Abstract
Bacterial pathogens, such as Listeria monocytogenes, can show resistance to disinfection and persistence on working surfaces, permitting them to survive and contaminate food products. Persistence—a complex phenomenon involving interactions between many bacteria within a biofilm—is modulated by in situ characteristics. This study aimed to describe, in silico, the microbiota identified in a swine slaughterhouse after sanitation procedures to better understand the presence of L. monocytogenes on these surfaces. Molecular tools for characterization of microbial communities were used to assess the relative contribution of different bacteria resulting from this phenomenon, and the 16S rRNA sequencing method was used on samples from meat conveyor belt surfaces collected on four sampling visits to study the co-occurrence between L. monocytogenes and other bacteria. From the background microbiota, a total of six genera were found to be negatively correlated with Listeria spp., suggesting Listeria growth inhibition, competition, or at least an absence of shared habitats. Based on these results, a complete scenario of interactions of Listeria with components of background microbiota was established. This work contributes to identifying avenues that could prevent the growth and persistence of L. monocytogenes on food-processing surfaces.
1. Introduction
The presence of Listeria monocytogenes in food processing plants is a major concern for industries and food control authorities [1,2,3]. The ubiquitous nature of this bacterium combined with its ability to grow in harsh conditions, including high salt concentrations, high acidity levels, and large temperature ranges, makes its control within food production environments particularly challenging [4]. Indeed, the ability of L. monocytogenes to persist within food-processing environments was reported in many studies [4,5,6,7], and the circumstances associated with this persistence phenomenon appear to be complex [8]. L. monocytogenes’ biofilm formation and resistance to commonly used disinfectants in food industries may partly explain this persistence [5,6,7,9]. Some authors found an increased ability for biofilm formation in L. monocytogenes’ persistent strains. Moreover, the detection of genes conferring resistance to disinfectants, such as quaternary ammonium compounds (QACs), was correlated with persistence phenotypes in strains isolated from food processing plants [2,10,11]. Another factor that could influence the persistence of L. monocytogenes in the food industry is the structure and composition of bacterial communities present in biofilms, which are expected to include species other than L. monocytogenes [12,13]. As an example, it was reported that microbial communities dominated by Gram-negative bacterial species identified on a salmon slaughterhouse conveyor belt seemed to have an inhibitory effect on L. monocytogenes in a mixed biofilm [13]. This previous study was based on a culture-based method to identify communities present on conveyor surfaces. Consequently, only the most abundant and cultivable bacteria were detected, which limited the ability to explore links and/or interactions between L. monocytogenes and other bacteria present on biofilms from meat conveyor surfaces. While bacteria from specific species could influence the occurrence of other species for many reasons—such as competing for the same resources [14,15]—the identification of co-occurrences between L. monocytogenes and other species might help us find innovative solutions for controlling the proliferation and/or persistence of this pathogen.
So far, there is scant information about the co-occurrence of L. monocytogenes and other bacteria species over time in the food production environment. Studies that were conducted include one that showed L. monocytogenes was outcompeted by a mixture of bacterial species present on wooden shelves mainly due to competition for nutrients [16,17]. Inhibition of L. monocytogenes growth could also be the result of bacteriocin secretion by other species, such as Enterococcus faecium [18]. Other studies, in contrast, demonstrated positive interactions between L. monocytogenes and other bacterial genera [19,20]. Indeed, for example, Flavobacterium spp., which was part of the background microbiota of a seafood processing plant, was reported to enhance the colonization of L. monocytogenes on stainless steel surfaces [19]. Finally, others reported no interaction between L. monocytogenes and other bacteria such as Pseudomonas fluorescens [12,21].
The aim of this study was to describe, independent of the cultivability of the bacteria, (1) the microbiota diversity on meat conveyor surfaces from a swine slaughterhouse after cleaning and disinfection procedures and (2) the co-occurrence of Listeria spp. with other bacterial communities present on meat conveyor surfaces.
2. Materials and Methods
2.1. Sample Collection, Processing, and Culture of Listeria monocytogenes
Samples were collected in the cutting facility of one swine slaughterhouse during four visits and after cleaning and disinfection operations. The disinfection procedures were applied by industry employees as part of their routine activities and consisted of an application of QACs-based disinfectants at a concentration between 150 and 200 ppm, on a daily basis, after each meat-cutting process.
On each visit, the belt of three meat conveyors was sampled at the beginning, middle, and end of the conveyor. Each sampling consisted of a swabbing of 1 m2 of the belt surface after mechanical mobilization (brushing) using prewarmed wet swabs with a D/E (Dey-Engley) neutralizing broth (Innovation Diagnostic, Saint-Eustache, QC, Canada) to neutralize a broad spectrum of disinfectants and antiseptics. A total of 48 swab samples were collected. Each swab was shaken vigorously in 100 mL of sodium chloride solution (0.9% of NaCl in nuclease-free water). A volume of 10 mL was transferred into a separate tube and centrifuged, and the supernatants were discarded. The tubes containing pellets were immediately stored at −80 °C for DNA extraction. The detection of L. monocytogenes was performed on the 90 mL of remaining suspension according to the Compendium of Analytical Methods, MFHPB-30 [22] with few modifications. Briefly, all samples were enriched in 90 mL of 2X concentrated University of Vermont media 1 (UVM-1; Innovation Diagnostics, Saint-Eustache, QC, Canada) and incubated at 30 °C for 48 h. A second enrichment with Fraser broth media (Innovation Diagnostics, Saint-Eustache, QC, Canada) was used at 37 °C for 24 h. The cultures were plated on a selective COMPASS Listeria agar (Innovation Diagnostics, Saint-Eustache, QC, Canada) media, and typical colonies were streaked on sheep blood agar (Oxoid, Nepean, ON, Canada). The confirmation of Listeria monocytogenes was performed by a polymerase chain reaction (PCR) used for serogrouping [23]. In addition to these swab samples, one control, consisting of a clean swab and brush transported during the sampling visit, was collected on two sampling visits (1 and 4) and submitted to the same bacteriological and molecular analyses as the swab samples.
