
Interplay between Arabidopsis thaliana Genotype, Plant Growth and Rhizosphere Colonization by Phytobeneficial Phenazine-Producing Pseudomonas chlororaphis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Cultivation
2.2. Mutant Screening
2.3. Bacterial Inoculation in the Rhizosphere
2.4. Rhizosphere Harvest and Sample Processing
2.5. Bacterial Quantification by qPCR
2.6. Phenotypic Characterization of the Root System of Sand-Grown Plants
2.7. Statistical Analyses
3. Results
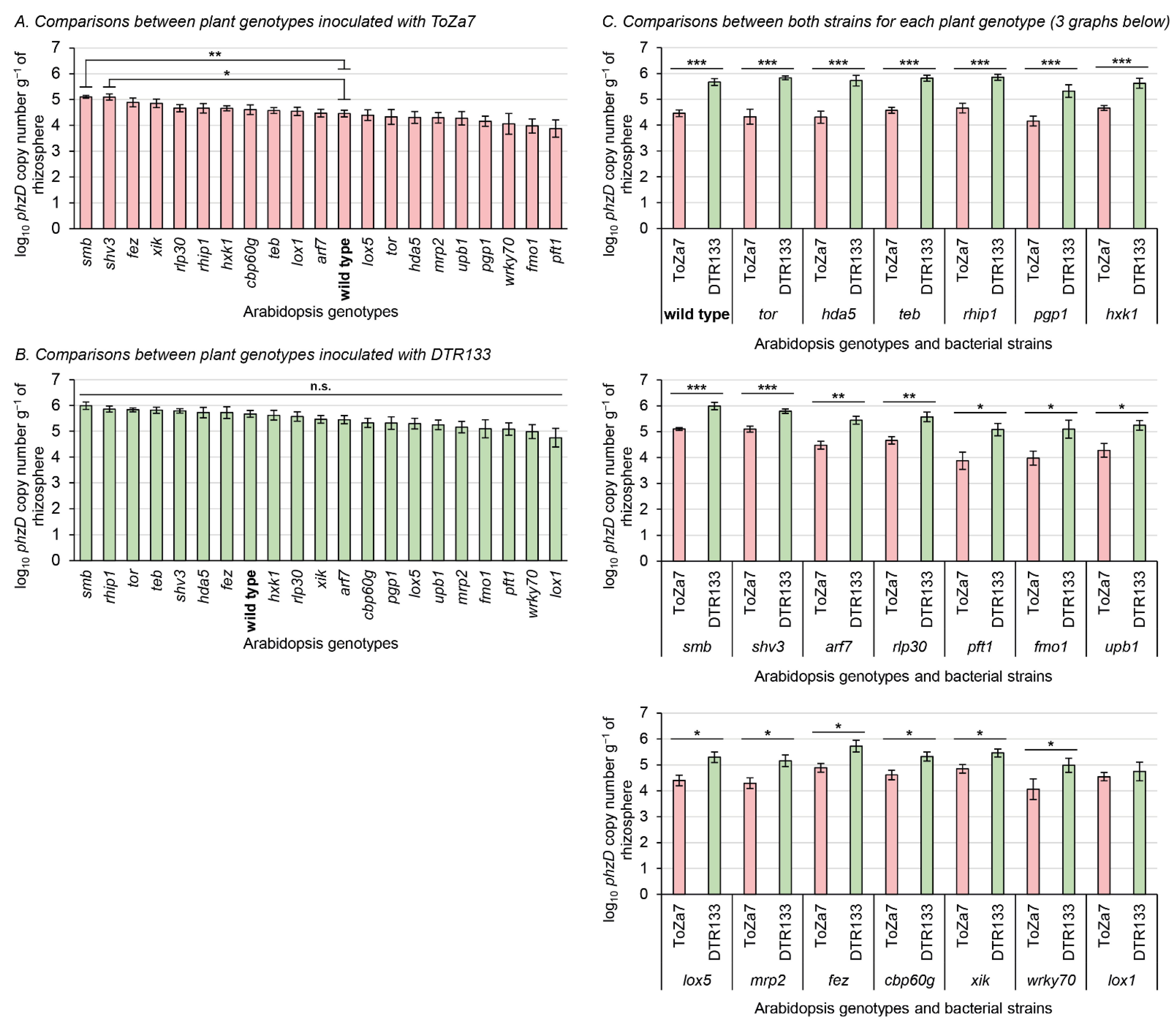
3.1. Rhizosphere Colonization of Mutant Plants by Both Pseudomonas Strains
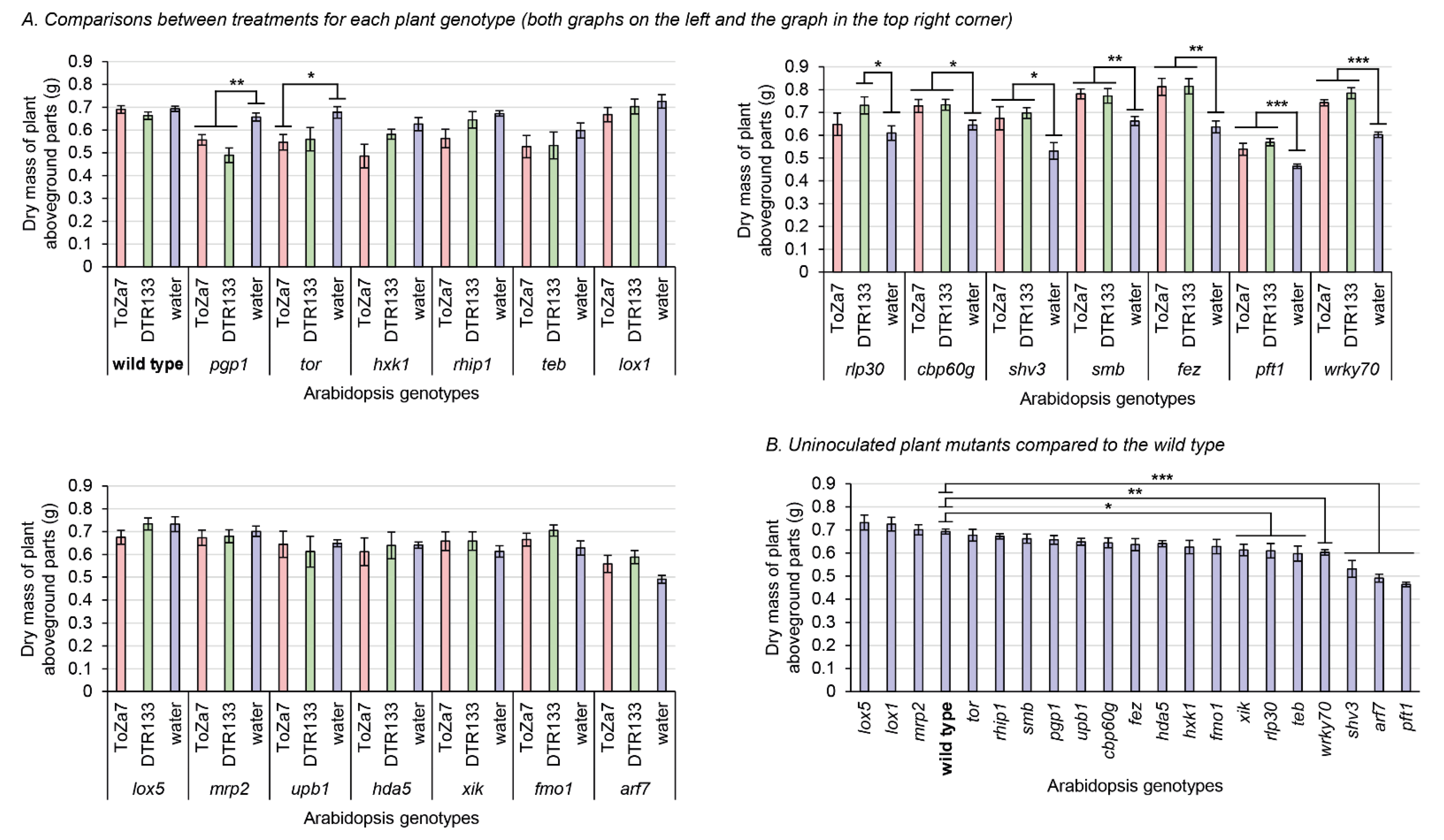
3.2. Aboveground Biomass Accumulation in the Inoculated Mutants Plants
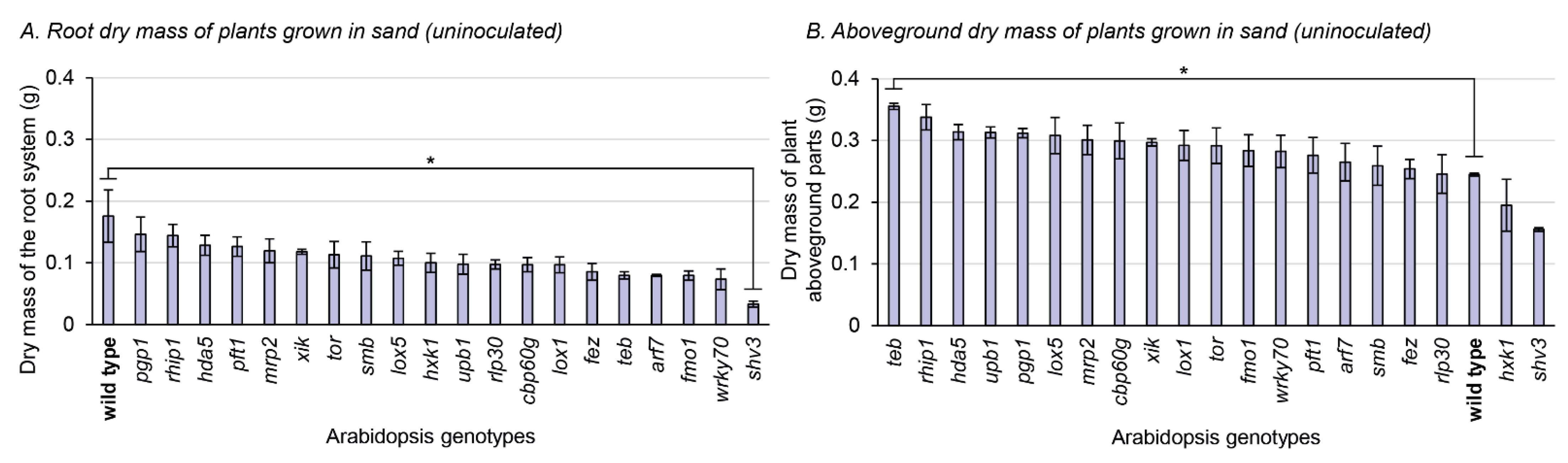
3.3. Phenotypic Characterization of the Mutants Grown in Sterile Sand
4. Discussion
4.1. Rhizosphere Colonization Is Affected by the Plant Genotype
4.2. Bacterial Inoculation on Several Plant Mutants Differently Impacts Plant Biomass
4.3. Changes in Root Exudates Composition Do Not Alter Rhizosphere Colonization
4.4. The teb Mutation Interplays with the Plant Substrate to Affect Aboveground Biomass Accumulation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Affected Gene | Forward Primer | Reverse Primer | Ta (°C) |
|---|---|---|---|---|
| CS66051 | pgp1 | GTTGAAGAACCCGGCGATAC | AGCAACAACTACGGGGAAGA | 53 |
| CS66052 | mrp2 | TGGATGGGTTACATCTATGCG | TGCAATCACTATTCATAGCCAATC | 49 |
| SALK_059431C | lox1-1 | AGCTCCTTGAACCTCACTTCC | GAGACGCTATTTGGAATTCCC | 50 |
| SALK_044826C | lox5-1 | ATCACATGACAGGCCCAATAG | TGATCGATTCGATTCGAAATC | 47 |
| CS69137 | rhip1-1 | CTTCTTCTGCAGAAATGGTGG | CTTTTTCACGAAAATTGGCTG | 47 |
| SALK_093312C | hda5 | ATCTGGGAGAAGCTTCAGCTC | CTGCATTCAGGAAAAGCAAAG | 49 |
| CS69135 | hxk1-3 | TTGTTTTTGATTCCAAATCGG | TCATCAAATGAGGAGGAATCG | 46 |
| CS24607 | arf7-1 | CAGCTAGATCGTTCGAAATGG | AGCACATCACCATTTAGGTGC | 50 |
| SALK_024208C | shv3-2 | GAAGGTTGTCACGAAGACTCG | TCCAAAACAGAAAAATGCTGG | 48 |
| SALK_143526C | smb-3 | GTCGTCATCATCATCTGCATC | GTGTATAACGCGCACACACAC | 50 |
| SALK_007846C | tor | CGTTAACACTTGGACCCTGTC | ACCCCTTTTTGGTTCAACAAC | 51 |
| SALK_018851C | teb-5 | ATTCATTGCTCGGCATCTATG | CGATTCATTGGATTGTTTTGG | 47 |
| SALK_026163C | fmo1-1 | CTTTTCGGTTGGACTTGGAAC | CTGCTTTGGACGTATCCTACG | 51 |
| SALK_025663C | fez-2 | TAGACATTGTGTCGGTGCTTG | GTATAGGAACAGCTCGAGGCC | 52 |
| SALK_025198C | wrky70-1 | TGATCTTCGGAATCCATGAAG | CAAACCACACCAAGAGGAAAG | 49 |
| CS868100 | upb1-1 | CTGAACTTCGAATTTGGATGC | ACACCCTTGTGGACACTTGTC | 49 |
| SALK_129555C | pft1-2 | TGGAACTGGTCCAACAGAAAC | TGCATTGGCTTTCTTCCATAC | 50 |
| SALK_067972C | xik-2 | GGGTAGCAAGATACTCCTCGG | GCAAGAGCAACTCAATTCTGG | 51 |
| SALK_023199C | cbp60g-1 | TCAATGAAGATTCGGAACAGC | ACTTCCGACTCCTAGTCCAGC | 50 |
| CS65465 | rlp30-2 | TCCCGACAAATGAATTCTCAC | TGTCGACGAGAAGCTTAGCTC | 49 |
| LBb1.3—Primer targeting the inserted T-DNA | ATTTTGCCGATTTCGGAAC | Reverse primer of each genotype | 48 | |
References
- Hiltner, L. Über Neuere Erfahrungen Und Probleme Auf Dem Gebiete Der Bodenbakteriologie Unter Besonderer Berücksichtigung Der Gründüngung Und Brache. Arb. Dtsch. Landwirtsch. Ges. 1904, 98, 59–78. [Google Scholar]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going Back to the Roots: The Microbial Ecology of the Rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.J.; Bloemberg, G.V. Life in the Rhizosphere. In Pseudomonas Vol. 1—Genomics, Life Style and Molecular Architecture; Ramos, J.-L., Ed.; Springer: New York, NY, USA, 2004; pp. 403–430. ISBN 1-4613-4788-2. [Google Scholar]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, J.M.; Whipps, J.M. Substrate Flow in the Rhizosphere. Plant Soil 1990, 129, 1–10. [Google Scholar] [CrossRef]
