
High Abundance of Thaumarchaeota Found in Deep Metamorphic Subsurface in Eastern China

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Sample Collection
2.2. Morphological Observation of Microbes in Biofilm
2.3. DNA Extraction from Biofilm, Amplification of 16S rRNA Genes, and Sequencing
2.4. Microbial Community Analyses
2.5. Phylogenetic Analysis of Phylum Thaumarchaeota from the Deep Subsurface
3. Results and Discussion
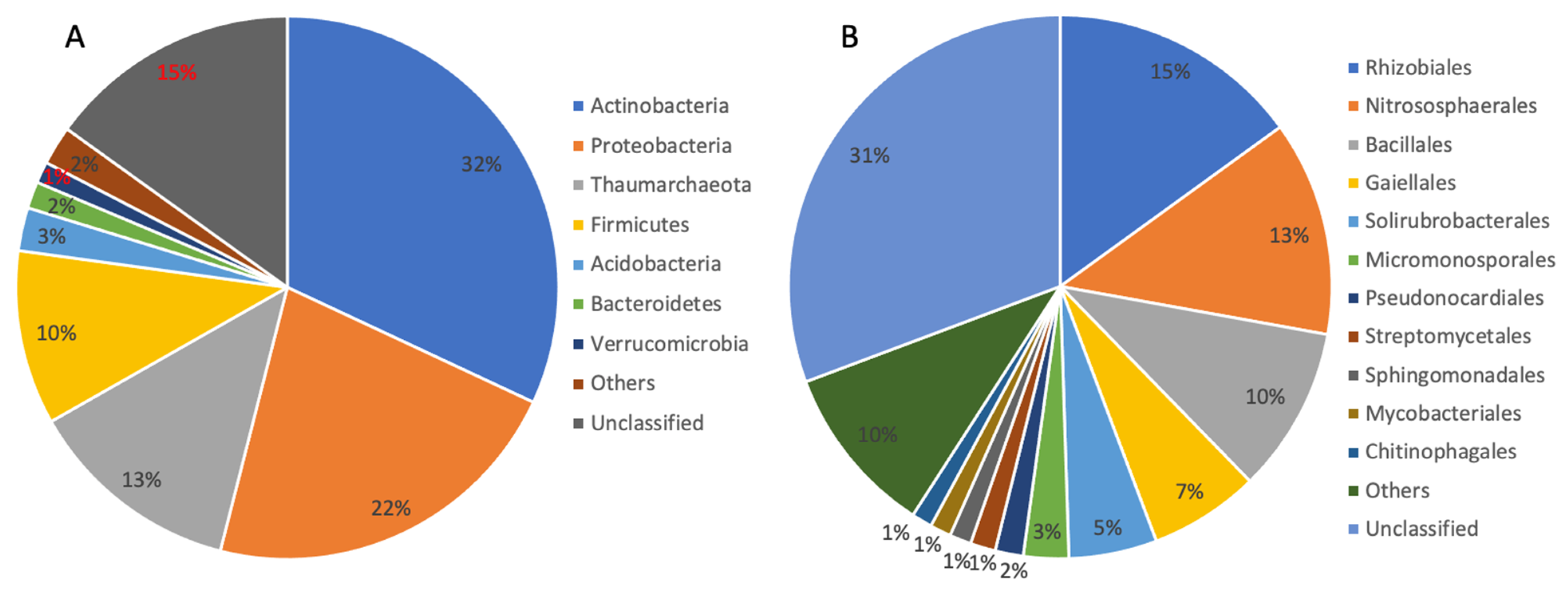
3.1. Microbial Community in the Biofilm Sample
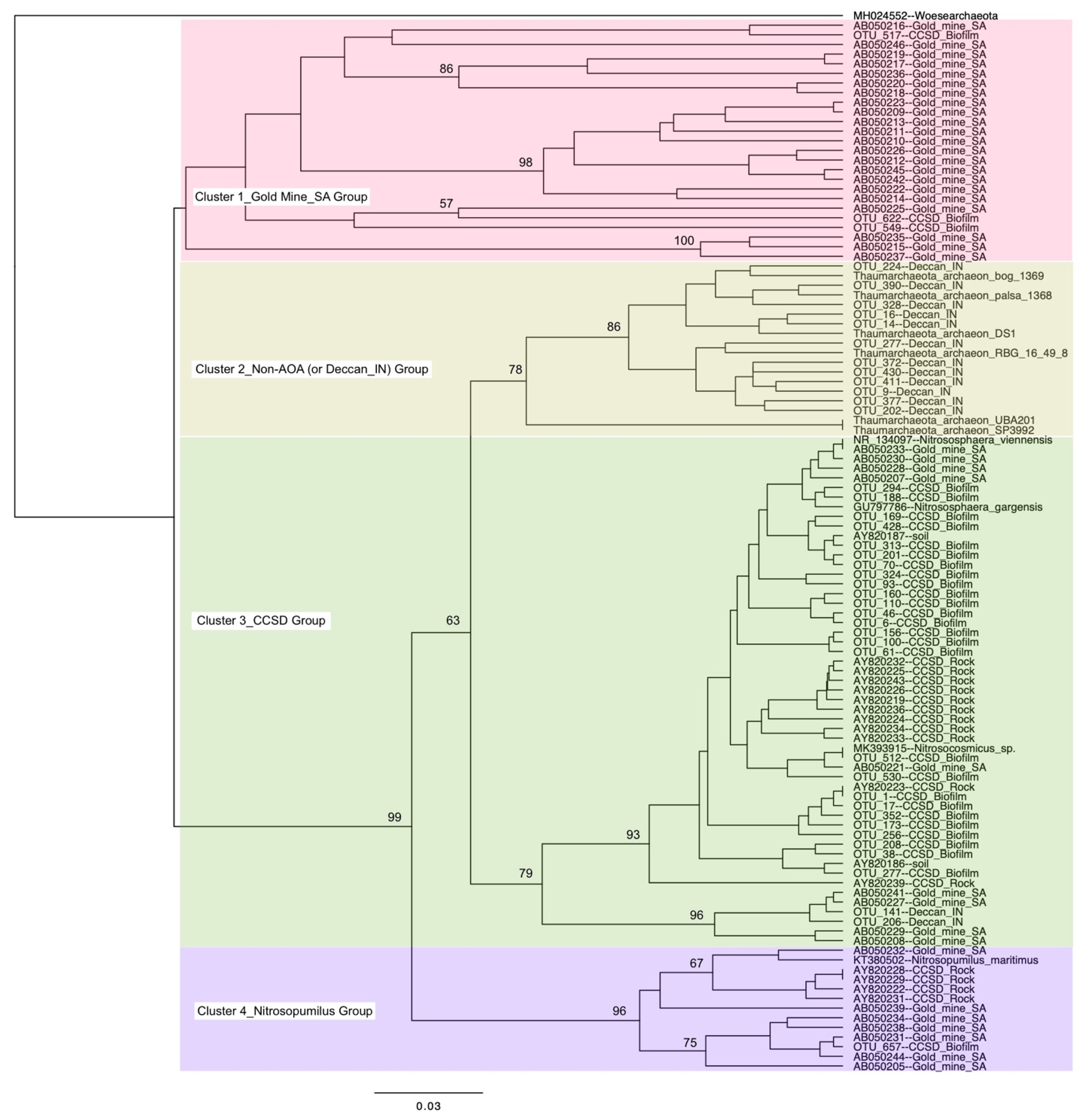
3.2. Thaumarchaeota Diversity in the Deep Subsurface
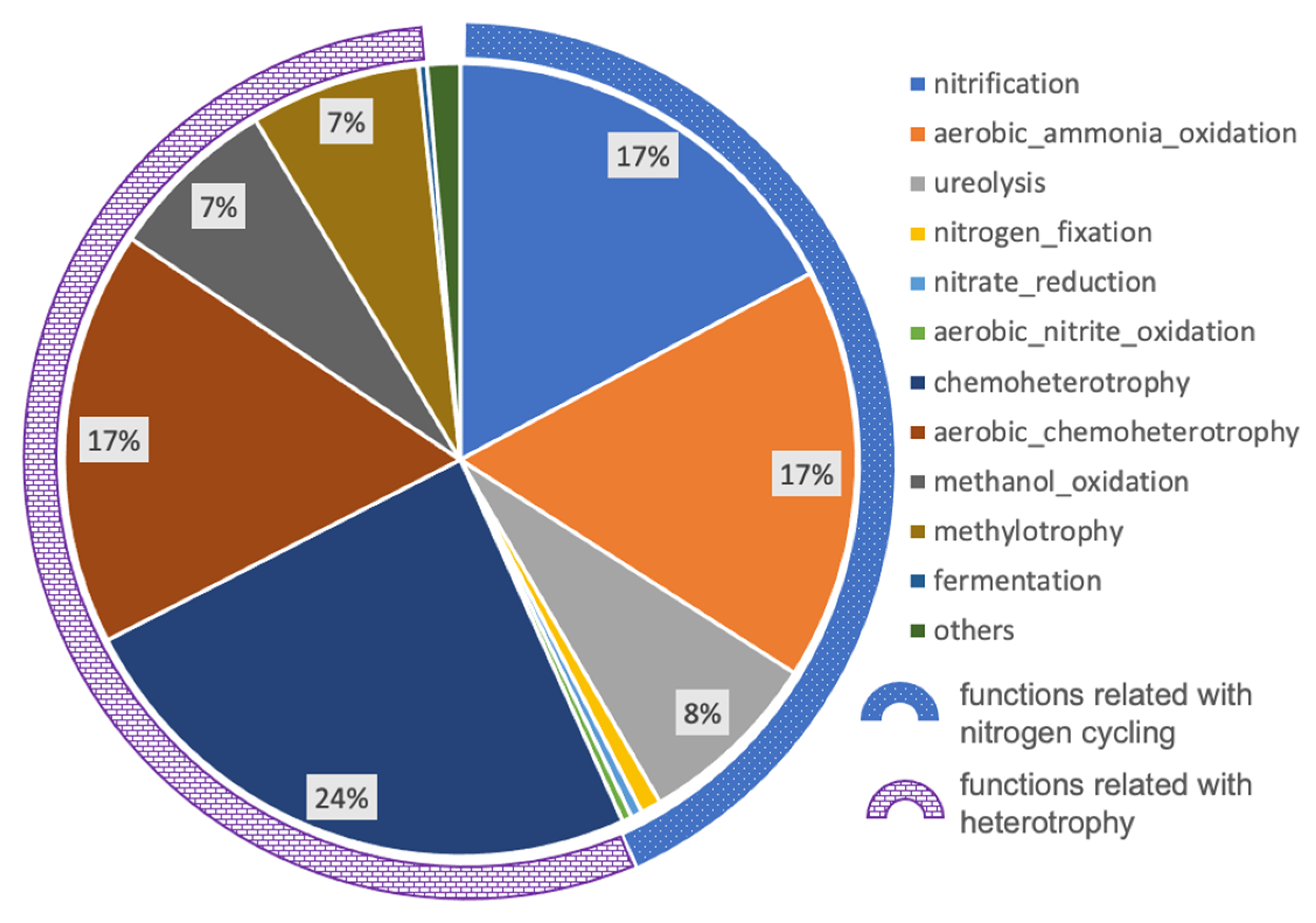
3.3. Potential Nitrogen Cycling in the Deep Subsurface
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brochier-Armanet, C.; Boussau, B.; Gribaldo, S.; Forterre, P. Mesophilic Crenarchaeota: Proposal for a third archaeal phylum, the Thaumarchaeota. Nat. Rev. Microbiol. 2008, 6, 245–252. [Google Scholar] [CrossRef]
- Reji, L.; Cardarelli, E.L.; Boye, K.; Bargar, J.R.; Francis, C.A. Diverse ecophysiological adaptations of subsurface Thaumarchaeota in floodplain sediments revealed through genome-resolved metagenomics. ISME J. 2021. [Google Scholar] [CrossRef]
- Ren, M.; Feng, X.; Huang, Y.; Wang, H.; Hu, Z.; Clingenpeel, S.; Swan, B.K.; Fonseca, M.M.; Posada, D.; Stepanauskas, R.; et al. Phylogenomics suggests oxygen availability as a driving force in Thaumarchaeota evolution. ISME J. 2019, 13, 2150–2161. [Google Scholar] [CrossRef]
- Pester, M.; Schleper, C.; Wagner, M. The Thaumarchaeota: An emerging view of their phylogeny and ecophysiology. Curr. Opin. Microbiol. 2011, 14, 300–306. [Google Scholar] [CrossRef] [Green Version]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [Green Version]