2.2. DNA Extraction, 16S rDNA Construction Library, and Bioinformatics Analysis
The conserved pellets from the step above were used for phenol chloroform DNA extraction protocol. Briefly, bacteria were lysed with a lysis buffer (Tris-Hcl, EDTA, NaCl, and SDS) and content was extracted using glass beads by vortexing using the FastPrep procedure (MP Biomedical, Solon, OH, USA). The phenol chloroform isoamylic (25:24:1 v:v) solution was added to the supernatant. After mixing by inversion for 2 min, the solution was centrifuged, and the pellets were discarded. An additional 2 min of mixing was conducted, and, finally, the solution was centrifuged to collect the supernatant. Ammonium acetate (10 mM) and cold ethanol 100% were added to the supernatant, and the suspension was kept at −80 °C for 24 h for DNA precipitation. After precipitation, the tubes were centrifuged at 21,004× g for 15 min, and the pellets were washed with ethanol before drying. The DNA was solubilized in 50 µL of nuclease-free water and stored at −20 °C before PCR amplification of the 16S rDNA V4 region. The primers 515F/806R pair [24] was used to amplify the V4 region, and the libraries were prepared using the NEXTERA kit (Illumina, Inc., San Diego, CA, USA) following recommendations from the manufacturer. The Illumina Miseq technology was used to perform the paired-end sequencing at Genome Québec Innovation Center (https://www.genomequebec.com/en/home/, accessed on 12 April 2016).
Data were analyzed using Quantitative Insights Into Microbial Ecology (QIIME) 2 v 2017.12 pipeline [25]. The demultiplexed reads were filtered and denoised using Dada2 v 2017.12.1 software [26]. The first 13 low-quality bases and the last 10 ones (at position 240) from both of the reverse (R1) and forward (F1) reads were trimmed during this step based on the quality plots that QIIME 2 generated. The resulting two separated tables containing amplicon single variants (ASVs) and sequences of each ASV were used for the downstream analyses.
ASVs table and their corresponding sequences were clustered into operational taxonomic units (OTU) using VSEARCH [27] v2018.8.0 and a closed-reference clustering with identity cutoff set at 99% against the SILVA database [28]. To minimize the impact of potential false positives due to cross-contamination, we used the decontam R package [29] to detect and remove potential OTUs detected as contaminants, using default settings.
Taxonomic composition of samples was determined by performing a pre-trained, naïve Bayes classifier on the reference taxonomy and sequences from SILVA database with 99% identity [28]. The feature classifier sklearn-classify [28] was used for taxonomic classification.
2.3. Diversity of OTUs by Sampling Visit and Listeria monocytogenes Culture Status
The alpha and beta diversities of OTUs were estimated for each sample rarefied at 20,000 sequences per sample. This threshold was defined based on the sample having the lowest number of sequences (n = 20,287) and was supported by the exploration of the rarefaction curves performed using phyloseq R package [30] v1.26.1, considering that this sample size was sufficient to reach the curve plateau for the majority of samples (Supplementary Figure S1 and Supplementary Table S1).
The alpha diversity, Shannon diversity index [31], and Shannon evenness index [32] were estimated in each rarefied sample. For the two indices, the median values of samples were compared between visits using a Kruskal–Wallis test followed by pairwise comparisons with Bonferroni adjustment for multiple testing. The median values were also compared according to the L. monocytogenes culture status of samples using a Wilcoxon test.
For the beta diversity, Bray–Curtis and Jaccard dissimilarity indices were estimated to explore the quantitative (community abundance) and qualitative (presence/absence) dissimilarities of OTUs in each sample, respectively, using phyloseq package in R. The Bray–Curtis and Jaccard indices were compared between visits and according to L. monocytogenes culture status using pairwise PERMANOVA analyses with 999 permutations.
2.4. Description of the Taxonomy Composition
To explore the taxonomy composition of the bacterial communities in OTUs from sampled conveyor surfaces, the relative abundance of their genera was described using the phyloseq package. All OTUs with missing values for the genus rank were excluded. The relative abundance of the most frequent genus in OTUs according to the sampling visits and to the L. monocytogenes culture status was also evaluated.