- Neumann, G.; Römheld, V. The Release of Root Exudates as Affected by the Plant Physiological Status. In The Rhizosphere: Biochemistry and Organic Substances at the Soil-Plant Interface; Pinton, R., Varanini, Z., Nannipieri, P., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 23–72. [Google Scholar]
- Badri, D.V.; Vivanco, J.M. Regulation and Function of Root Exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Bakker, M.G.; Sugiyama, A.; Manter, D.K.; Vivanco, J.M. Root Exudation of Phytochemicals in Arabidopsis Follows Specific Patterns That Are Developmentally Programmed and Correlate with Soil Microbial Functions. PLoS ONE 2013, 8, e55731. [Google Scholar] [CrossRef]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loqué, D.; Bowen, B.P.; et al. Dynamic Root Exudate Chemistry and Microbial Substrate Preferences Drive Patterns in Rhizosphere Microbial Community Assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Carvalhais, L.C.; Dennis, P.G.; Fedoseyenko, D.; Hajirezaei, M.-R.; Borriss, R.; von Wirén, N. Root Exudation of Sugars, Amino Acids, and Organic Acids by Maize as Affected by Nitrogen, Phosphorus, Potassium, and Iron Deficiency. J. Plant. Nutr. Soil Sci. 2011, 174, 3–11. [Google Scholar] [CrossRef]
- Uren, N.C. Types, Amounts, and Possible Functions of Compounds Released into the Rhizosphere by Soil-Grown Plants. In The Rhizosphere: Biochemistry and Organic Substances at the Soil-Plant Interface; Pinton, R., Varanini, Z., Nannipieri, P., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 1–21. [Google Scholar]
- Jaeger, C.H.; Lindow, S.E.; Miller, W.; Clark, E.; Firestone, M.K. Mapping of Sugar and Amino Acid Availability in Soil around Roots with Bacterial Sensors of Sucrose and Tryptophan. Appl. Environ. Microbiol. 1999, 65, 2685–2690. [Google Scholar] [CrossRef] [Green Version]
- Sasse, J.; Martinoia, E.; Northen, T. Feed Your Friends: Do Plant Exudates Shape the Root Microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Reverdy, A.; She, Q.; Sun, B.; Chai, Y. The Role of Rhizodeposits in Shaping Rhizomicrobiome. Environ. Microbiol. Rep. 2019, 12, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.S.; Cavell, A.C.; Dolan, L.; Roberts, K.; Grierson, C.S. Genetic Interactions during Root Hair Morphogenesis in Arabidopsis. Plant Cell 2000, 12, 1961–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertin, C.; Yang, X.; Weston, L.A. The Role of Root Exudates and Allelochemicals in the Rhizosphere. Plant Soil 2003, 256, 67–83. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere Microbiome Assemblage Is Affected by Plant Development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, P.R.; Miller, A.J.; Dennis, P.G. Do Root Exudates Exert More Influence on Rhizosphere Bacterial Community Structure than Other Rhizodeposits. In Molecular Microbial Ecology of the Rhizosphere: Volume 1 & 2; de Bruijn, F.J., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; Volume 1, pp. 229–242. [Google Scholar]
- Zboralski, A.; Biessy, A.; Filion, M. Rhizosphere Colonization by Plant-Beneficial Pseudomonas spp.: Thriving in a Heterogeneous and Challenging Environment. In Advances in PGPR Research; Singh, H.B., Ed.; CAB International: Oxfordshire, UK, 2017; pp. 197–217. [Google Scholar]
- Weston, L.A.; Ryan, P.R.; Watt, M. Mechanisms for Cellular Transport and Release of Allelochemicals from Plant Roots into the Rhizosphere. J. Exp. Bot. 2012, 63, 3445–3454. [Google Scholar] [CrossRef]
- Badri, D.V.; Loyola-Vargas, V.M.; Broeckling, C.D.; De-la-Pena, C.; Jasinski, M.; Santelia, D.; Martinoia, E.; Sumner, L.W.; Banta, L.M.; Stermitz, F.; et al. Altered Profile of Secondary Metabolites in the Root Exudates of Arabidopsis ATP-Binding Cassette Transporter Mutants. Plant Physiol. 2008, 146, 762–771. [Google Scholar] [CrossRef] [Green Version]
