
Impact of IFN-γ Deficiency on the Cardiomyocyte Function in the First Stage of Experimental Chagas Disease

 , , ,
, , , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
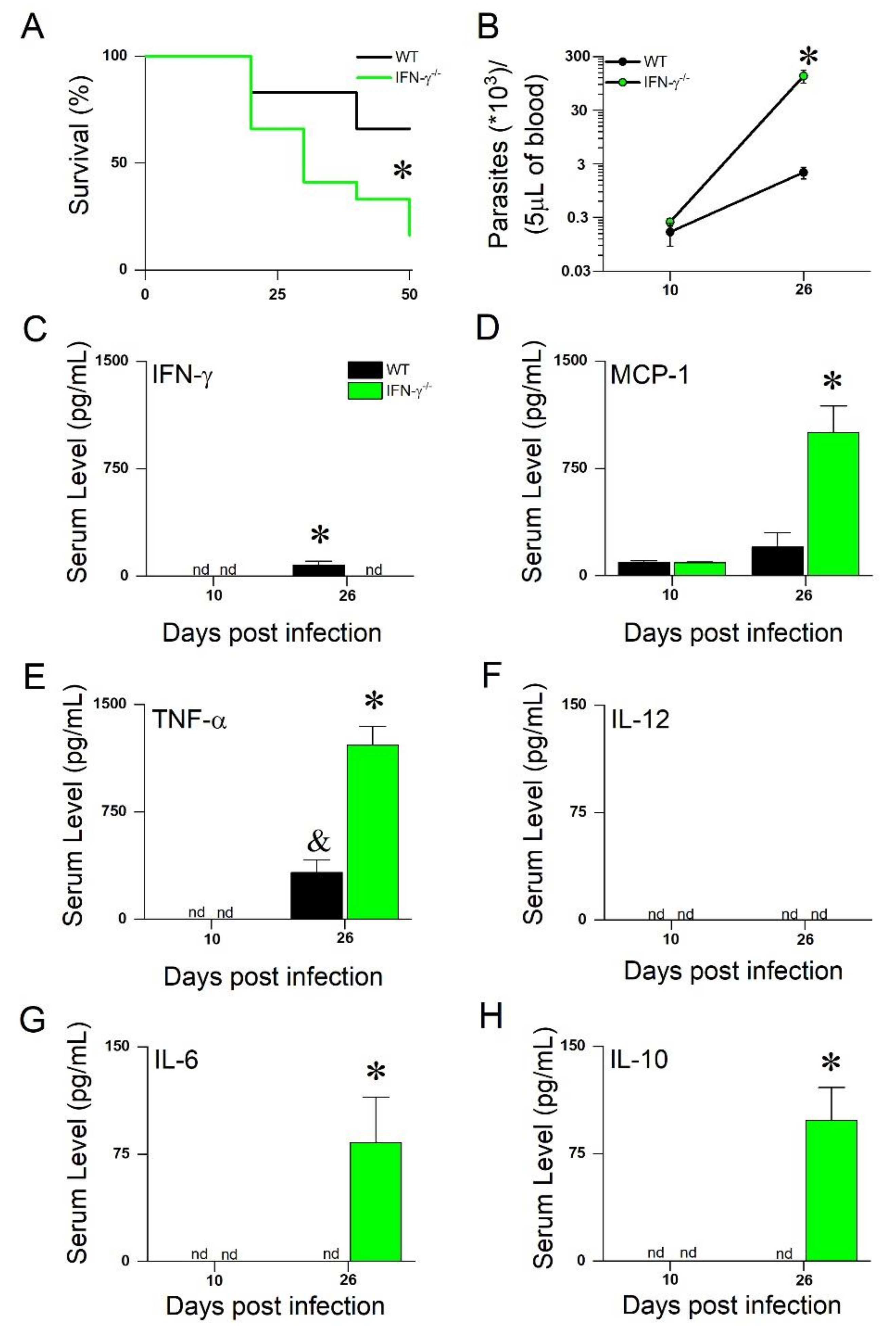
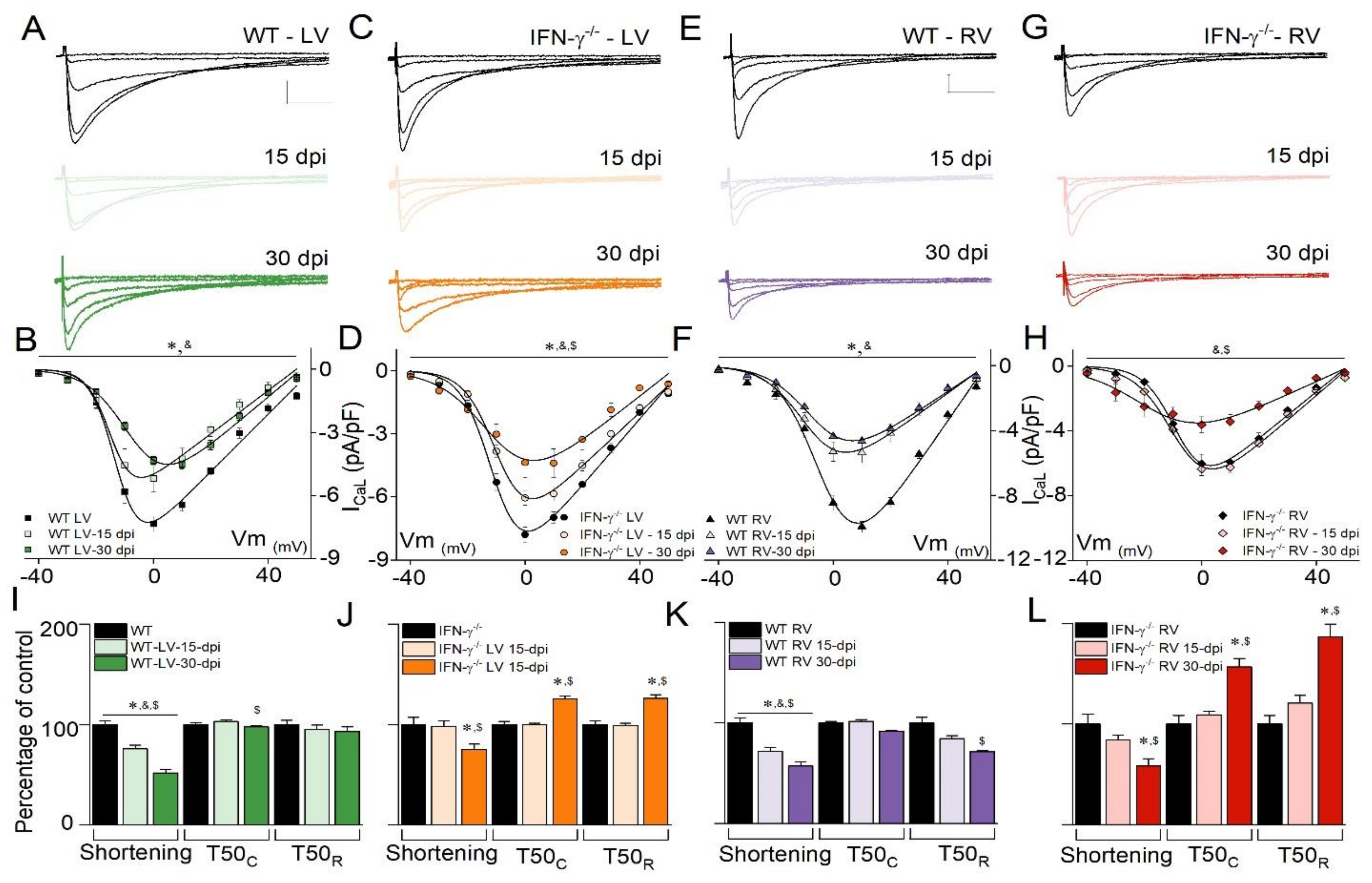
3. Results
4. Discussion
Study Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Fact Sheets-Chagas Disease; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Lidani, K.C.F.; Andrade, F.A.; Bavia, L.; Damasceno, F.S.; Beltrame, M.H.; Messias-Reason, I.J.; Sandri, T.L. Chagas disease: From discovery to a worldwide health problem. Front. Public Health 2019, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Bonney, K.M. Chagas disease in the 21st century: A public health success or an emerging threat? Parasite (Paris, France) 2014, 21, 11. [Google Scholar] [CrossRef] [PubMed]
- Bahia-Oliveira, L.M.; Gomes, J.A.; Rocha, M.O.; Moreira, M.C.; Lemos, E.M.; Luz, Z.M.; Pereira, M.E.S.; Coffman, R.L.; Dias, J.C.P.; Cançado, J.R.; et al. IFN-gamma in human Chagas’ disease: Protection or pathology? Braz. J. Med. Biol. Res. 1998, 31, 