2.5. Description of Bacterial Communities
To explore the relationships between microbial communities, particularly between Listeria and other genera, a network was created from positive and/or negative correlations between bacterial genera. Rare genera were first filtered to remove all OTUs corresponding to the genus with fewer than 20 occurrences across all samples. The filtered table was then used to construct the network based on three indicators: Spearman correlation, Bray–Curtis, and Kullback–Leibler dissimilarities. This network was built using the CoNet program [33] and implemented with Cytoscape software v3.3.0 [34]. A threshold of 1500 top- and bottom-scoring (for anti-correlations) links was included for each of the three indicators. For each link between two genera, 100 renormalized permutations and bootstrap score distributions were computed. Brown’s method was used to merge all measure-specific p-values of the indicators [35], with false discovery rate controlled at 5% using the Benjamini–Hochberg correction. Only links with confirmed associations for all 3 indicators were kept in the final network. The resulting network was visualized in Gephi v 0.9.2 [36] using the Yifan Hu algorithm [37]. Nodes (i.e., genera with significant interactions) were clustered using the constant Potts model with the resolution set at 0.1 [38] to identify the community structure of the network, with the size of each node representing the relative abundance of the genus. A second network was created from the previous one limited to the nodes directly or indirectly connected to Listeria spp. For better clarity of the visualization, the network was filtered based on the Spearman correlation coefficient used in the analysis as the weight of the link. Thus, only nodes with a correlation coefficient higher than 0.45 were kept.
3. Results
L. monocytogenes was detected by a culture-based method in 13 (27.1%) out of the 48 samples. The first and second visits showed the highest proportion of positive samples (Table 1).

Table 1.
Bacteriological detection of L. monocytogenes in samples from swine slaughterhouse conveyor surfaces collected after cleaning and sanitation, by sampling visit.
From the 48 samples, three samples were not sequenced due to the small amount of extracted DNA. The number of sequences in the remaining 45 samples after quality control procedures using Dada2 ranged from 20,287 to 229,410 sequences per sample (Supplementary Table S1). All downstream analyses were performed based on the metadata table shown in Supplementary Table S1.
3.1. Diversity of the Microbiota on Conveyor Surfaces Was Different between Visits
Shannon diversity and evenness indices showed a significant difference of both richness and evenness in OTUs (Kruskal–Wallis, p = 0.00005 and p = 0.00009, respectively) between sampling visits. Samples from visit S1 were less diverse compared to all other sampling visits (Kruskal–Wallis, p < 0.05, Figure 1a). Moreover, a significantly higher diversity and higher evenness were observed in S2 compared to S4 (Figure 1a). Alpha diversity comparisons between culture-positive and culture-negative samples for L. monocytogenes showed no significant differences (Figure 1b).
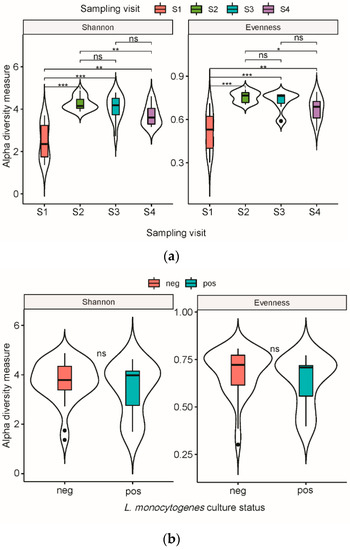
Figure 1.
Violin box plot representation of alpha diversity of bacterial OTUs from meat conveyor surfaces in a cutting facility from a swine slaughterhouse. (a) Diversity was measured by Shannon diversity index and Shannon evenness index according to sampling visit, with medians compared using the Kruskal–Walis test. (b) Diversity was measured by Shannon diversity index and Shannon evenness index according to Listeria monocytogenes culture status, with medians compared using the Wilcoxon test. Statistical significance: n.s., p > 0.05 *, p < 0.05 **, p < 0.01 ***, p < 0.001.
When richness estimations were compared between visits by pairwise PERMANOVA analysis based on Bray–Curtis and Jaccard indices, the diversities were significantly different between all pairs of visits (pseudo-F test, p = 0.001 for both indices), except between S2 and S3 (Table 2; Figure 2a,b). Diversity comparisons between the group of samples positive and negative for L. monocytogenes showed no difference (Figure 2c,d).
Table 2.
Pairwise PERMANOVA analysis based on Bray–Curtis and analysis of variance using Jaccard distance matrices with 999 permutations.
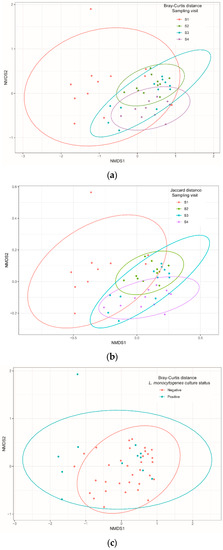
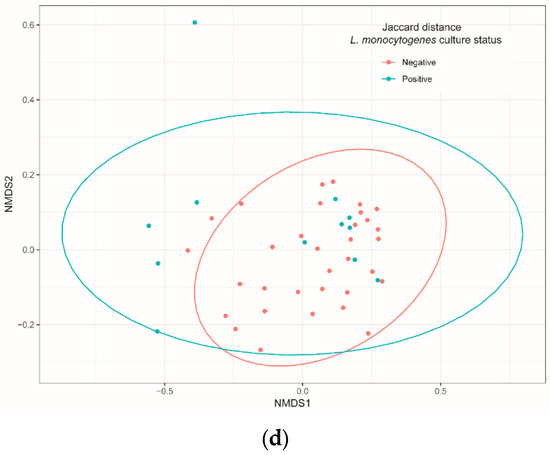
Figure 2.