- Cox, D.E.; Dyer, S.; Weir, R.; Cheseto, X.; Sturrock, M.; Coyne, D.; Torto, B.; Maule, A.G.; Dalzell, J.J. ABC Transporter Genes ABC-C6 and ABC-G33 Alter Plant-Microbe-Parasite Interactions in the Rhizosphere. Sci. Rep. 2019, 9, 19899. [Google Scholar] [CrossRef]
- Badri, D.V.; Loyola-Vargas, V.M.; Du, J.; Stermitz, F.R.; Broeckling, C.D.; Iglesias-Andreu, L.; Vivanco, J.M. Transcriptome Analysis of Arabidopsis Roots Treated with Signaling Compounds: A Focus on Signal Transduction, Metabolic Regulation and Secretion. New Phytol. 2008, 179, 209–223. [Google Scholar] [CrossRef]
- Doornbos, R.F.; Geraats, B.P.J.; Kuramae, E.E.; Van Loon, L.C.; Bakker, P.A.H.M. Effects of Jasmonic Acid, Ethylene, and Salicylic Acid Signaling on the Rhizosphere Bacterial Community of Arabidopsis Thaliana. Mol. Plant Microbe Interact. 2011, 24, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Lebeis, S.L.; Paredes, S.H.; Lundberg, D.S.; Breakfield, N.; Gehring, J.; McDonald, M.; Malfatti, S.; Del Rio, T.G.; Jones, C.D.; Tringe, S.G.; et al. Salicylic Acid Modulates Colonization of the Root Microbiome by Specific Bacterial Taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef] [Green Version]
- Carvalhais, L.C.; Dennis, P.G.; Badri, D.V.; Kidd, B.N.; Vivanco, J.M.; Schenk, P.M. Linking Jasmonic Acid Signaling, Root Exudates, and Rhizosphere Microbiomes. Mol. Plant Microbe Interact. 2015, 28, 1049–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin-A-Woeng, T.F.C.; Bloemberg, G.V.; Mulders, I.H.M.; Dekkers, L.C.; Lugtenberg, B.J.J. Root Colonization by Phenazine-1-Carboxamide-Producing Bacterium Pseudomonas Chlororaphis PCL1391 Is Essential for Biocontrol of Tomato Foot and Root Rot. Mol. Plant Microbe Interact. 2000, 13, 1340–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloemberg, G.V.; Lugtenberg, B.J. Molecular Basis of Plant Growth Promotion and Biocontrol by Rhizobacteria. Curr. Opin. Plant. Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological Control of Soil-Borne Pathogens by Fluorescent Pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Weller, D.M. Biological Control of Soilborne Plant Pathogens in the Rhizosphere with Bacteria. Annu. Rev. Phytopathol. 1988, 26, 379–407. [Google Scholar] [CrossRef]
- Weller, D.M. Pseudomonas Biocontrol Agents of Soilborne Pathogens: Looking Back over 30 Years. Phytopathology 2007, 97, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Mercado-Blanco, J. Pseudomonas Strains That Exert Biocontrol of Plant Pathogens. In Pseudomonas; Ramos, J.-L., Goldberg, J.B., Filloux, A., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 121–172. ISBN 94-017-9554-1. [Google Scholar]
- Palleroni, N.J. Genus I Pseudomonas. In Bergey’s Manual of Systematic Bacteriology; Krieg, N.R., Holt, J.C., Eds.; The Williams and Wilkins Co.: Baltimore, MD, USA, 1984; pp. 141–199. [Google Scholar]
- Stanier, R.Y.; Palleroni, N.J.; Doudoroff, M. The Aerobic Pseudomonads a Taxonomic Study. J. Gen. Microbiol. 1966, 43, 159–271. [Google Scholar] [CrossRef] [Green Version]
- Chin-A-Woeng, T.F.C.; Bloemberg, G.V.; Lugtenberg, B.J.J. Phenazines and Their Role in Biocontrol by Pseudomonas Bacteria. New Phytol. 2003, 157, 503–523. [Google Scholar] [CrossRef] [Green Version]
- Mavrodi, D.V.; Parejko, J.A.; Mavrodi, O.V.; Kwak, Y.-S.; Weller, D.M.; Blankenfeldt, W.; Thomashow, L.S. Recent Insights into the Diversity, Frequency and Ecological Roles of Phenazines in Fluorescent Pseudomonas spp. Environ. Microbiol. 2013, 15, 675–686. [Google Scholar] [CrossRef]
- Biessy, A.; Filion, M. Phenazines in Plant-Beneficial Pseudomonas spp.: Biosynthesis, Regulation, Function and Genomics. Environ. Microbiol. 2018, 20, 3905–3917. [Google Scholar] [CrossRef] [Green Version]