127–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha Rodrigues, D.B.; dos Reis, M.A.; Romano, A.; Pereira, S.A.; Teixeira, A.P.; Tostes, S., Jr.; Rodrigues, V. In situ expression of regulatory cytokines by heart inflammatory cell.s in Chagas’ disease patients with heart failure. Clin. Dev. Immunol. 2012, 2012, 361730. [Google Scholar] [PubMed] [Green Version]
- Sousa, G.; Gomes, J.; Fares, R.C.G.; Damásio, M.P.D.S.; Chaves, A.T.; Ferreira, K.S.; Nunes, M.C.P.; Medeiros, N.; Valente, V.A.A.; Correa-Oliveira, R.; et al. Plasma cytokine expression is associated with cardiac morbidity in chagas disease. PLoS ONE 2014, 9, e87082. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.R.; Frade, A.F.; Baron, M.A.; Navarro, I.C.; Kalil, J.; Chevillard, C.; Cunha-Neto, E. Interferon-gamma and other inflammatory mediators in cardiomyocyte signaling during Chagas disease cardiomyopathy. World J. Cardio. 2014, 6, 782–790. [Google Scholar] [CrossRef] [PubMed]
- AALAS. Available online: https://www.aalas.org/about-aalas/position-papers/scientific-basis-for-regulation-of-animal-care-and-use2021 (accessed on 29 November 2021).
