
Molecular Epidemiology of Multidrug-Resistant Pneumococci among Ghanaian Children under Five Years Post PCV13 Using MLST

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pneumococcal Isolates and Serotyping
2.2. Antibiotic Susceptibility Testing
2.3. DNA Extraction of MDR Isolates
2.4. PCR Amplification of Resistance Genes
2.5. Virulence Genes Determination
2.6. Multilocus Sequence Typing (MLST)
2.7. Statistical Analysis
2.8. Ethical Approval
3. Results
3.1. Brief Description of Isolates
3.2. Antibiotic Resistance Pattern
3.3. Prevalence of Resistance Genes in MDR Isolates
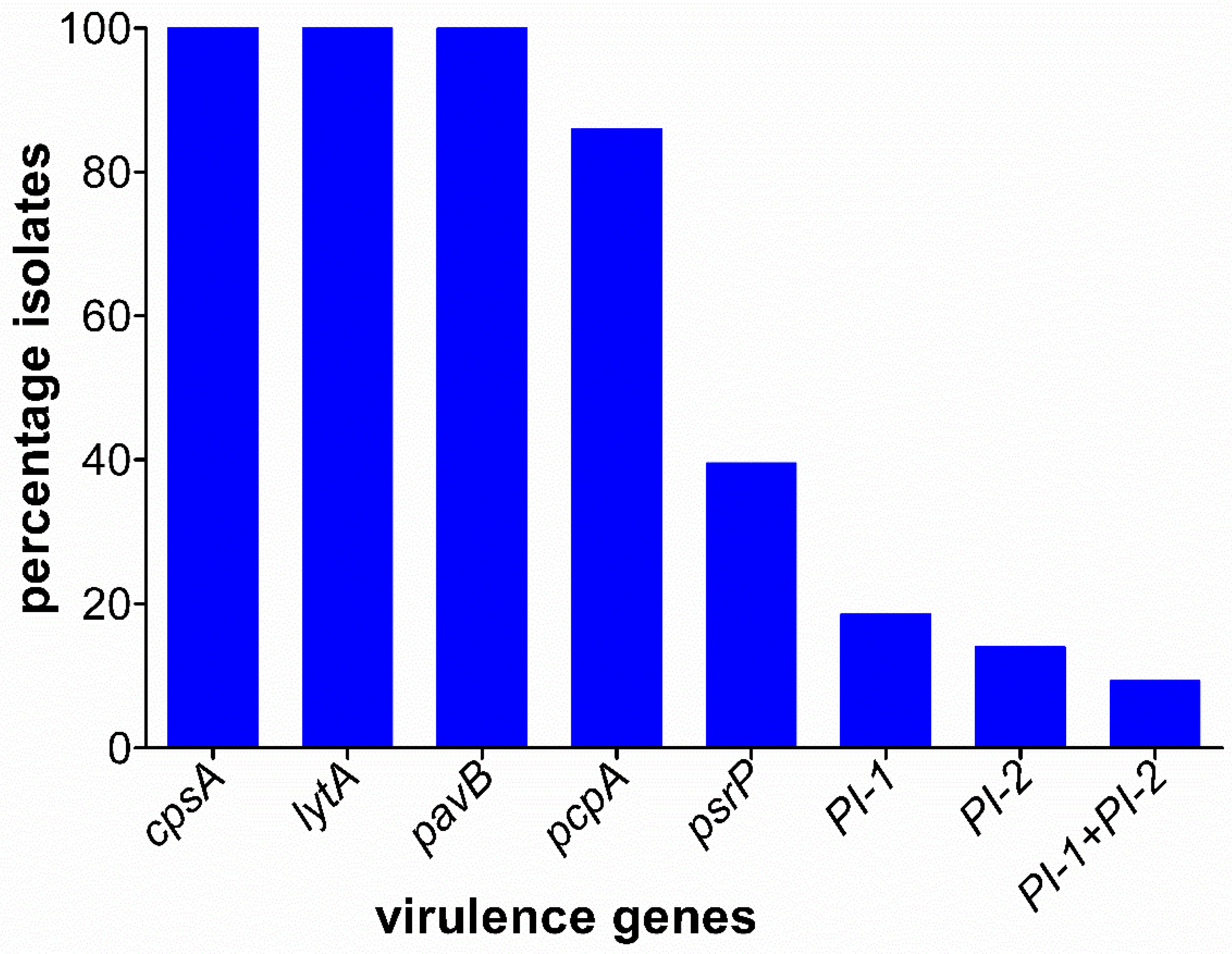
3.4. Prevalence of Virulence Genes
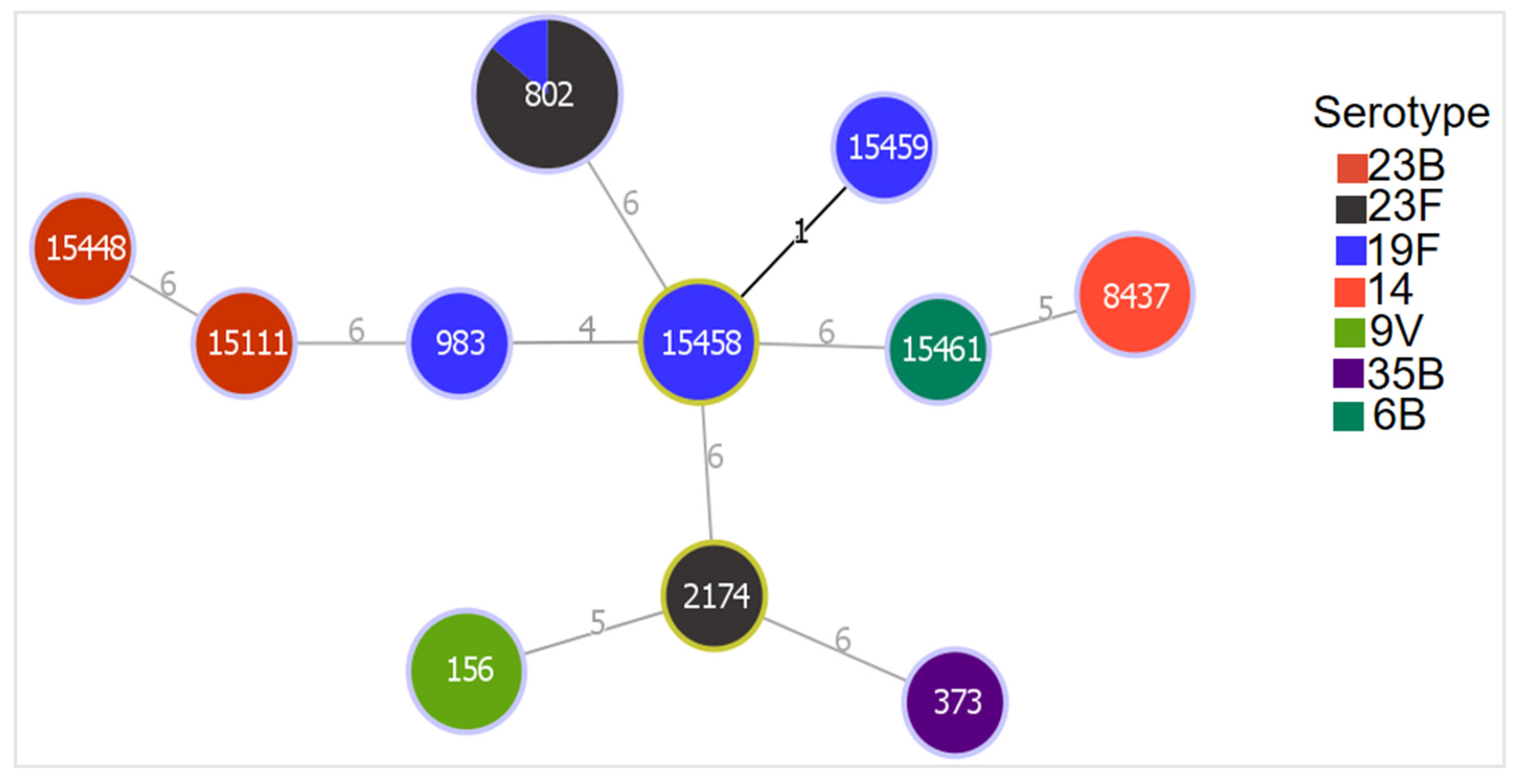
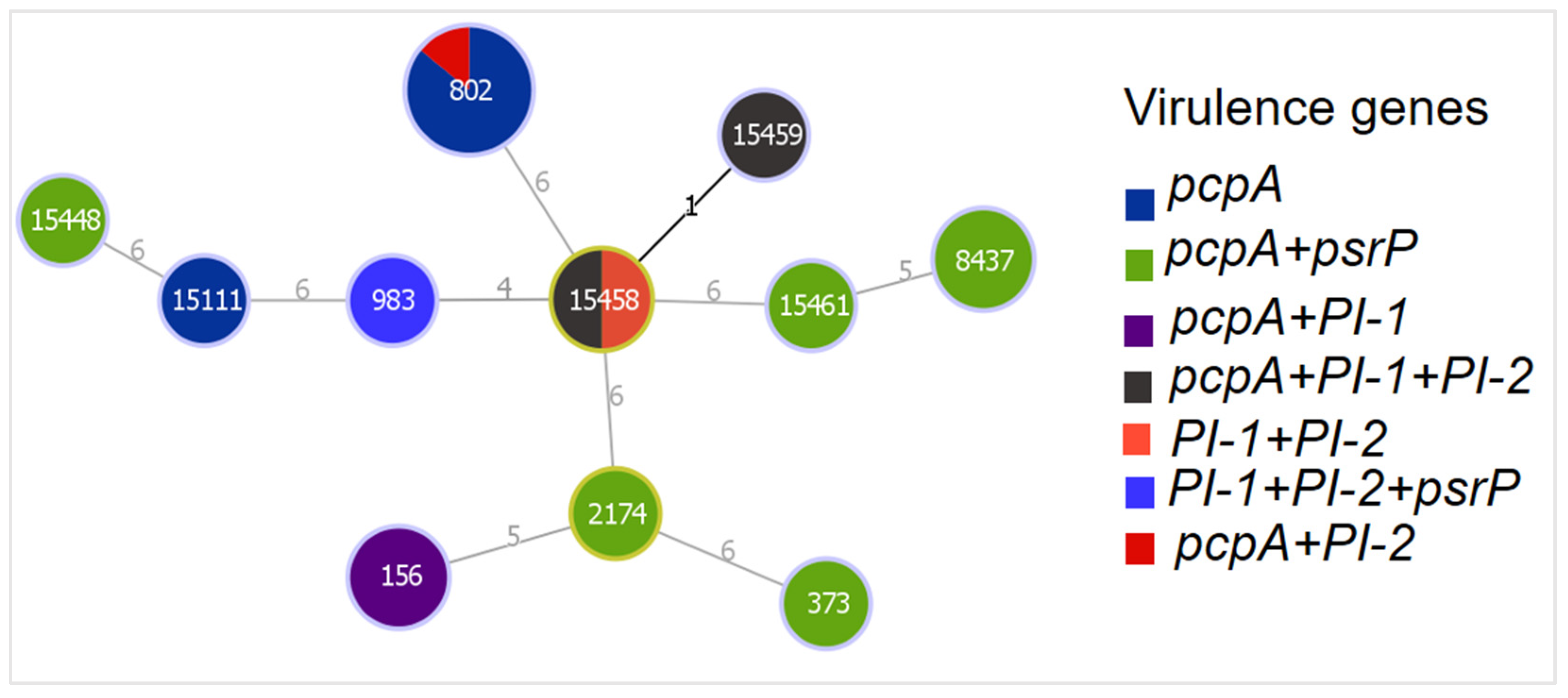
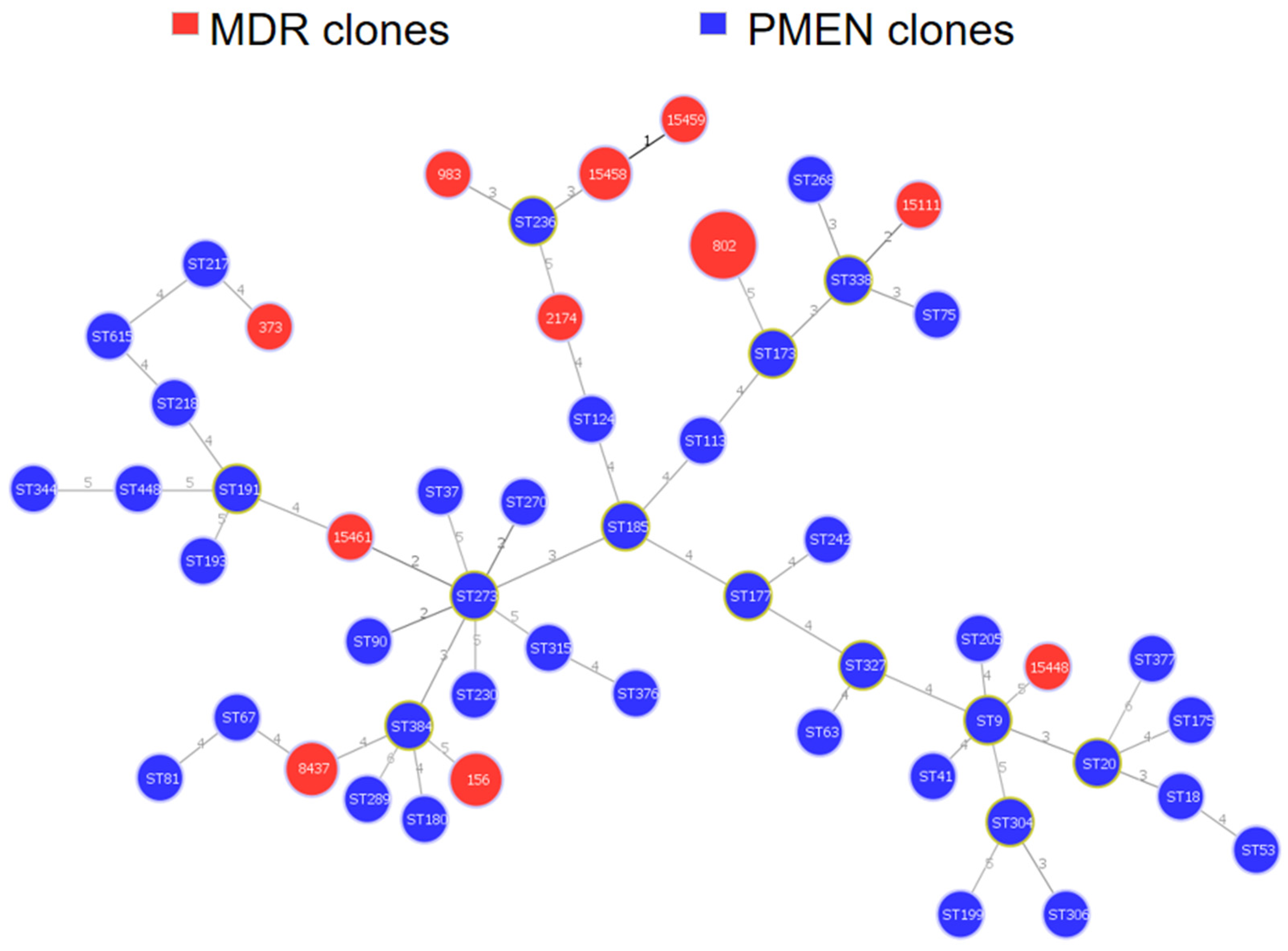
3.5. Genetic Relatedness of MDR Isolates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014; p. xxii. 232p. [Google Scholar]
- Wahl, B.; O’Brien, K.L.; Greenbaum, A.; Majumder, A.; Liu, L.; Chu, Y.; Lukšić, I.; Nair, H.; McAllister, D.A.; Campbell, H.; et al. Burden of Streptococcus pneumoniae and Haemophilus influenzae type b disease in children in the era of conjugate vaccines: Global, regional, and national estimates for 2000–15. Lancet Glob. Health 2018, 6, e744–e757. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Cervero Liceras, F.; Flasche, S.; Sidharta, S.; Yoong, J.; Sundaram, N.; Jit, M. Effect and cost-effectiveness of pneumococcal conjugate vaccination: A global modelling analysis. Lancet Glob. Health 2019, 7, e58–e67. [Google Scholar] [CrossRef] [Green Version]