- Kitzinger, K.; Padilla, C.C.; Marchant, H.K.; Hach, P.F.; Herbold, C.W.; Kidane, A.T.; Konneke, M.; Littmann, S.; Mooshammer, M.; Niggemann, J.; et al. Cyanate and urea are substrates for nitrification by Thaumarchaeota in the marine environment. Nat. Microbiol. 2019, 4, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Gubry-Rangin, C.; Kratsch, C.; Williams, T.A.; McHardy, A.C.; Embley, T.M.; Prosser, J.I.; Macqueen, D.J. Coupling of diversification and pH adaptation during the evolution of terrestrial Thaumarchaeota. Proc. Natl. Acad. Sci. USA 2015, 112, 9370–9375. [Google Scholar] [CrossRef] [Green Version]
- Jung, M.Y.; Park, S.J.; Min, D.; Kim, J.S.; Rijpstra, W.I.; Sinninghe Damste, J.S.; Kim, G.J.; Madsen, E.L.; Rhee, S.K. Enrichment and characterization of an autotrophic ammonia-oxidizing archaeon of mesophilic crenarchaeal group I.1a from an agricultural soil. Appl. Environ. Microbiol. 2011, 77, 8635–8647. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Seuradge, B.J.; Neufeld, J.D. Biogeography of soil Thaumarchaeota in relation to soil depth and land usage. FEMS Microbiol. Ecol. 2017, 93, fiw246. [Google Scholar] [CrossRef]
- Cardarelli, E.L.; Bargar, J.R.; Francis, C.A. Diverse Thaumarchaeota Dominate Subsurface Ammonia-oxidizing Communities in Semi-arid Floodplains in the Western United States. Microb. Ecol. 2020, 80, 778–792. [Google Scholar] [CrossRef]
- de la Torre, J.R.; Walker, C.B.; Ingalls, A.E.; Konneke, M.; Stahl, D.A. Cultivation of a thermophilic ammonia oxidizing archaeon synthesizing crenarchaeol. Environ. Microbiol. 2008, 10, 810–818. [Google Scholar] [CrossRef]
- Hedlund, B.P.; Dodsworth, J.A.; Cole, J.K.; Panosyan, H.H. An integrated study reveals diverse methanogens, Thaumarchaeota, and yet-uncultivated archaeal lineages in Armenian hot springs. Antonie Van Leeuwenhoek 2013, 104, 71–82. [Google Scholar] [CrossRef]
- Hou, W.; Wang, S.; Dong, H.; Jiang, H.; Briggs, B.R.; Peacock, J.P.; Huang, Q.; Huang, L.; Wu, G.; Zhi, X.; et al. A comprehensive census of microbial diversity in hot springs of Tengchong, Yunnan Province China using 16S rRNA gene pyrosequencing. PLoS ONE 2013, 8, e53350. [Google Scholar] [CrossRef]