- Shioya, T. A simple technique for isolating healthy heart cells from mouse models. J. Physiol. Sci. 2007, 57, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman-Campos, D.; Duarte, H.L.L.; Sales, P.A.; Natali, A.J.; Ropert, C.; Gazzinelli, R.T.; Cruz, J.S. Changes in cellular contractility and cytokines profile during Trypanosoma cruzi infection in mice. Basic Res. Cardiol. 2009, 104, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Gazzinelli, R.T.; Oswald, I.P.; Hieny, S.; James, S.L.; Sher, A. The microbicidal activity of interferon-gamma-treated macrophages against Trypanosoma cruzi involves an L-arginine-dependent, nitrogen oxide-mediated mechanism inhibitable by interleukin-10 and transforming growth factor-beta. Eur. J. Immunol. 1992, 22, 2501–2506. [Google Scholar] [CrossRef] [PubMed]
- Santiago, H.C.; Lombana, C.Z.G.; Macedo, J.P.; Utsch, L.; Tafuri, W.L.; Campagnole-Santos, M.J.; Alves, R.O.; Alves-Filho, J.C.F.; Romanha, A.J.; Cunha, F.Q.; et al. NADPH phagocyte oxidase knockout mice control trypanosoma cruzi proliferation, but develop circulatory collapse and succumb to infection. PLoS Negl. Trop. Dis. 2012, 6, e1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman-Campos, D.; Sales-Junior, P.; Santos-Miranda, A.; Joviano-Santos, J.V.; Ropert, C.; Cruz, J.S. Deletion of inducible nitric oxide synthase delays the onset of cardiomyocyte electrical remodeling in experimental Chagas disease. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2020, 1866, 165949. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.C.; Davoli-Ferreira, M.; Medina, T.S.; Sesti-Costa, R.; Silva, G.K.; Lopes, C.D.; Cardozo, L.E.; Gava, F.N.; Lyroni, K.; Dias, F.C.; et al. Canonical PI3Kgamma signaling in myeloid cells restricts Trypanosoma cruzi infection and dampens chagasic myocarditis. Nat. Commun. 2018, 9, 1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman-Campos, D.; Sales-Junior, P.; Duarte, H.L.; Gomes, E.R.; Lara, A.; Campos, P.; Rocha, N.N.; Resende, R.R.; Ferreira, A.; Guatimosim, S.; et al. Novel insights into the development of chagasic cardiomyopathy: Role of PI3Kinase/NO axis. Int. J. Cardiol. 2013, 167, 3011–3020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roman-Campos, D.; Sales-Junior, P.; Costa, A.D.; Souza, D.S.; Santos-Miranda, A.; Joviano-Santos, J.V.; Ropert, C.; Cruz, J.S. Impact of IFN-γ Deficiency on the Cardiomyocyte Function in the First Stage of Experimental Chagas Disease. Microorganisms 2022, 10, 271. https://doi.org/10.3390/microorganisms10020271
Roman-Campos D, Sales-Junior P, Costa AD, Souza DS, Santos-Miranda A, Joviano-Santos JV, Ropert C, Cruz JS. Impact of IFN-γ Deficiency on the Cardiomyocyte Function in the First Stage of Experimental Chagas Disease. Microorganisms. 2022; 10(2):271. https://doi.org/10.3390/microorganisms10020271
Chicago/Turabian StyleRoman-Campos, Danilo, Policarpo Sales-Junior, Alexandre D. Costa, Diego Santos Souza, Artur Santos-Miranda, Julliane V. Joviano-Santos, Catherine Ropert, and Jader S. Cruz. 2022. "Impact of IFN-γ Deficiency on the Cardiomyocyte Function in the First Stage of Experimental Chagas Disease" Microorganisms 10, no. 2: 271. https://doi.org/10.3390/microorganisms10020271
APA StyleRoman-Campos, D., Sales-Junior, P., Costa, A. D., Souza, D. S., Santos-Miranda, A., Joviano-Santos, J. V., Ropert, C., & Cruz, J. S. (2022). Impact of IFN-γ Deficiency on the Cardiomyocyte Function in the First Stage of Experimental Chagas Disease. Microorganisms, 10(2), 271. https://doi.org/10.3390/microorganisms10020271