- Cillóniz, C.; Garcia-Vidal, C.; Ceccato, A.; Torres, A. Antimicrobial Resistance Among Streptococcus pneumoniae. In Antimicrobial Resistance in the 21st Century; Springer: Berlin/Heidelberg, Germany, 2018; pp. 13–38. [Google Scholar] [CrossRef]
- Mills, R.O.; Abdullah, M.R.; Akwetey, S.A.; Sappor, D.C.; Cole, I.; Baffuor-Asare, M.; Bolivar, J.A.; Gamez, G.; van der Linden, M.P.G.; Hammerschmidt, S. Post-Vaccination Streptococcus pneumoniae Carriage and Virulence Gene Distribution among Children Less Than Five Years of Age, Cape Coast, Ghana. Microorganisms 2020, 8, 1987. [Google Scholar] [CrossRef]
- Pichichero, M.E. Pneumococcal whole-cell and protein-based vaccines: Changing the paradigm. Expert Rev. Vaccines 2017, 16, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Andrejko, K.; Ratnasiri, B.; Hausdorff, W.P.; Laxminarayan, R.; Lewnard, J.A. Antimicrobial resistance in paediatric Streptococcus pneumoniae isolates amid global implementation of pneumococcal conjugate vaccines: A systematic review and meta-regression analysis. Lancet Microb. 2021, 2, e450–e460. [Google Scholar] [CrossRef]
- Dayie, N.; Tettey, E.Y.; Newman, M.J.; Bannerman, E.; Donkor, E.S.; Labi, A.K.; Slotved, H.C. Pneumococcal carriage among children under five in Accra, Ghana, five years after the introduction of pneumococcal conjugate vaccine. BMC Pediatr. 2019, 19, 316. [Google Scholar] [CrossRef] [PubMed]