Non-metric multidimensional scaling (NMDS) of Bray–Curtis and Jaccard distances of bacteria communities identified using 16S Miseq sequencing technology on samples isolated from cutting facility conveyor surfaces according to sampling visit (a,b) and Listeria monocytogenes culture status (c,d). Pairwise PERMANOVA with 999 permutations (using pseudo-F ratios) was used for all comparisons.
3.2. Relative Abundance and Microbial Composition Changed between Visits
A total of 114 different genera were obtained from OTUs. Overall, 98.8% of the OTUs were from the five most abundant phyla, i.e., Proteobacteria, Actinobacteria, Bacteroidetes, Firmicutes, and Acidobacteria. The most frequent phylum was Proteobacteria with a relative abundance of 73.2%, followed by Bacteroidetes (12.4%), Actinobacteria (9.9%), and Firmicutes (2.7%). Pseudomonas, Sphingomonas, Acinetobacter, and Chryseobacterium were the most abundant genera identified. Proportions of OTUs belonging to these genera changed between sampling visits. Pseudomonas was highly dominant in S1, whereas the dominant genus was Acinetobacter in S2 and S3 and Sphingomonas in S4 (Figure 3a). Listeria appeared among the least abundant genera; thus, it was identified almost exclusively in S1 and S2, which were also the visits with the highest proportion of culture-positive L. monocytogenes samples. Results showed that Pseudomonas was the most abundant genus in OTUs from L. monocytogenes culture-positive samples, whereas Sphingomonas was the most abundant genus in the culture-negative samples (Figure 3b).
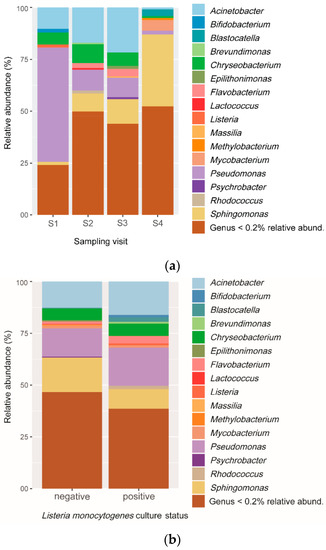
Figure 3.
Relative frequency of different taxonomic profiles identified from OTUs in cutting facility conveyor surfaces. (a) Relative abundance of the top sixteen genera according to the sampling periods. (b) Relative abundance of the top sixteen genera according to L. monocytogenes culture results.
3.3. Differential Community Interaction
Our results showed the presence of correlations forming 1027 links and involving 90 nodes representing the genera before applying the threshold of 0.45 for the Spearman correlation coefficient (Figure 4a and Supplementary Table S2). A total of 46 genera interacted with Listeria negatively and interestingly; Pseudomonas was observed to interact negatively with the genus Williamsia, and this latter had a direct link with Listeria (Figure 4a).
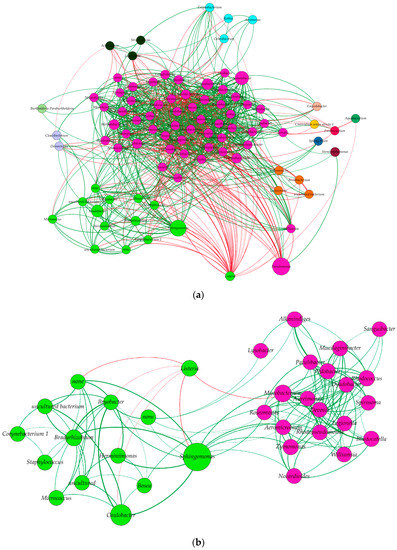
Figure 4.
Network construction based on Spearman correlation, Bray–Curtis dissimilarities, and Kullback–Leibler dissimilarities. (a) General network of all associations between genera clustered according to constant Potts model (communities sharing most links between each other). The size of each node refers to the abundance of the genus, the thickness of the link refers to its weight (presented here by the Spearman correlation coefficient), and the color of each node refers to positive (green) and negative interactions (red), (b) Network construction based on the direct associations between Listeria genus and the other genera and indirect associations between those genera and others. Links colored in red represent negative associations between the genera while green links represent positive associations. The node labelled “none” refers to the unidentified genus from the Elusimicrobia phylum.
When a threshold of ≥0.45 for the Spearman correlation was applied from the previous step and the network was filtered to keep only genera interacting directly or indirectly with Listeria genus, one community was found to interact with Listeria community, which was composed of 13 other genera, covering 21.1% of the nodes (Figure 4b). According to our results, the Listeria genus had no direct positive correlation with any other bacterial genera (Figure 4b), but direct negative correlations were present between Listeria and the six genera, Herminiimonas, Bryobacter, Caulobacter, None (genus not identified), Sphingomonas, and Mycobacterium, belonging to the six phyla, Proteobacteria, Acidobacteria, Proteobacteria, Elusimicrobia, Proteobacteria, and Actinobacteria, respectively (Figure 4b). Interestingly, Proteobacteria and Actinobacteria were the phyla which had the highest numbers of direct associations with Listeria, while Acidobacteria, Bacteroidetes, and Elucimicrobia had only two, one, and one genus, respectively, interacting with Listeria. From the Proteobacteria phylum, the Sphyngomonas genus accounted for the most abundant genera interacting negatively with Listeria (Figure 4b).