- Thomashow, L.S.; Weller, D.M. Role of a Phenazine Antibiotic from Pseudomonas Fluorescens in Biological Control of Gaeumannomyces Graminis Var. Tritici. J. Bacteriol. 1988, 170, 3499–3508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavrodi, D.V.; Blankenfeldt, W.; Thomashow, L.S. Phenazine Compounds in Fluorescent Pseudomonas spp. Biosynthesis and Regulation. Annu. Rev. Phytopathol. 2006, 44, 417–445. [Google Scholar] [CrossRef] [PubMed]
- Selin, C.; Habibian, R.; Poritsanos, N.; Athukorala, S.N.P.; Fernando, D.; De Kievit, T.R. Phenazines Are Not Essential for Pseudomonas Chlororaphis PA23 Biocontrol of Sclerotinia Sclerotiorum, but Do Play a Role in Biofilm Formation. FEMS Microbiol. Ecol. 2010, 71, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeTourneau, M.K.; Marshall, M.J.; Cliff, J.B.; Bonsall, R.F.; Dohnalkova, A.C.; Mavrodi, D.V.; Devi, S.I.; Mavrodi, O.V.; Harsh, J.B.; Weller, D.M.; et al. Phenazine-1-Carboxylic Acid and Soil Moisture Influence Biofilm Development and Turnover of Rhizobacterial Biomass on Wheat Root Surfaces. Environ. Microbiol. 2018, 20, 2178–2194. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wilks, J.C.; Danhorn, T.; Ramos, I.; Croal, L.; Newman, D.K. Phenazine-1-Carboxylic Acid Promotes Bacterial Biofilm Development via Ferrous Iron Acquisition. J. Bacteriol. 2011, 193, 3606–3617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeTourneau, M.; Marshall, M.J.; Grant, M.R.; Freeze, P.M.; Strawn, D.; Lai, B.; Dohnalkova, A.; Harsh, J.B.; Weller, D.M.; Thomashow, L.S. Phenazine-1-Carboxylic Acid-Producing Bacteria Enhance the Reactivity of Iron Minerals in Dryland and Irrigated Wheat Rhizospheres. Environ. Sci. Technol. 2019, 53, 14273–14284. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, M.; Cook, R.J.; Thomashow, L.S.; Weller, D.M.; Pierson, L.S. Contribution of Phenazine Antibiotic Biosynthesis to the Ecological Competence of Fluorescent Pseudomonads in Soil Habitats. Appl. Environ. Microbiol. 1992, 58, 2616–2624. [Google Scholar] [CrossRef] [Green Version]
- Mirleau, P.; Philippot, L.; Corberand, T.; Lemanceau, P. Involvement of Nitrate Reductase and Pyoverdine in Competitiveness of Pseudomonas Fluorescens Strain C7R12 in Soil. Appl. Environ. Microbiol. 2001, 67, 2627–2635. [Google Scholar] [CrossRef] [Green Version]
- Latour, X.; Delorme, S.; Mirleau, P.; Lemanceau, P. Identification of Traits Implicated in the Rhizosphere Competence of Fluorescent Pseudomonads: Description of a Strategy Based on Population and Model Strain Studies. Agronomie 2003, 23, 397–405. [Google Scholar] [CrossRef]
- Zboralski, A.; Filion, M. Genetic Factors Involved in Rhizosphere Colonization by Phytobeneficial Pseudomonas spp. Comput. Struct. Biotechnol. J. 2020, 18, 3539–3554. [Google Scholar] [CrossRef]
- Mazzola, M. Mechanisms of Natural Soil Suppressiveness to Soilborne Diseases. Antonie Van Leeuwenhoek 2002, 81, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, M.; Funnell, D.L.; Raaijmakers, J.M. Wheat Cultivar-Specific Selection of 2,4-Diacetylphloroglucinol-Producing Fluorescent Pseudomonas Species from Resident Soil Populations. Microb. Ecol. 2004, 48, 338–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Fuente, L.; Landa, B.B.; Weller, D.M. Host Crop Affects Rhizosphere Colonization and Competitiveness of 2, 4-Diacetylphloroglucinol-Producing Pseudomonas Fluorescens. Phytopathology 2006, 96, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.P.; Handelsman, J.; Goodman, R.M. Genetic Basis in Plants for Interactions with Disease-Suppressive Bacteria. Proc. Natl. Acad. Sci. USA 1999, 96, 4786–4790. [Google Scholar] [CrossRef] [Green Version]