- Dayie, N.T.; Arhin, R.E.; Newman, M.J.; Dalsgaard, A.; Bisgaard, M.; Frimodt-Moller, N.; Slotved, H.C. Multidrug-Resistant Streptococcus pneumoniae Isolates from Healthy Ghanaian Preschool Children. Microb. Drug Resist. 2015, 21, 636–642. [Google Scholar] [CrossRef]
- Mills, R.O.; Twum-Danso, K.; Owusu-Agyei, S.; Donkor, E.S. Epidemiology of pneumococcal carriage in children under five years of age in Accra, Ghana. Infect. Dis. 2015, 47, 326–331. [Google Scholar] [CrossRef]
- Opintan, J.; Newman, M.J.; Arhin, R.E.; Donkor, E.S.; Gyansah-Lutterodt, M.; Mills-Pappoe, W. Laboratory-based nationwide surveillance of antimicrobial resistance in Ghana. Infect. Drug Resist. 2015, 8, 379. [Google Scholar] [CrossRef] [Green Version]
- Raddaoui, A.; Tanfous, F.B.; Chebbi, Y.; Achour, W.; Baaboura, R.; Benhassen, A. High prevalence of multidrug-resistant international clones among macrolide-resistant Streptococcus pneumoniae isolates in immunocompromised patients in Tunisia. Int. J. Antimicrob. Agents 2018, 52, 893–897. [Google Scholar] [CrossRef]
- Schroeder, M.R.; Stephens, D.S. Macrolide Resistance in Streptococcus pneumoniae. Front. Cell Infect. Microbiol. 2016, 6, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; van Tonder, A.; Lambertsen, L.M.; Hakenbeck, R.; Parkhill, J.; Bentley, S.D.; Brueggemann, A.B. Evidence of antimicrobial resistance-conferring genetic elements among pneumococci isolated prior to 1974. BMC Genom. 