4. Discussion
In this study, we described the diversity of the microbiota present on swine meat conveyor surfaces from four sampling visits. We also identified, in silico, the co-occurrence of this microbiota with the Listeria genus, including Listeria monocytogenes, as confirmed by a culture-based method.
The proportion of culture-positive samples for L. monocytogenes observed in this study was in the same range as reported in other studies [4]. The relative abundance of Listeria spp. revealed by 16S metagenomic in culture-positive samples for L. monocytogenes was higher than in culture-negative samples. This concordance between the two methods may suggest that L. monocytogenes was the most prevalent species among Listeria spp. present on conveyor surfaces. Moreover, classification at the species level only identified L. monocytogenes among the Listeria genus (Supplementary Figure S2).
Diversity analyses showed significant differences in both composition and organization of microbial communities depending on the sampling visits. Continuous new sources of bacteria from incoming materials or persons entering the processing environment should be considered in interpreting such diversity. It was reported, for example, that carcasses contaminated by L. monocytogenes from internal organs, such as tonsils and tongue, during the slaughtering process may contaminate plant environments [39,40]. Additionally, the plant workers can be an important source of contamination of food processing plant environments and products since humans are known to be an important reservoir, and the risk of contamination is associated with the level of sanitary practices [41].
From one batch to another, the microbiota composition changed, and these modifications were particularly noticeable due to the long intervals of time in our study (i.e., from three weeks to eight months between two consecutive visits). In addition, fluctuations in humidity, temperature, nutrient access, shear forces, and other physicochemical stresses could affect microbiota composition on conveyor surfaces over time, and one may think that only bacteria that can withstand these fluctuations could establish a resident niche, as supported by Fagerlund, Møretrø, Heir, Briandet, and Langsrud [12].
Overall, there was no significant difference in diversity indices of background microbiota between the culture-positive and culture-negative samples and L. monocytogenes. In our conditions, the presence of L. monocytogenes was not affected by the variability (relative abundance of species) and variety (number of different species) of species composing the microbiota of the meat conveyor surfaces.
According to taxonomic classification, Proteobacteria was the most abundant phylum, followed by Bacteroidetes, Actinobacteria, Firmicutes, and Acidobacteria. Few data are available to confirm these distributions. Despite most phyla being the same as observed in another study, Actinobacteria was reported as one of the most abundant phyla detected in meat samples [42]. This discrepancy could be due to their sampling period, which was before cleaning and sanitation on meat conveyor surfaces, whereas our sampling was done after cleaning and sanitation. It was previously reported that such factors may affect the composition of the bacterial community [12].
Among the 114 identified genera, the Gram-negative Sphingomonas, Pseudomonas, Acinetobacter, Chryseobacterium, and Caulobacter were the most abundant ones across all sampling visits. These genera have already been pointed out as the most abundant background microbiota of meat conveyor surfaces [12], as well as of processing plant equipment in fish production [2,12,13]. Furthermore, Pseudomonas and Acinetobacter were reported previously for their ability to survive cleaning and their high biofilm-forming ability [12], which may help to explain their high abundance. The Sphingomonas genus was not detected when culture-based approaches were used, while sequence-based, cultivation-independent approaches found this genus as the most abundant, with Pseudomonas spp. [43]. The failure to detect Sphingomonas with a culture-based method may be the result of higher requirements in growth factors, such as Mg2+ for some Sphingomonas, as suggested by some researchers [44], and/or to interspecies competition [43]. Our findings confirmed this genus forms a significant part of the microbial communities and underlined the complementarity of both methods for description of bacterial composition in a given environment.
Listeria and Staphylococcus were among the most abundant genera in the Firmicutes phylum detected on these cleaned meat conveyor surfaces. Considering previous studies describing Listeria (and particularly L. innocua and L. monocytogenes) as a weak competitor in presence of Pseudomonas and Acinetobacter [12,13], it was expected that this genus would be less abundant in our communities, particularly when both Pseudomonas and Acinetobacter were well represented. The co-occurrence of Listeria and Pseudomonas genera reported in this study has to then be considered in accordance with other studies where an increase in the number of cells of L. monocytogenes was reported with the growth of Pseudomonas putida or P. fragi biofilms [12,45]. Moreover, when Pseudomonas and Listeria were grown in a mixed biofilm with other genera, it was shown that Pseudomonas dominated but did not eliminate Listeria from the biofilm [13]. Our results suggest that Listeria formed niches within Pseudomonas biofilm and could withstand cleaning and disinfection procedures and other harsh conditions. All the aforementioned statements were, however, based on culture-based methods, which are limited for identifying several bacteria from a given sample. Indeed, the strong competition that occurs during enrichment steps in cultures, as well as the viable but non-cultivable properties of some bacterial species on surfaces, underlines the need for non-culture-based analysis. For this reason, we aimed to study these interactions considering all bacterial populations using the 16S rDNA sequence-based methods. It is worth noting that these methods cannot distinguish between dead and viable bacteria. Thus, once possible associations are identified, the interactions might need to be confirmed, e.g., via cultivation-based methods.