- Persello-Cartieaux, F.; David, P.; Sarrobert, C.; Thibaud, M.-C.; Achouak, W.; Robaglia, C.; Nussaume, L. Utilization of Mutants to Analyze the Interaction between Arabidopsis Thaliana and Its Naturally Root-Associated Pseudomonas. Planta 2001, 212, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Zboralski, A.; Biessy, A.; Savoie, M.-C.; Novinscak, A.; Filion, M. Metabolic and Genomic Traits of Phytobeneficial Phenazine-Producing Pseudomonas spp. Are Linked to Rhizosphere Colonization in Arabidopsis Thaliana and Solanum Tuberosum. Appl. Environ. Microbiol. 2020, 86, e02443-19. [Google Scholar] [CrossRef]
- Biessy, A.; Novinscak, A.; St-Onge, R.; Léger, G.; Zboralski, A.; Filion, M. Inhibition of Three Potato Pathogens by Phenazine-Producing Pseudomonas spp. Is Associated with Multiple Biocontrol-Related Traits. mSphere 2021, 6, e00427-21. [Google Scholar] [CrossRef]
- Ghirardi, S.; Dessaint, F.; Mazurier, S.; Corberand, T.; Raaijmakers, J.M.; Meyer, J.-M.; Dessaux, Y.; Lemanceau, P. Identification of Traits Shared by Rhizosphere-Competent Strains of Fluorescent Pseudomonads. Microb. Ecol. 2012, 64, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Kamou, N.N.; Cazorla, F.; Kandylas, G.; Lagopodi, A.L. Induction of Defense-Related Genes in Tomato Plants after Treatments with the Biocontrol Agents Pseudomonas Chlororaphis ToZa7 and Clonostachys Rosea IK726. Arch. Microbiol. 2019, 202, 257–267. [Google Scholar] [CrossRef]
- Kamou, N.N.; Karasali, H.; Menexes, G.; Kasiotis, K.M.; Bon, M.C.; Papadakis, E.N.; Tzelepis, G.D.; Lotos, L.; Lagopodi, A.L. Isolation Screening and Characterisation of Local Beneficial Rhizobacteria Based upon Their Ability to Suppress the Growth of Fusarium Oxysporum f. Sp. Radicis-Lycopersici and Tomato Foot and Root Rot. Biocontrol Sci. Technol. 2015, 25, 928–949. [Google Scholar] [CrossRef]
- Landa, B.B.; Cachinero-Díaz, J.M.; Lemanceau, P.; Jiménez-Díaz, R.M.; Alabouvette, C. Effect of Fusaric Acid and Phytoanticipins on Growth of Rhizobacteria and Fusarium Oxysporum. Can. J. Microbiol. 2002, 48, 971–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okushima, Y. Functional Genomic Analysis of the AUXIN RESPONSE FACTOR Gene Family Members in Arabidopsis Thaliana: Unique and Overlapping Functions of ARF7 and ARF19. Plant Cell 2005, 17, 444–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Tsuda, K.; Sato, M.; Cohen, J.D.; Katagiri, F.; Glazebrook, J. Arabidopsis CaM Binding Protein CBP60g Contributes to MAMP-Induced SA Accumulation and Is Involved in Disease Resistance against Pseudomonas Syringae. PLoS Pathog. 2009, 5, e1000301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willemsen, V.; Bauch, M.; Bennett, T.; Campilho, A.; Wolkenfelt, H.; Xu, J.; Haseloff, J.; Scheres, B. The NAC Domain Transcription Factors FEZ and SOMBRERO Control the Orientation of Cell Division Plane in Arabidopsis Root Stem Cells. Dev. Cell. 2008, 15, 913–922. [Google Scholar] [CrossRef] [Green Version]
- Bartsch, M. Salicylic Acid-Independent ENHANCED DISEASE SUSCEPTIBILITY1 Signaling in Arabidopsis Immunity and Cell Death Is Regulated by the Monooxygenase FMO1 and the Nudix Hydrolase NUDT7. Plant Cell 2006, 18, 1038–1051. [Google Scholar] [CrossRef] [Green Version]
- Alonso, J.M. Genome-Wide Insertional Mutagenesis of Arabidopsis Thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.-P.; Tunc-Ozdemir, M.; Chang, Y.; Jones, A.M. Cooperative Control between AtRGS1 and AtHXK1 in a WD40-Repeat Protein Pathway in Arabidopsis Thaliana. Front. Plant Sci. 2015, 6, 851. [Google Scholar] [CrossRef] [Green Version]