2013, 14, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donkor, E.S. Molecular typing of the pneumococcus and its application in epidemiology in sub-Saharan Africa. Front. Cell. Infect. Microbiol. 2013, 3, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwambana-Adams, B.A.; Asiedu-Bekoe, F.; Sarkodie, B.; Afreh, O.K.; Kuma, G.K.; Owusu-Okyere, G.; Foster-Nyarko, E.; Ohene, S.-A.; Okot, C.; Worwui, A.K.; et al. An outbreak of pneumococcal meningitis among older children (≥5 years) and adults after the implementation of an infant vaccination programme with the 13-valent pneumococcal conjugate vaccine in Ghana. BMC Infect. Dis. 2016, 16, 575. [Google Scholar] [CrossRef] [Green Version]
- Sparding, N.; Dayie, N.T.; Mills, R.O.; Newman, M.J.; Dalsgaard, A.; Frimodt-Moller, N.; Slotved, H.C. Clonal distribution of pneumococcal serotype 19F isolates from Ghana. Infect. Genet. Evol. 2015, 31, 68–72. [Google Scholar] [CrossRef]
- Renner, L.A.; Usuf, E.; Mohammed, N.I.; Ansong, D.; Dankwah, T.; Kusah, J.T.; Owusu, S.K.; Awunyo, M.; Arhin, B.; Addo, Y.; et al. Hospital-based Surveillance for Pediatric Bacterial Meningitis in the Era of the 13-Valent Pneumococcal Conjugate Vaccine in Ghana. Clin. Infect. Dis. 2019, 69, S89–S96. [Google Scholar] [CrossRef]
- CLSI. M100 Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI: Wayne, PA, USA, 2017; Volume 37. [Google Scholar]