- Jiang, H.; Huang, Q.; Dong, H.; Wang, P.; Wang, F.; Li, W.; Zhang, C. RNA-based investigation of ammonia-oxidizing archaea in hot springs of Yunnan Province, China. Appl. Environ. Microbiol. 2010, 76, 4538–4541. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Dong, H.; Hou, W.; Jiang, H.; Huang, Q.; Briggs, B.R.; Huang, L. Greater temporal changes of sediment microbial community than its waterborne counterpart in Tengchong hot springs, Yunnan Province, China. Sci. Rep. 2014, 4, 7479. [Google Scholar] [CrossRef]
- Wang, S.; Hou, W.G.; Dong, H.L.; Jiang, H.C.; Huang, L.Q.; Wu, G.; Zhang, C.L.; Song, Z.Q.; Zhang, Y.; Ren, H.L.; et al. Control of Temperature on Microbial Community Structure in Hot Springs of the Tibetan Plateau. PLoS ONE 2013, 8, e62901. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.L.; Ye, Q.; Huang, Z.; Li, W.; Chen, J.; Song, Z.; Zhao, W.; Bagwell, C.; Inskeep, W.P.; Ross, C.; et al. Global occurrence of archaeal amoA genes in terrestrial hot springs. Appl. Environ. Microbiol. 2008, 74, 6417–6426. [Google Scholar] [CrossRef] [Green Version]
- Bar-On, Y.M.; Phillips, R.; Milo, R. The biomass distribution on Earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [Green Version]
- Dong, H. Recent developments and future directions of deep biosphere research. Chin. Sci. Bull. 2018, 63, 3885–3901. [Google Scholar] [CrossRef]
- Magnabosco, C.; Lin, L.H.; Dong, H.; Bomberg, M.; Ghiorse, W.; Stan-Lotter, H.; Pedersen, K.; Kieft, T.L.; van Heerden, E.; Onstott, T.C. The biomass and biodiversity of the continental subsurface. Nat. Geosci. 2018, 11, 707–717. [Google Scholar] [CrossRef]
- Colman, D.R.; Poudel, S.; Stamps, B.W.; Boyd, E.S.; Spear, J.R. The deep, hot biosphere: Twenty-five years of retrospection. Proc. Natl. Acad. Sci. USA 2017, 114, 6895–6903. [Google Scholar] [CrossRef] [Green Version]