- Vellosillo, T.; Martinez, M.; Lopez, M.A.; Vicente, J.; Cascon, T.; Dolan, L.; Hamberg, M.; Castresana, C. Oxylipins Produced by the 9-Lipoxygenase Pathway in Arabidopsis Regulate Lateral Root Development and Defense Responses through a Specific Signaling Cascade. Plant Cell 2007, 19, 831–846. [Google Scholar] [CrossRef] [Green Version]
- Kidd, B.N.; Edgar, C.I.; Kumar, K.K.; Aitken, E.A.; Schenk, P.M.; Manners, J.M.; Kazan, K. The Mediator Complex Subunit PFT1 Is a Key Regulator of Jasmonate-Dependent Defense in Arabidopsis. Plant Cell 2009, 21, 2237–2252. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Ellendorff, U.; Kemp, B.; Mansfield, J.W.; Forsyth, A.; Mitchell, K.; Bastas, K.; Liu, C.-M.; Woods-Tor, A.; Zipfel, C.; et al. A Genome-Wide Functional Investigation into the Roles of Receptor-like Proteins in Arabidopsis. Plant Physiol. 2008, 147, 503–517. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, S.; Ishii, T.; Matsunaga, T.; Tominaga, R.; Kuromori, T.; Wada, T.; Shinozaki, K.; Hirayama, T. The Glycerophosphoryl Diester Phosphodiesterase-like Proteins SHV3 and Its Homologs Play Important Roles in Cell Wall Organization. Plant Cell Physiol. 2008, 49, 1522–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, T.; van den Toorn, A.; Sanchez-Perez, G.F.; Campilho, A.; Willemsen, V.; Snel, B.; Scheres, B. SOMBRERO, BEARSKIN1, and BEARSKIN2 Regulate Root Cap Maturation in Arabidopsis. Plant Cell 2010, 22, 640–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, S. Arabidopsis TEBICHI, with Helicase and DNA Polymerase Domains, Is Required for Regulated Cell Division and Differentiation in Meristems. Plant Cell 2006, 18, 879–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deprost, D.; Yao, L.; Sormani, R.; Moreau, M.; Leterreux, G.; Nicolaï, M.; Bedu, M.; Robaglia, C.; Meyer, C. The Arabidopsis TOR Kinase Links Plant Growth, Yield, Stress Resistance and MRNA Translation. EMBO Rep. 2007, 8, 864–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional Regulation of ROS Controls Transition from Proliferation to Differentiation in the Root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Brader, G.; Kariola, T.; Tapio Palva, E. WRKY70 Modulates the Selection of Signaling Pathways in Plant Defense. Plant J. 2006, 46, 477–491. [Google Scholar] [CrossRef]
- Ojangu, E.-L.; Järve, K.; Paves, H.; Truve, E. Arabidopsis Thaliana Myosin XIK Is Involved in Root Hair as Well as Trichome Morphogenesis on Stems and Leaves. Protoplasma 2007, 230, 193–202. [Google Scholar] [CrossRef]
- Peremyslov, V.V.; Prokhnevsky, A.I.; Avisar, D.; Dolja, V.V. Two Class XI Myosins Function in Organelle Trafficking and Root Hair Development in Arabidopsis. Plant Physiol. 2008, 146, 1109–1116. [Google Scholar] [CrossRef] [Green Version]
- Springer, N.M. Isolation of Plant DNA for PCR and Genotyping Using Organic Extraction and CTAB. Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5515. [Google Scholar] [CrossRef]
- O’Malley, R.C.; Barragan, C.C.; Ecker, J.R. A User’s Guide to the Arabidopsis T-DNA Insertion Mutant Collections. In Plant Functional Genomics: Methods and Protocols; Alonso, J.M., Stepanova, A.N., Eds.; Humana Press: New York, NY, USA, 2015; pp. 323–342. [Google Scholar]
- King, E.O.; Ward, M.K.; Raney, D.E. Two Simple Media for the Demonstration of Pyocyanin and Fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Manefield, M.; Whiteley, A.S.; Bailey, M.J. DNA and RNA Extraction from Soil. In Molecular Microbial Ecology Manual; Kowalchuk, G.A., de Bruijn, F.J., Head, I.M., Akkermans, A.D.L., van Elsas, J.D., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 1, pp. 149–158. [Google Scholar]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.2-8; 2017; Available online: https://cran.r-project.org/web/packages/agricolae/index.html (accessed on 27 March 2020).