- du Plessis, M.; Smith, A.M.; Klugman, K.P. Rapid detection of penicillin-resistant Streptococcus pneumoniae in cerebrospinal fluid by a seminested-PCR strategy. J. Clin. Microbiol. 1998, 36, 453–457. [Google Scholar] [CrossRef] [Green Version]
- Daly, M.M.; Doktor, S.; Flamm, R.; Shortridge, D. Characterization and Prevalence of MefA, MefE, and the Associated msr(D) Gene in Streptococcus pneumoniae Clinical Isolates. J. Clin. Microbiol. 2004, 42, 3570–3574. [Google Scholar] [CrossRef] [Green Version]
- Shiojima, T.; Fujiki, Y.; Sagai, H.; Iyobe, S.; Morikawa, A. Prevalence of Streptococcus pneumoniae isolates bearing macrolide resistance genes in association with integrase genes of conjugative transposons in Japan. Clin. Microbiol. Infect. 2005, 11, 808–813. [Google Scholar] [CrossRef] [Green Version]
- Malhotra-Kumar, S.; Lammens, C.; Piessens, J.; Goossens, H. Multiplex PCR for simultaneous detection of macrolide and tetracycline resistance determinants in streptococci. Antimicrob. Agents Chemother. 2005, 49, 4798–4800. [Google Scholar] [CrossRef] [Green Version]
- Streptococcus pneumoniae. Available online: https://pubmlst.org/spneumoniae/ (accessed on 22 May 2020).
- Streptococcus Laboratory Resources and Protocols. Available online: https://www.cdc.gov/streplab/pneumococcus/resources.html (accessed on 6 February 2019).
- PHYLOViZ. Available online: https://www.phyloviz.net/ (accessed on 8 June 2020).
- Pneumococcal Molecular Epidemiology Network. Available online: https://www.pneumogen.net/pmen/ (accessed on 20 June 2020).