At a community network level, considering all bacterial populations and based on correlation and dissimilar methods, only negative correlations between Listeria and other genera were detected. Such results were previously reported where, among twenty-nine bacterial species recovered from a dairy environment, sixteen induced reduction of biofilm formation by L. monocytogenes [46]. Such findings suggest that these populations are not mutualistic; either they do not share similar niches or a competition exists, as reported previously [47]. The competition between species could be related to nutrient limitation and augmented by antimicrobial compound production, as reported by Giaouris et al. [9]. Negative associations were often reported with Listeria spp., particularly for L. monocytogenes, since its presence, particularly in biofilm co-culture, was repeatedly hampered [13,48,49]. Listeria monocytogenes is recognized as a weak competitor when placed in a co-culture [2,12,13], which is consistent with our results. It is noteworthy that, except for Sphingomonas, these bacteria have never been cultivated along with Listeria spp. The low abundance of these genera compared to Pseudomonas and Sphingomonas and the difficulty of cultivation or non-cultivable status of some of these genera, such as certain species of Herminiimonas [50] and Pseudoclavibacter [51], could be the reasons why they were not detected in classical bacteriology studies. The negative association between Listeria and Sphingomonas is supported by a previous study where Sphingomonas inhibited the growth of L. monocytogenes by production of astaxanthin [52]. Interestingly, species from the Paracoccus genus, such as P. marcusii, which have a negative interaction with Listeria genus, are reported to produce astaxanthin as well [53,54]. Similar, negative associations with the Listeria genus or L. monocytogenes were observed with other species such as Lactococcus lactis [48].
5. Conclusions
In summary, this study showed significant differences in composition and abundances of residual microbiota on pork meat conveyor surfaces over time, which highlights the non-stable bacterial community composition in the meat industry environment. However, the core bacteria population, represented by the most abundant genera, was often the same over time in this environment. From this background microbiota, a total of six genera, such as Sphingomonas, were found to interact negatively with Listeria spp., which could be explained by various factors, such as growth inhibition, absence of shared habitats, or competition, but further studies based on cultured methods are needed to interpret these negative interactions. These results provide interesting information regarding the relationship between background microbiota and Listeria spp. in order to identify potential bacterial species inhibiting its persistence within the food production environment.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/microorganisms10030613/s1, Supplementary Table S1: Sample identification with number of sequences, OTUs, Listeria monocytogenes culture-base status, and Shannon measure and its evenness. Supplementary Table S2: Total number of nodes and links resulting from the network construction based on the Spearman correlation, Bray–Curtis, and Kullback–Leibler dissimilarities. Supplementary Figure S1: Rarefaction curve of OTUs for each sample collected from meat conveyor surfaces. Supplementary Figure S2: Relative frequency of bacterial species identified from OTUs detected in cutting facility conveyor surfaces according to positive samples in Listeria monocytogenes culture-based method.
Author Contributions
Conceptualization, T.C., J.A., S.Q. and P.F.; Data curation, T.C. and J.A.; Formal analysis, T.C.; Funding acquisition, P.F.; Methodology, T.C. and J.A.; Project administration, S.Q. and P.F.; Resources, S.Q. and P.F.; Software, T.C. and J.A.; Supervision, S.Q.; Validation, T.C., J.A., S.Q. and P.F.; Visualization, T.C. and J.A.; Writing—original draft, T.C.; Writing—review and editing, T.C., J.A., S.Q. and P.F. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Natural Sciences and Engineering Research Council of Canada (NSERC) RDCPJ 520873-17. The funding organizations did not have any influence either in the design of the study, in the analysis and interpretation of data, or in the writing of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Publicly available datasets were analyzed in this study. These data can be found here: PRJNA728720.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sofos, J.N. Challenges to meat safety in the 21st century. Meat Sci. 2008, 78, 3–13. [Google Scholar] [CrossRef]
- Cherifi, T.; Carrillo, C.; Lambert, D.; Miniaï, I.; Quessy, S.; Larivière-Gauthier, G.; Blais, B.; Fravalo, P. Genomic characterization of Listeria monocytogenes isolates reveals that their persistence in a pig slaughterhouse is linked to the presence of benzalkonium chloride resistance genes. BMC Microbiol. 2018, 18, 220. [Google Scholar] [CrossRef] [PubMed]
- Hadjicharalambous, C.; Grispoldi, L.; Goga, B.C. Quantitative risk assessment of Listeria monocytogenes in a traditional RTE product. EFSA J. 2019, 17, e170906. [Google Scholar] [PubMed]