- Gold, T. The Deep, Hot Biosphere. Proc. Natl. Acad. Sci. USA 1992, 89, 6045–6049. [Google Scholar] [CrossRef] [Green Version]
- Stevens, T.O.; McKinley, J.P. Lithoautotrophic microbial ecosystems in deep basalt aquifers. Science 1995, 270, 450–455. [Google Scholar] [CrossRef]
- D’Hondt, S.; Inagaki, F.; Zarikian, C.A.; Abrams, L.J.; Dubois, N.; Engelhardt, T.; Evans, H.; Ferdelman, T.; Gribsholt, B.; Harris, R.N.; et al. Presence of oxygen and aerobic communities from sea floor to basement in deep-sea sediments. Nat. Geosci. 2015, 8, 299–304. [Google Scholar] [CrossRef] [Green Version]
- Orcutt, B.N.; Wheat, C.G.; Rouxel, O.; Hulme, S.; Edwards, K.J.; Bach, W. Oxygen consumption rates in subseafloor basaltic crust derived from a reaction transport model. Nat. Commun. 2013, 4, 2539. [Google Scholar] [CrossRef] [Green Version]
- Dutta, A.; Sar, P.; Sarkar, J.; Dutta Gupta, S.; Gupta, A.; Bose, H.; Mukherjee, A.; Roy, S. Archaeal Communities in Deep Terrestrial Subsurface Underneath the Deccan Traps, India. Front. Microbiol. 2019, 10, 1362. [Google Scholar] [CrossRef] [Green Version]
- Takai, K.; Moser, D.P.; DeFlaun, M.; Onstott, T.C.; Fredrickson, J.K. Archaeal diversity in waters from deep South African gold mines. Appl. Environ. Microbiol. 2001, 67, 5750–5760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Dong, H.; Xu, Z.; Zhao, D.; Zhang, C. Microbial diversity in ultra-high-pressure rocks and fluids from the Chinese Continental Scientific Drilling Project in China. Appl. Environ. Microbiol. 2005, 71, 3213–3227. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Hu, S.; Huang, S.; Yang, W.; Wang, J.; Yuan, Y.; Yang, S. Heat flow study at the Chinese Continental Scientific Drilling site: Borehole temperature, thermal conductivity, and radiogenic heat production. J. Geophys. Res. 2008, 113, B02404. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Zhang, Z.; Hoefs, J.; van den Kerkhof, A. Ultrahigh-pressure metamorphic rocks from the Chinese Continental Scientific Drilling Project: II Oxygen isotope and fluid inclusion distributions through vertical sections. Contrib. Mineral. Petrol. 2006, 152, 443–458. [Google Scholar] [CrossRef]
- Luo, L.Q.; Zhan, X.C.; Sun, Q. Fluid geoanalysis in the Chinese continental scientific drilling project. Geostand. Geoanal. Res. 2004, 28, 325–331. [Google Scholar] [CrossRef]
- Xia, Q.Y.; Wang, X.; Zeng, Q.; Guo, D.Y.; Zhu, Z.H.; Chen, H.Y.; Dong, H.L. Mechanisms of Enhanced Antibacterial Activity by Reduced Chitosan-Intercalated Nontronite. Environ. Sci. Technol. 2020, 54, 5207–5217. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Zhang, Z.; Cai, W.; Liu, W.; Xu, M.; Yin, H.; Wang, A.; He, Z.; Deng, Y. Biodiversity and species competition regulate the resilience of microbial biofilm community. Mol. Ecol. 2017, 26, 6170–6182. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Zhang, G.; Dong, H.; Jiang, H.; Xu, Z.; Eberl, D.D. Unique Microbial Community in Drilling Fluids from Chinese Continental Scientific Drilling. Geomicrobiol. J. 2007, 23, 499–514. [Google Scholar] [CrossRef]
- Dai, X.; Wang, Y.; Luo, L.; Pfiffner, S.M.; Li, G.; Dong, Z.; Xu, Z.; Dong, H.; Huang, L. Detection of the deep biosphere in metamorphic rocks from the Chinese continental scientific drilling. Geobiology 2021, 19, 278–291. [Google Scholar] [CrossRef]
- Brochier-Armanet, C.; Gribaldo, S.; Forterre, P. Spotlight on the Thaumarchaeota. ISME J. 2012, 6, 227–230. [Google Scholar] [CrossRef]