- Wang, C.-Y.; Chen, Y.-H.; Fang, C.; Zhou, M.-M.; Xu, H.-M.; Jing, C.-M.; Deng, H.-I.; Cai, H.-J.; Jia, K.; Han, S.-Z.; et al. Antibiotic resistance profiles and multidrug resistance patterns of Streptococcus pneumoniae in pediatrics. Medicine 2019, 98, e15942. [Google Scholar] [CrossRef] [PubMed]
- Vorobieva, S.J.V.; Furberg, A.S.; Slotved, H.C.; Bazhukova, T.; Haldorsen, B.; Caugant, D.A.; Sundsfjord, A.; Valentiner-Branth, P.; Simonsen, G.S. Epidemiological and molecular characterization of Streptococcus pneumoniae carriage strains in pre-school children in Arkhangelsk, northern European Russia, prior to the introduction of conjugate pneumococcal vaccines. BMC Infect. Dis. 2020, 20, 279. [Google Scholar] [CrossRef]
- Kobayashi, M.; Conklin, L.M.; Bigogo, G.; Jagero, G.; Hampton, L.; Fleming-Dutra, K.E.; Junghae, M.; Carvalho, M.d.G.; Pimenta, F.; Beall, B.; et al. Pneumococcal carriage and antibiotic susceptibility patterns from two cross-sectional colonization surveys among children aged < 5 years prior to the introduction of 10-valent pneumococcal conjugate vaccine—Kenya, 2009–2010. BMC Infect. Dis. 2017, 17, 25. [Google Scholar] [CrossRef] [Green Version]
- Mbeye, N.M.; ter Kuile, F.O.; Davies, M.A.; Phiri, K.S.; Egger, M.; Wandeler, G.; Ie, D.E.A.S.A. Cotrimoxazole prophylactic treatment prevents malaria in children in sub-Saharan Africa: Systematic review and meta-analysis. Trop. Med. Int. Health 2014, 19, 1057–1067. [Google Scholar] [CrossRef] [Green Version]
- Adetifa, I.M.O.; Antonio, M.; Okoromah, C.A.N.; Ebruke, C.; Inem, V.; Nsekpong, D.; Bojang, A.; Adegbola, R.A. Pre-Vaccination Nasopharyngeal Pneumococcal Carriage in a Nigerian Population: Epidemiology and Population Biology. PLoS ONE 2012, 7, e30548. [Google Scholar] [CrossRef]
- Ministry of Health; Programme, Ghana National Drug Programs (GNDP). Standard Treatment Guidelines, 7th ed.; Yamens Press Limited: Accra, Ghana, 2017. [Google Scholar]
- Adeapena, W.; Afari-Asiedu, S.; Najjemba, R.; Griensven, J.V.; Delamou, A.; Ohene Buabeng, K.; Poku Asante, K. Antibiotic Use in a Municipal Veterinary Clinic in Ghana. Trop. Med. Infect. Dis. 2021, 6, 138. [Google Scholar] [CrossRef]
- Ousmane, S.; Diallo, B.; Ouedraogo, R. Genetic Determinants of Tetracycline Resistance in Clinical Streptococcus pneumoniae Serotype 1 Isolates from Niger. Antibiotics 2018, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Beheshti, M.; Jabalameli, F.; Feizabadi, M.M.; Hahsemi, F.B.; Beigverdi, R.; Emaneini, M. Molecular characterization, antibiotic resistance pattern and capsular types of invasive Streptococcus pneumoniae isolated from clinical samples in Tehran, Iran. BMC Microbiol. 2020, 20, 167. [Google Scholar] [CrossRef]
- Reinert, R.R.; Ringelstein, A.; van der Linden, M.; Cil, M.Y.; Al-Lahham, A.; Schmitz, F.J. Molecular epidemiology of macrolide-resistant Streptococcus pneumoniae isolates in Europe. J. Clin. Microbiol. 2005, 43, 1294–1300. [Google Scholar] [CrossRef] [Green Version]
- Labi, A.-K.; Obeng-Nkrumah, N.; Dayie, N.T.K.D.; Egyir, B.; Sampane-Donkor, E.; Newman, M.J.; Opintan, J.A. Antimicrobial use in hospitalized patients: A multicentre point prevalence survey across seven hospitals in Ghana. JAC-Antimicrob. Resist. 2021, 3, 87. [Google Scholar] [CrossRef]
- Hema-Ouangraoua, S.; Aziz Maiga, A.; Cairns, M.; Zongo, I.; Frederic, N.; Serge Yerbanga, R.; Tamboura, B.; Badji, H.; Gore-Langton, G.; Kuepfer, I.; et al. Impact of the addition of azithromycin to antimalarials used for seasonal malaria chemoprevention on antimicrobial resistance of Streptococcus pneumoniae. Trop. Med. Int. Health 2019, 24, 1442–1454. [Google Scholar] [CrossRef] [PubMed]