- Ortiz, S.; Lopez, V.; Villatoro, D.; Lopez, P.; Davila, J.C.; Martinez-Suarez, J.N. A 3-year surveillance of the genetic diversity and persistence of Listeria monocytogenes in an Iberian pig slaughterhouse and processing plant. Foodborne Pathog. Dis. 2010, 7, 1177–1184. [Google Scholar] [CrossRef]
- Moretro, T.; Schirmer, B.C.T.; Heir, E.; Fagerlund, A.; Hjemli, P.; Langsrud, S. Tolerance to quaternary ammonium compound disinfectants may enhance growth of Listeria monocytogenes in the food industry. Int. J. Food Microbiol. 2017, 241, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Suárez, J.V.; Ortiz, S.; López-Alonso, V. Potential Impact of the Resistance to Quaternary Ammonium Disinfectants on the Persistence of Listeria monocytogenes in Food Processing Environments. Front. Microbiol. 2016, 7, 638. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.M.; Leonard, N.; Jordan, K. Physiological and transcriptional characterization of persistent and nonpersistent Listeria monocytogenes isolates. Appl. Environ. Microbiol. 2011, 77, 6559–6569. [Google Scholar] [CrossRef]
- Carpentier, B.; Cerf, O. Review—Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 2011, 145, 1–8. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Hebraud, M.; Chorianopoulos, N.; Langsrud, S.; Moretro, T.; Habimana, O.; Desvaux, M.; Renier, S.; Nychas, G.J. Attachment and biofilm formation by foodborne bacteria in meat processing environments: Causes, implications, role of bacterial interactions and control by alternative novel methods. Meat Sci. 2014, 97, 298–309. [Google Scholar] [CrossRef]
- Dutta, V.; Elhanafi, D.; Kathariou, S. Conservation and distribution of the benzalkonium chloride resistance cassette bcrABC in Listeria monocytogenes. Appl. Environ. Microbiol. 2013, 79, 6067–6074. [Google Scholar] [CrossRef]
- Elhanafi, D.; Dutta, V.; Kathariou, S. Genetic characterization of plasmid-associated benzalkonium chloride resistance determinants in a Listeria monocytogenes strain from the 1998–1999 outbreak. Appl. Environ. Microbiol. 2010, 76, 8231–8238. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Møretrø, T.; Heir, E.; Briandet, R.; Langsrud, S. Cleaning and disinfection of biofilms composed of Listeria monocytogenes and background microbiota from meat processing surfaces. J. Appl. Environ. Microbiol. 2017, 83, e01046-17. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Langsrud, S.; Schirmer, B.C.; Møretrø, T.; Heir, E. Genome analysis of Listeria monocytogenes sequence type 8 strains persisting in salmon and poultry processing environments and comparison with related strains. PLoS ONE 2016, 11, e0151117. [Google Scholar] [CrossRef] [PubMed]
- Zupančič, J.; Raghupathi, P.K.; Houf, K.; Burmølle, M.; Sørensen, S.J.; Gunde-Cimerman, N. Synergistic Interactions in Microbial Biofilms Facilitate the Establishment of Opportunistic Pathogenic Fungi in Household Dishwashers. Front. Microbiol. 2018, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Elias, S.; Banin, E. Multi-species biofilms: Living with friendly neighbors. FEMS Microbiol. Rev. 2012, 36, 990–1004. [Google Scholar] [CrossRef]
- Guillier, L.; Stahl, V.; Hezard, B.; Notz, E.; Briandet, R. Modelling the competitive growth between Listeria monocytogenes and biofilm microflora of smear cheese wooden shelves. Int. J. Food Microbiol. 2008, 128, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Mariani, C.; Oulahal, N.; Chamba, J.-F.; Dubois-Brissonnet, F.; Notz, E.; Briandet, R. Inhibition of Listeria monocytogenes by resident biofilms present on wooden shelves used for cheese ripening. Food Control 2011, 22, 1357–1362. [Google Scholar] [CrossRef]
- Arihara, K.; Cassens, R.G.; Luchansky, J.B. Characterization of bacteriocins from Enterococcus faecium with activity against Listeria monocytogenes. Int. J. Food Microbiol. 1993, 19, 123–134. [Google Scholar] [CrossRef]
- Bremer, P.J.; Monk, I.; Osborne, C.M. Survival of Listeria monocytogenes attached to stainless steel surfaces in the presence or absence of Flavobacterium spp. J. Food Prot. 2001, 64, 1369–1376. [Google Scholar] [CrossRef]
- Shedleur-Bourguignon, F.; Thériault, W.P.; Longpré, J.; Thibodeau, A.; Fravalo, P. Use of an ecosystem-based approach to shed light on the heterogeneity of the contamination pattern of Listeria monocytogenes on conveyor belt surfaces in a swine slaughterhouse in the province of Quebec, Canada. Pathogens 2021, 10, 1368. [Google Scholar] [CrossRef]
- Overney, A.; Jacques-André-Coquin, J.; Ng, P.; Carpentier, B.; Guillier, L.; Firmesse, O. Impact of environmental factors on the culturability and viability of Listeria monocytogenes under conditions encountered in food processing plants. Int. J. Food Microbiol. 2017, 244, 74–81. [Google Scholar] [CrossRef]
- Pagotto, F.; Hebert, K.; Farber, J. Isolation of Listeria monocytogenes and Other Listeria spp. from Foods and Environmental Samples; HPB Methods for the Microbiological Analysis of Foods, Method MFHPB-30; Health Canada, Health Products and Food Branch: Ottawa, ON, Canada, 2011; Volume 2. [Google Scholar]