- Kraft, B.; Jehmlich, N.; Larsen, M.; Bristow, L.A.; Könneke, M.; Thamdrup, B.; Canfield, D.E. Oxygen and nitrogen production by an ammonia-oxidizing archaeon. Science 2022, 375, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, L.; Franca, L.; Rainey, F.A.; Schumann, P.; Nobre, M.F.; da Costa, M.S. Gaiella occulta gen. nov., sp. nov., a novel representative of a deep branching phylogenetic lineage within the class Actinobacteria and proposal of Gaiellaceae fam. nov. and Gaiellales ord. nov. Syst. Appl. Microbiol. 2011, 34, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Green, P.N. Methylobacterium. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–8. [Google Scholar]
- Whitman, W.B. Solirubrobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–5. [Google Scholar]
- Bose, H.; Dutta, A.; Roy, A.; Gupta, A.; Mukhopadhyay, S.; Mohapatra, B.; Sarkar, J.; Roy, S.; Kazy, S.K.; Sar, P. Microbial diversity of drilling fluids from 3000 m deep Koyna pilot borehole provides insights into the deep biosphere of continental earth crust. Sci. Drill. 2020, 27, 1–23. [Google Scholar] [CrossRef]
- Krumholz, L.R.; McKinley, J.P.; Ulrich, F.A.; Suflita, J.M. Confined subsurface microbial communities in Cretaceous rock. Nature 1997, 386, 64–66. [Google Scholar] [CrossRef]
- Pedersen, K. Microbial life in deep granitic rock. Fems Microbiol. Rev. 1997, 20, 399–414. [Google Scholar] [CrossRef]
- Biggs-Weber, E.; Aigle, A.; Prosser, J.I.; Gubry-Rangin, C. Oxygen preference of deeply-rooted mesophilic thaumarchaeota in forest soil. Soil Biol. Biochem. 2020, 148, 107848. [Google Scholar] [CrossRef]
- Zou, D.; Liu, H.; Li, M. Community, Distribution, and Ecological Roles of Estuarine Archaea. Front. Microbiol. 2020, 11, 2060. [Google Scholar] [CrossRef]
- Kerou, M.; Ponce-Toledo, R.I.; Zhao, R.; Abby, S.S.; Hirai, M.; Nomaki, H.; Takaki, Y.; Nunoura, T.; Jorgensen, S.L.; Schleper, C. Genomes of Thaumarchaeota from deep sea sediments reveal specific adaptations of three independently evolved lineages. ISME J. 2021, 15, 2792–2808. [Google Scholar] [CrossRef]