- Abdulai, A.A.; Agana-Nsiire, P.; Biney, F.; Kwakye-Maclean, C.; Kyei-Faried, S.; Amponsa-Achiano, K.; Simpson, S.V.; Bonsu, G.; Ohene, S.-A.; Ampofo, W.K.; et al. Community-based mass treatment with azithromycin for the elimination of yaws in Ghana—Results of a pilot study. PLOS Negl. Trop. Dis. 2018, 12, e0006303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzaraly, N.D.; Mohd Desa, M.N.; Muthanna, A.; Masri, S.N.; Taib, N.M.; Suhaili, Z.; Sulaiman, N.; Baharin, N.H.Z.; Shuan, C.Y.; Ariffin, Z.; et al. Antimicrobial susceptibility, serotype distribution, virulence profile and molecular typing of piliated clinical isolates of pneumococci from east coast, Peninsular Malaysia. Sci. Rep. 2021, 11, 8220. [Google Scholar] [CrossRef] [PubMed]
- Safari, D.; Valentiya, F.; Salsabila, K.; Paramaiswari, W.T.; Tafroji, W.; Hammerschmidt, S.; Hadinegoro, S.R. The prevalence of pilus islets in Streptococcus pneumoniae isolates from healthy children in Indonesia. Access. Microbiol. 2021, 3, acmi000184. [Google Scholar] [CrossRef] [PubMed]
- Hjalmarsdottir, M.A.; Petursdottir, B.; Erlendsdottir, H.; Haraldsson, G.; Kristinsson, K.G. Prevalence of pilus genes in pneumococci isolated from healthy preschool children in Iceland: Association with vaccine serotypes and antibiotic resistance. J. Antimicrob. Chemother. 2015, 70, 2203–2208. [Google Scholar] [CrossRef] [Green Version]
- Aguiar, S.I.; Serrano, I.; Pinto, F.R.; Melo-Cristino, J.; Ramirez, M. The presence of the pilus locus is a clonal property among pneumococcal invasive isolates. BMC Microbiol. 2008, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Nurse-Lucas, M.; McGee, L.; Hawkins, P.A.; Swanston, W.H.; Akpaka, P.E. Serotypes and genotypes of Streptococcus pneumoniae isolates from Trinidad and Tobago. Int. J. Infect. Dis. 2016, 46, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Beall, B.; McEllistrem, M.C.; Gertz, R.E.; Wedel, S.; Boxrud, D.J.; Gonzalez, A.L.; Medina, M.J.; Pai, R.; Thompson, T.A.; Harrison, L.H.; et al. Pre- and Postvaccination Clonal Compositions of Invasive Pneumococcal Serotypes for Isolates Collected in the United States in 1999, 2001, and 2002. J. Clin. Microbiol. 2006, 44, 999–1017. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, G.; Garcia-Garcia, S.; Lanaspa, M.; Ciruela, P.; Esteva, C.; Fernandez de Sevilla, M.; Diaz-Conradi, A.; Marti, C.; Motje, M.; Galles, C.; et al. Serotype and clonal distribution dynamics of invasive pneumococcal strains after PCV13 introduction (2011-2016): Surveillance data from 23 sites in Catalonia, Spain. PLoS ONE 2020, 15, e0228612. [Google Scholar] [CrossRef]
- Ndlangisa, K.M.; du Plessis, M.; Wolter, N.; de Gouveia, L.; Klugman, K.P.; von Gottberg, A.; Germs, S.A. Population snapshot of Streptococcus pneumoniae causing invasive disease in South Africa prior to introduction of pneumococcal conjugate vaccines. PLoS ONE 2014, 9, e107666. [Google Scholar] [CrossRef] [Green Version]
- Wyres, K.L.; Lambertsen, L.M.; Croucher, N.J.; McGee, L.; von Gottberg, A.; Linares, J.; Jacobs, M.R.; Kristinsson, K.G.; Beall, B.W.; Klugman, K.P.; et al. Pneumococcal capsular switching: A historical perspective. J. Infect. Dis. 2013, 207, 439–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leimkugel, J.; Adams Forgor, A.; Gagneux, S.; Pflüger, V.; Flierl, C.; Awine, E.; Naegeli, M.; Dangy, J.P.; Smith, T.; Hodgson, A.; et al. An Outbreak of Serotype 1Streptococcus pneumoniaeMeningitis in Northern Ghana with Features That Are Characteristic ofNeisseria meningitidisMeningitis Epidemics. J. Infect. Dis. 2005, 192, 192–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pneumococcal Isolates | Antibiotic Resistance Number (%) | Antibiotic Resistance Genes Number (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Serotypes | Number | CRO | ERY | CLI | TET | CHL | COT | PEN | Pbp2b | tetM | ermB | mefA |