- Kérouanton, A.; Marault, M.; Petit, L.; Grout, J.; Dao, T.T.; Brisabois, A. Evaluation of a multiplex PCR assay as an alternative method for Listeria monocytogenes serotyping. J. Microbiol. Methods. 2010, 80, 134–137. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.M.; Proctor, D.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley-Interscience: New York, NY, USA; London, UK, 1969; p. 286. [Google Scholar]
- Faust, K.; Raes, J. CoNet app: Inference of biological association networks using Cytoscape. F1000Research 2016, 5, 1519. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.B. 400: A Method for combining non-independent, one-sided tests of significance. Biometrics 1975, 31, 987–992. [Google Scholar] [CrossRef]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. In Proceedings of the International AAAI Conference on Web and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Hu, Y. Algorithms for visualizing large networks. Comb. Probab. Comput. 2011, 5, 180–186. [Google Scholar]
- Traag, V.A.; Waltman, L.; Van Eck, N.J. From Louvain to Leiden: Guaranteeing well-connected communities. Sci. Rep. 2019, 9, 5233. [Google Scholar] [CrossRef]
- Autio, T.; Säteri, T.; Fredriksson-Ahomaa, M.; Rahkio, M.; Lundén, J.; Korkeala, H. Listeria monocytogenes contamination pattern in pig slaughterhouses. J. Food Prot. 2000, 63, 1438–1442. [Google Scholar] [CrossRef]
- Nesbakken, T.; Kapperud, G.; Caugant, D.A. Pathways of Listeria monocytogenes contamination in the meat processing industry. Int. J. Food Microbiol. 1996, 31, 161–171. [Google Scholar] [CrossRef]
- Marriott, N.G.; Schilling, M.W.; Gravani, R.B. Food Contamination Sources. In Principles of Food Sanitation; Springer International Publishing: Cham, Switzerland, 2018; pp. 83–91. [Google Scholar]
- Rodríguez-López, P.; Bernárdez, M.; Rodríguez-Herrera, J.J.; Comesaña, Á.S.; Cabo, M.L. Identification and metagenetic characterisation of Listeria monocytogenes-harbouring communities present in food-related industrial environments. Food Control 2019, 95, 6–17. [Google Scholar] [CrossRef]
- Brightwell, G.; Boerema, J.; Mills, J.; Mowat, E.; Pulford, D. Identifying the bacterial community on the surface of Intralox™ belting in a meat boning room by culture-dependent and culture-independent 16S rDNA sequence analysis. Int. J. Food Microbiol. 2006, 109, 47–53. [Google Scholar] [CrossRef]
- Hoo, H.; Hashidoko, Y.; Islam, M.T.; Tahara, S. Requirement of a relatively high threshold level of Mg2+ for cell growth of a rhizoplane bacterium, Sphingomonas yanoikuyae EC-S001. Appl. Environ. Microbiol. 2004, 70, 5214–5221. [Google Scholar] [CrossRef]
- Giaouris, E.; Chorianopoulos, N.; Doulgeraki, A.; Nychas, G.-J. Co-culture with Listeria monocytogenes within a dual-species biofilm community strongly increases resistance of Pseudomonas putida to benzalkonium chloride. PLoS ONE 2013, 8, e77276. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, B.; Chassaing, D. Interactions in biofilms between Listeria monocytogenes and resident microorganisms from food industry premises. Int. J. Food Microbiol. 2004, 97, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, Y.; Wu, H.; Høiby, N.; Molin, S.; Song, Z.J. Current understanding of multi-species biofilms. Int. J. Oral Sci. 2011, 3, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Podtburg, T.C.; Zhao, P.; Chen, D.; Baker, D.A.; Cords, B.; Doyle, M.P. Reduction by Competitive Bacteria of Listeria monocytogenes in Biofilms and Listeria Bacteria in Floor Drains in a Ready-to-eat Poultry Processing Plant. J. Food Prot. 2013, 76, 601–607. [Google Scholar] [CrossRef]
- Fox, E.M.; Solomon, K.; Moore, J.E.; Wall, P.G.; Fanning, S. Phylogenetic profiles of in-house microflora in drains at a food production facility: Comparison and biocontrol implications of Listeria-positive and -negative bacterial populations. J. Appl. Environ. Microbiol. 2014, 80, 3369–3374. [Google Scholar] [CrossRef]
- Kim, S.-J.; Park, S.-J.; Jung, M.-Y.; Kim, J.-G.; Madsen, E.L.; Rhee, S.-K. An uncultivated nitrate-reducing member of the genus Herminiimonas degrades toluene. J. Appl. Environ. Microbiol. 2014, 80, 3233–3243. [Google Scholar] [CrossRef]
- Oyaert, M.; De Baere, T.; Breyne, J.; De Laere, E.; Mariën, S.; Waets, P.; Laffut, W. First Case of Pseudoclavibacter bifida Bacteremia in an Immunocompromised Host with Chronic Obstructive Pulmonary Disease (COPD). J. Clin. Microbiol. 2013, 51, 1973–1976. [Google Scholar] [CrossRef][Green Version]
- Mageswari, A.; Subramanian, P.; Srinivasan, R.; Karthikeyan, S.; Gothandam, K.M. Astaxanthin from psychrotrophic Sphingomonas faeni exhibits antagonism against food-spoilage bacteria at low temperatures. Microbiol. Res. 2015, 179, 38–44. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, Y.S.; Choi, T.-J.; Lee, W.J.; Kim, Y.T. Paracoccus haeundaensis sp. nov., a Gram-negative, halophilic, astaxanthin-producing bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1699–1702. [Google Scholar] [CrossRef]
- Oren, A. Characterization of pigments of prokaryotes and their use in taxonomy and classification. In Methods in Microbiology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 38, pp. 261–282. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).