- Jung, Y.J.; Kim, H.J.; Hur, M. Mesorhizobium terrae sp. nov., a novel species isolated from soil in Jangsu, Korea. Antonie Van Leeuwenhoek 2020, 113, 1279–1287. [Google Scholar] [CrossRef]
- Tao, J.; Wang, S.; Liao, T.; Luo, H. Evolutionary origin and ecological implication of a unique nif island in free-living Bradyrhizobium lineages. ISME J. 2021, 15, 3195–3206. [Google Scholar] [CrossRef] [PubMed]
- Jones, F.P.; Clark, I.M.; King, R.; Shaw, L.J.; Woodward, M.J.; Hirsch, P.R. Novel European free-living, non-diazotrophic Bradyrhizobium isolates from contrasting soils that lack nodulation and nitrogen fixation genes—A genome comparison. Sci. Rep. 2016, 6, 25858. [Google Scholar] [CrossRef] [PubMed]
- Jourand, P.; Giraud, E.; Bena, G.; Sy, A.; Willems, A.; Gillis, M.; Dreyfus, B.; de Lajudie, P. Methylobacterium nodulans sp. nov., for a group of aerobic, facultatively methylotrophic, legume root-nodule-forming and nitrogen-fixing bacteria. Int. J. Syst. Evol. Microbiol. 2004, 54, 2269–2273. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Hou, W.; Zeng, X.; Wang, S.; Dong, H. High Abundance of Thaumarchaeota Found in Deep Metamorphic Subsurface in Eastern China. Microorganisms 2022, 10, 542. https://doi.org/10.3390/microorganisms10030542
Zhang W, Hou W, Zeng X, Wang S, Dong H. High Abundance of Thaumarchaeota Found in Deep Metamorphic Subsurface in Eastern China. Microorganisms. 2022; 10(3):542. https://doi.org/10.3390/microorganisms10030542
Chicago/Turabian StyleZhang, Wenhui, Weiguo Hou, Xiangzhi Zeng, Shang Wang, and Hailiang Dong. 2022. "High Abundance of Thaumarchaeota Found in Deep Metamorphic Subsurface in Eastern China" Microorganisms 10, no. 3: 542. https://doi.org/10.3390/microorganisms10030542
APA StyleZhang, W., Hou, W., Zeng, X., Wang, S., & Dong, H. (2022). High Abundance of Thaumarchaeota Found in Deep Metamorphic Subsurface in Eastern China. Microorganisms, 10(3), 542. https://doi.org/10.3390/microorganisms10030542