | R | S | R | S | R | S | MDR | MSSA |
| SA-41 | 1 | 2 | 0.5 | S | S | S | S | R | S | R | S | S | R | MDR | MSSA |
| SA-42 | 1 | 4 | 0.5 | S | S | S | S | R | S | R | S | S | S | R | MSSA |
| SA-43 | 1 | 2 | 0.5 | S | S | S | S | S | S | R | S | S | S | R | MSSA |
| SA-44 | 2 | 2 | 8 | S | S | R | R | R | S | R | S | S | S | MDR | MRSA |
| SA-45 | 1 | 2 | 0.25 | S | S | S | S | R | S | R | S | R | S | MDR | MSSA |
| SA-46 | 1 | 2 | 8 | S | S | R | R | R | S | R | S | R | S | MDR | MRSA |
| SA-47 | 1 | 4 | 0.5 | S | S | S | S | S | S | R | S | R | R | MDR | MSSA |
| SA-48 | 2 | 2 | 0.5 | S | S | S | S | S | S | R | S | R | R | MDR | MSSA |
| SA-49 | 1 | 4 | 0.5 | S | S | S | S | S | S | R | S | R | R | MDR | MSSA |
| SA-50 | 1 | 2 | 0.5 | S | S | S | S | S | S | R | S | S | S | R | MSSA |
| SA-51 | 1 | 4 | 8 | S | S | R | R | R | R | R | R | R | R | MDR | MRSA |
| SA-52 | 1 | 2 | <0.5 | S | S | S | S | R | R | S | S | S | R | MDR | MSSA |
| SA-53 | 2 | 2 | <0.5 | S | S | S | S | R | R | S | R | S | R | MDR | MSSA |
| SA-54 | 0.5 | 2 | <0.5 | S | S | S | S | S | R | S | S | S | S | R | MSSA |
| SA-55 | 1 | 2 | <0.5 | S | S | S | S | R | R | S | S | S | S | R | MSSA |
| SA-56 | 0.5 | 2 | <0.5 | S | S | S | S | R | R | S | S | S | S | R | MSSA |
| SA-57 | 0.5 | 2 | <0.5 | S | S | S | S | R | R | S | S | S | S | R | MSSA |
| SA-58 | 1 | 2 | <0.5 | S | S | S | S | R | R | S | S | S | S | R | MSSA |
| SA-59 | 1 | 2 | <0.5 | S | S | S | S | R | S | S | S | S | S | R | MSSA |
| SA-60 | 2 | 2 | 0.5 | S | S | S | S | R | R | S | S | S | S | R | MSSA |
| SA-61 | 1 | 2 | <0.5 | S | S | S | S | R | S | S | R | S | S | R | MSSA |
| SA-62 | 1 | 2 | <0.5 | S | S | S | S | R | R | S | R | S | R | MDR | MSSA |
| SA-63 | 0.5 | 2 | <0.5 | S | S | S | S | R | R | S | R | S | S | MDR | MSSA |
| SA-64 | 0.5 | 2 | <0.5 | S | S | S | S | R | R | S | S | S | S | R | MSSA |
| SA-65 | 1 | 2 | <0.5 | S | S | S | S | R | S | R | S | S | S | R | MSSA |
| SA-66 | 1 | 2 | <0.5 | S | S | S | S | R | S | S | S | S | S | R | MSSA |
| SA-67 | 1 | 4 | 0.25 | S | S | S | S | S | S | R | S | R | R | MDR | MSSA |
| SA-68 | 1 | 4 | 0.5 | S | S | S | S | R | S | R | S | S | R | MDR | MSSA |
| SA-69 | 0.5 | 4 | 8 | S | S | R | R | R | S | S | S | S | R | MDR | MRSA |
| SA-70 | 1 | 4 | 8 | S | S | R | R | S | R | R | S | S | R | MDR | MRSA |
| SA-71 | 1 | 4 | 32 | S | S | R | R | R | R | R | S | S | S | MDR | MRSA |
| SA-72 | 2 | 4 | 32 | S | S | R | R | R | R | R | S | S | R | MDR | MRSA |
| SA-73 | 2 | 4 | 0.5 | S | S | S | S | S | S | R | S | S | R | R | MSSA |
| SA-74 | 1 | 4 | 8 | S | S | R | R | R | S | R | S | S | R | MDR | MRSA |
| SA-75 | 2 | 4 | 0.5 | S | S | R | R | S | S | S | S | S | S | R | MRSA |
| SA-76 | 1 | 4 | 0.5 | S | S | R | R | R | S | R | S | S | R | MDR | MRSA |
| SA-77 | 2 | 4 | 32 | S | S | R | R | R | R | R | S | S | R | MDR | MRSA |
| SA-78 | 1 | 4 | 2 | S | S | S | S | R | R | R | S | S | R | MDR | MSSA |
| SA-79 | 1 | 2 | 16 | S | S | R | R | S | R | R | S | S | R | MDR | MRSA |
| SA-80 | 2 | 2 | 0.5 | S | S | R | R | S | R | R | R | R | S | MDR | MRSA |
| Isolate ID | mecA | mecC | vanA | SCCmec Type |
|---|---|---|---|---|
| SA-23 | + | - | - | IVd |
| SA-24 | + | - | - | II |
| SA-25 | + | - | - | II |
| SA-27 | - | - | - | |
| SA-28 | + | - | - | II |
| SA-29 | + | - | - | II |
| SA-38 | + | - | - | III |
| SA-44 | + | - | - | III |
| SA-46 | + | - | - | III |
| SA-51 | + | - | - | III |
| SA-69 | + | - | - | III |
| SA-70 | + | - | - | III |
| SA-71 | + | - | - | III |
| SA-72 | - | - | - | |
| SA-74 | - | - | - | |
| SA-75 | - | - | - | |
| SA-76 | + | - | - | Iva |
| SA-77 | + | - | - | Iva |
| SA-79 | + | - | - | Iva |
| SA-80 | + | - | - | Iva |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brahma, U.; Suresh, A.; Murthy, S.; Bhandari, V.; Sharma, P. Antibiotic Resistance and Molecular Profiling of the Clinical Isolates of Staphylococcus aureus Causing Bovine Mastitis from India. Microorganisms 2022, 10, 833. https://doi.org/10.3390/microorganisms10040833
Brahma U, Suresh A, Murthy S, Bhandari V, Sharma P. Antibiotic Resistance and Molecular Profiling of the Clinical Isolates of Staphylococcus aureus Causing Bovine Mastitis from India. Microorganisms. 2022; 10(4):833. https://doi.org/10.3390/microorganisms10040833
Chicago/Turabian StyleBrahma, Umarani, Akash Suresh, Shweta Murthy, Vasundhra Bhandari, and Paresh Sharma. 2022. "Antibiotic Resistance and Molecular Profiling of the Clinical Isolates of Staphylococcus aureus Causing Bovine Mastitis from India" Microorganisms 10, no. 4: 833. https://doi.org/10.3390/microorganisms10040833
APA StyleBrahma, U., Suresh, A., Murthy, S., Bhandari, V., & Sharma, P. (2022). Antibiotic Resistance and Molecular Profiling of the Clinical Isolates of Staphylococcus aureus Causing Bovine Mastitis from India. Microorganisms, 10(4), 833. https://doi.org/10.3390/microorganisms10040833