| 23B | 10 | 0 | 1(10) | 1(10) | 10(100) | 0(0) | 10(100) | 10(100) | 4(40) | 10(100) | 1(10) | 0(0) |
| 23F | 9 | 0 | 0. | 0 | 9(100) | 7(77.8) | 9(100) | 9(100) | 8(88.8) | 9(100) | 0(0) | 0(0) |
| 19F | 6 | 1(16.7) | 4(66.7) | 4(66.7) | 6(100) | 1(16.7) | 6(100) | 6(100) | 6(100) | 6(100) | 3(50) | 1(16.7) |
| 14 | 4 | 0 | 0 | 0 | 4(100) | 2(50) | 4(100) | 4(100) | 3(75) | 4(100) | 0(0) | 0(0) |
| 6B | 3 | 0 | 1 | 1(33.3) | 3(100) | 0(0) | 3(100) | 3(100) | 2(66.7) | 3(100) | 1(33.3) | 0(0) |
| 15A | 3 | 1(33.3) | 0 | 0 | 3(100) | 1(33.3) | 3(100) | 1(33.3) | 0(0) | 3(100) | 0(0) | 0(0) |
| 35B | 2 | 0 | 2(100) | 2(100) | 2(100) | 0(0) | 2(100) | 1(50) | 0(0) | 2(100) | 2(100) | 0(0) |
| 3 | 2 | 0 | 0 | 0 | 2(100) | 0(0) | 2(100) | 2(100) | 1(50) | 2(100) | 0(0) | 0(0) |
| 9V | 2 | 0 | 2(100) | 0 | 2(100) | 0(0) | 2(100) | 2(100) | 2(100) | 2(100) | 0(0) | 2(100) |
| 6A | 1 | 0 | 0 | 0 | 1(100) | 0(0) | 1(100) | 1(100) | 0(0) | 1(100) | 0(0) | 0(0) |
| 38 | 1 | 0 | 1(100) | 0 | 1(100) | 0(0) | 1(100) | 1(100) | 0(0) | 1(100) | 0(0) | 1(100) |
| Isolate ID | aroE | gdh | gki | recP | spi | xpt | ddl | ST | Serotype |
|---|---|---|---|---|---|---|---|---|---|
| S125 | 7 | 11 | 10 | 1 | 6 | 8 | 1 | 156 | 9V |
| S276 | 7 | 11 | 10 | 1 | 6 | 8 | 1 | 156 | 9V |
| S106 | 7 | 13 | 4 | 5 | 7 | 88 | 9 | 373 | 35B |
| H130 | 10 | 13 | 53 | 1 | 72 | 38 | 31 | 802 | 19F |
| H148 | 10 | 13 | 53 | 1 | 72 | 38 | 31 | 802 | 23F |
| S33 | 10 | 13 | 53 | 1 | 72 | 38 | 31 | 802 | 23F |
| S34 | 10 | 13 | 53 | 1 | 72 | 38 | 31 | 802 | 23F |
| S305 | 10 | 13 | 53 | 1 | 72 | 38 | 31 | 802 | 23F |
| S341 | 10 | 13 | 53 | 1 | 72 | 38 | 31 | 802 | 23F |
| S578 | 10 | 13 | 53 | 1 | 72 | 38 | 31 | 802 | 23F |
| S243 | 15 | 16 | 19 | 15 | 3 | 104 | 63 | 983 | 19F |
| S41 | 7 | 16 | 8 | 8 | 6 | 142 | 14 | 2174 | 23F |
| S294 | 2 | 89 | 9 | 38 | 6 | 1 | 18 | 8437 | 14 |
| S300 | 2 | 89 | 9 | 38 | 6 | 1 | 18 | 8437 | 14 |
| H9 | 4 | 16 | 19 | 15 | 55 | 20 | 31 | 15458 * | 19F |
| S26 | 4 | 16 | 19 | 15 | 55 | 16 | 31 | 15459 * | 19F |
| S85 | 4 | 16 | 19 | 15 | 55 | 20 | 31 | 15458 * | 19F |
| S579 | 8 | 6 | 1 | 2 | 6 | 1 | 31 | 15461 * | 6B |
| S237 | 1 | 43 | 41 | 18 | 13 | 37 | 8 | 15448 * | 23B |
| S238 | 12 | 13 | 8 | 6 | 3 | 6 | 8 | 15111 * | 23B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mills, R.O.; Abdullah, M.R.; Akwetey, S.A.; Sappor, D.C.; Gámez, G.; Hammerschmidt, S. Molecular Epidemiology of Multidrug-Resistant Pneumococci among Ghanaian Children under Five Years Post PCV13 Using MLST. Microorganisms 2022, 10, 469. https://doi.org/10.3390/microorganisms10020469
Mills RO, Abdullah MR, Akwetey SA, Sappor DC, Gámez G, Hammerschmidt S. Molecular Epidemiology of Multidrug-Resistant Pneumococci among Ghanaian Children under Five Years Post PCV13 Using MLST. Microorganisms. 2022; 10(2):469. https://doi.org/10.3390/microorganisms10020469
Chicago/Turabian StyleMills, Richael O., Mohammed R. Abdullah, Samuel A. Akwetey, Dorcas C. Sappor, Gustavo Gámez, and Sven Hammerschmidt. 2022. "Molecular Epidemiology of Multidrug-Resistant Pneumococci among Ghanaian Children under Five Years Post PCV13 Using MLST" Microorganisms 10, no. 2: 469. https://doi.org/10.3390/microorganisms10020469
APA StyleMills, R. O., Abdullah, M. R., Akwetey, S. A., Sappor, D. C., Gámez, G., & Hammerschmidt, S. (2022). Molecular Epidemiology of Multidrug-Resistant Pneumococci among Ghanaian Children under Five Years Post PCV13 Using MLST. Microorganisms, 10(2), 469. https://doi.org/10.3390/microorganisms10020469