- Gao, X.; Alvo, M.; Chen, J.; Li, G. Nonparametric Multiple Comparison Procedures for Unbalanced One-Way Factorial Designs. J. Stat. Plan. Inference 2008, 138, 2574–2591. [Google Scholar] [CrossRef]
- Konietschke, F.; Placzek, M.; Schaarschmidt, F.; Hothorn, L.A. Nparcomp: An R Software Package for Nonparametric Multiple Comparisons and Simultaneous Confidence Intervals. J. Stat. Softw. 2015, 64, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Hawes, M.; Allen, C.; Turgeon, B.G.; Curlango-Rivera, G.; Minh Tran, T.; Huskey, D.A.; Xiong, Z. Root Border Cells and Their Role in Plant Defense. Annu. Rev. Phytopathol. 2016, 54, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Haldar, S.; Sengupta, S. Plant-Microbe Cross-Talk in the Rhizosphere: Insight and Biotechnological Potential. Open Microbiol. J. 2015, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Keunen, E.; Remans, T.; Opdenakker, K.; Jozefczak, M.; Gielen, H.; Guisez, Y.; Vangronsveld, J.; Cuypers, A. A Mutant of the Arabidopsis Thaliana LIPOXYGENASE1 Gene Shows Altered Signalling and Oxidative Stress Related Responses after Cadmium Exposure. Plant Physiol. Biochem. 2013, 63, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Noirot-Gros, M.-F.; Shinde, S.; Larsen, P.E.; Zerbs, S.; Korajczyk, P.J.; Kemner, K.M.; Noirot, P.H. Dynamics of Aspen Roots Colonization by Pseudomonads Reveals Strain-Specific and Mycorrhizal-Specific Patterns of Biofilm Formation. Front. Microbiol. 2018, 9, 853. [Google Scholar] [CrossRef] [PubMed]
- Pliego, C.; de Weert, S.; Lamers, G.; de Vicente, A.; Bloemberg, G.; Cazorla, F.M.; Ramos, C. Two Similar Enhanced Root-Colonizing Pseudomonas Strains Differ Largely in Their Colonization Strategies of Avocado Roots and Rosellinia Necatrix Hyphae. Environ. Microbiol. 2008, 10, 3295–3304. [Google Scholar] [CrossRef]
- Carvalhais, L.C.; Schenk, P.M.; Dennis, P.G. Jasmonic Acid Signalling and the Plant Holobiont. Curr. Opin. Microbiol. 2017, 37, 42–47. [Google Scholar] [CrossRef]
- Biessy, A.; Novinscak, A.; Blom, J.; Léger, G.; Thomashow, L.S.; Cazorla, F.M.; Jošić, D.; Filion, M. Diversity of Phytobeneficial Traits Revealed by Whole-genome Analysis of Worldwide-isolated Phenazine-producing Pseudomonas spp. Environ. Microbiol. 2019, 21, 437–455. [Google Scholar] [CrossRef] [Green Version]
- AbuQamar, S.; Chen, X.; Dhawan, R.; Bluhm, B.; Salmeron, J.; Lam, S.; Dietrich, R.A.; Mengiste, T. Expression Profiling and Mutant Analysis Reveals Complex Regulatory Networks Involved in Arabidopsis Response to Botrytis Infection. Plant J. 2006, 48, 28–44. [Google Scholar] [CrossRef]
- Knoth, C.; Ringler, J.; Dangl, J.L.; Eulgem, T. Arabidopsis WRKY70 Is Required for Full RPP4 -Mediated Disease Resistance and Basal Defense against Hyaloperonospora Parasitica. Mol. Plant Microbe Interact. 2007, 20, 120–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Mortel, J.E.; de Vos, R.C.H.; Dekkers, E.; Pineda, A.; Guillod, L.; Bouwmeester, K.; van Loon, J.J.A.; Dicke, M.; Raaijmakers, J.M. Metabolic and Transcriptomic Changes Induced in Arabidopsis by the Rhizobacterium Pseudomonas Fluorescens SS101. Plant Physiol. 2012, 160, 2173–2188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.-H.; Huang, Z.-Y.; Xie, P.; Gu, C.; Li, K.; Wang, D.-C.; Yu, Y.-Y.; Fan, Z.-H.; Wang, C.-J.; Wang, Y.-P.; et al. Transcription Factors WRKY70 and WRKY11 Served as Regulators in Rhizobacterium Bacillus Cereus AR156-Induced Systemic Resistance to Pseudomonas Syringae Pv. Tomato DC3000 in Arabidopsis. J. Exp. Bot. 2016, 67, 157–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.; Zhang, Y.; Li, Y.; Zhang, Q.; Ding, Y.; Zhang, Y. ChIP-Seq Reveals Broad Roles of SARD1 and CBP60g in Regulating Plant Immunity. Nat. Commun. 2015, 6, 10159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundaravelpandian, K.; Chandrika, N.N.P.; Tsai, Y.-H.; Schmidt, W. PFT1-Controlled ROS Balance Is Critical for Multiple Stages of Root Hair Development in Arabidopsis. Plant Signal. Behav. 2013, 8, e24066. [Google Scholar] [CrossRef]
- Zhang, W.; Fraiture, M.; Kolb, D.; Löffelhardt, B.; Desaki, Y.; Boutrot, F.F.G.; Tör, M.; Zipfel, C.; Gust, A.A.; Brunner, F. Arabidopsis RECEPTOR-LIKE PROTEIN30 and Receptor-like Kinase SUPPRESSOR OF BIR1-1/EVERSHED Mediate Innate Immunity to Necrotrophic Fungi. Plant Cell 2013, 25, 4227–4241. [Google Scholar] [CrossRef] [Green Version]
- Nisa, M.; Bergis, C.; Pedroza-Garcia, J.; Drouin-Wahbi, J.; Mazubert, C.; Bergounioux, C.; Benhamed, M.; Raynaud, C. The Plant DNA Polymerase Theta Is Essential for the Repair of Replication-associated DNA Damage. Plant J. 2021, 106, 1197–1207. [Google Scholar] [CrossRef]

| Affected Gene | Related Protein | Phenotypic Impacts of the Mutation | Accession Number | Reference |
|---|---|---|---|---|
| wild type | - | - | CS70000 (Col-0) | - |
| arf7 | AUXIN RESPONSE FACTOR 7 | Impaired phototropic and gravitropic response in hypocotyls, reduced numbers of lateral roots, epinastic rosette leaves, reduced auxin sensitivity in hypocotyl growth | CS24607 | [59] |
| cbp60g | CAM-BINDING PROTEIN 60-LIKE G | Enhanced susceptibility to P. syringae | SALK_023199C | [60] |
| fez | FEZ | Reduced number of cell layers in the root caps | SALK_025663C | [61] |
| fmo1 | FLAVIN-DEPENDENT MONOOXYGENASE 1 | Increased susceptibility to virulent P. syringae infection | SALK_026163C | [62] |
| hda5 | HISTONE DEACETYLASE 5 | Increased root hair density | SALK_093312C | [63] |
| hxk1 | HEXOKINASE 1 | Reduced growth in roots, leaf and stem length, rosette size and inflorescence | CS69135 | [64] |
| lox1 | LIPOXYGENASE 1 | Moderate increase in the length of the primary root and increased number of emergent and lateral roots | SALK_059431C | [65] |
| lox5 | LIPOXYGENASE 5 | Moderate increase in the length of the primary root and increased number of emergent and lateral roots | SALK_044826C | [65] |
| mrp2 | MULTIDRUG RESISTANCE- ASSOCIATED PROTEIN 2 | Changes in root exudates composition and shorter primary root | CS66052 | [21] |
| pft1 | PHYTOCHROME AND FLOWERING TIME 1 | Enhanced susceptibility to leaf infecting necrotrophic pathogens | SALK_129555C | [66] |
| pgp1 | P-GLYCO- PROTEIN 1 | Changes in root exudates composition and increased lateral root formation | CS66051 | [21] |
| rhip1 | RGS1-HXK1 INTERACTING PROTEIN 1 | Longer roots in young seedlings and larger inflorescence | CS69137 | [64] |
| rlp30 | RECEPTOR LIKE PROTEIN 30 | Enhanced susceptibility to bacterial and fungal pathogens | CS65465 | [67] |
| shv3 | SHAVEN 3 | No tip growth in almost all root hair cells | SALK_024208C | [68] |
| smb | SOMBRERO | Additional cell layers in the root caps | SALK_143526C | [61,69] |
| teb | TEBICHI | Short-root phenotype and reduced size of the aerial system, with highly serrated and asymmetric leaves and a fasciated stem | SALK_018851C | [70] |
| tor | TARGET OF RAPAMYCIN | Larger plants, more resistant to osmotic stress | SALK_007846C | [71] |
| upb1 | UPBEAT 1 | Longer roots, increased number of cortex cell | CS868100 | [72] |
| wrky70 | WRKY DNA-BINDING PROTEIN 70 | Enhanced susceptibility to bacterial and fungal pathogens | SALK_025198C | [73] |
| xik | MYOSIN XI-K | Reduced length and altered shape in root hair | SALK_067972C | [74,75] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zboralski, A.; Saadia, H.; Novinscak, A.; Filion, M. Interplay between Arabidopsis thaliana Genotype, Plant Growth and Rhizosphere Colonization by Phytobeneficial Phenazine-Producing Pseudomonas chlororaphis. Microorganisms 2022, 10, 660. https://doi.org/10.3390/microorganisms10030660
Zboralski A, Saadia H, Novinscak A, Filion M. Interplay between Arabidopsis thaliana Genotype, Plant Growth and Rhizosphere Colonization by Phytobeneficial Phenazine-Producing Pseudomonas chlororaphis. Microorganisms. 2022; 10(3):660. https://doi.org/10.3390/microorganisms10030660
Chicago/Turabian StyleZboralski, Antoine, Hara Saadia, Amy Novinscak, and Martin Filion. 2022. "Interplay between Arabidopsis thaliana Genotype, Plant Growth and Rhizosphere Colonization by Phytobeneficial Phenazine-Producing Pseudomonas chlororaphis" Microorganisms 10, no. 3: 660. https://doi.org/10.3390/microorganisms10030660
APA StyleZboralski, A., Saadia, H., Novinscak, A., & Filion, M. (2022). Interplay between Arabidopsis thaliana Genotype, Plant Growth and Rhizosphere Colonization by Phytobeneficial Phenazine-Producing Pseudomonas chlororaphis. Microorganisms, 10(3), 660. https://doi.org/10.3390/microorganisms10030660