
The LCP Family Protein, Psr, Is Required for Cell Wall Integrity and Virulence in Streptococcus agalactiae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Growth Curve
2.3. Antibiotic Assay
2.4. Live/Dead Assay
2.5. Triton X-100 Susceptibility
2.6. Lysozyme Susceptibility
2.7. Capsule Measurement by Buoyant Density
2.8. Measurement of Capsule in Supernatants
2.9. Chain Length Analysis
2.10. Fluorescent Vancomycin Assay
2.11. Zebrafish Infection
2.12. Statistical Analysis
3. Results
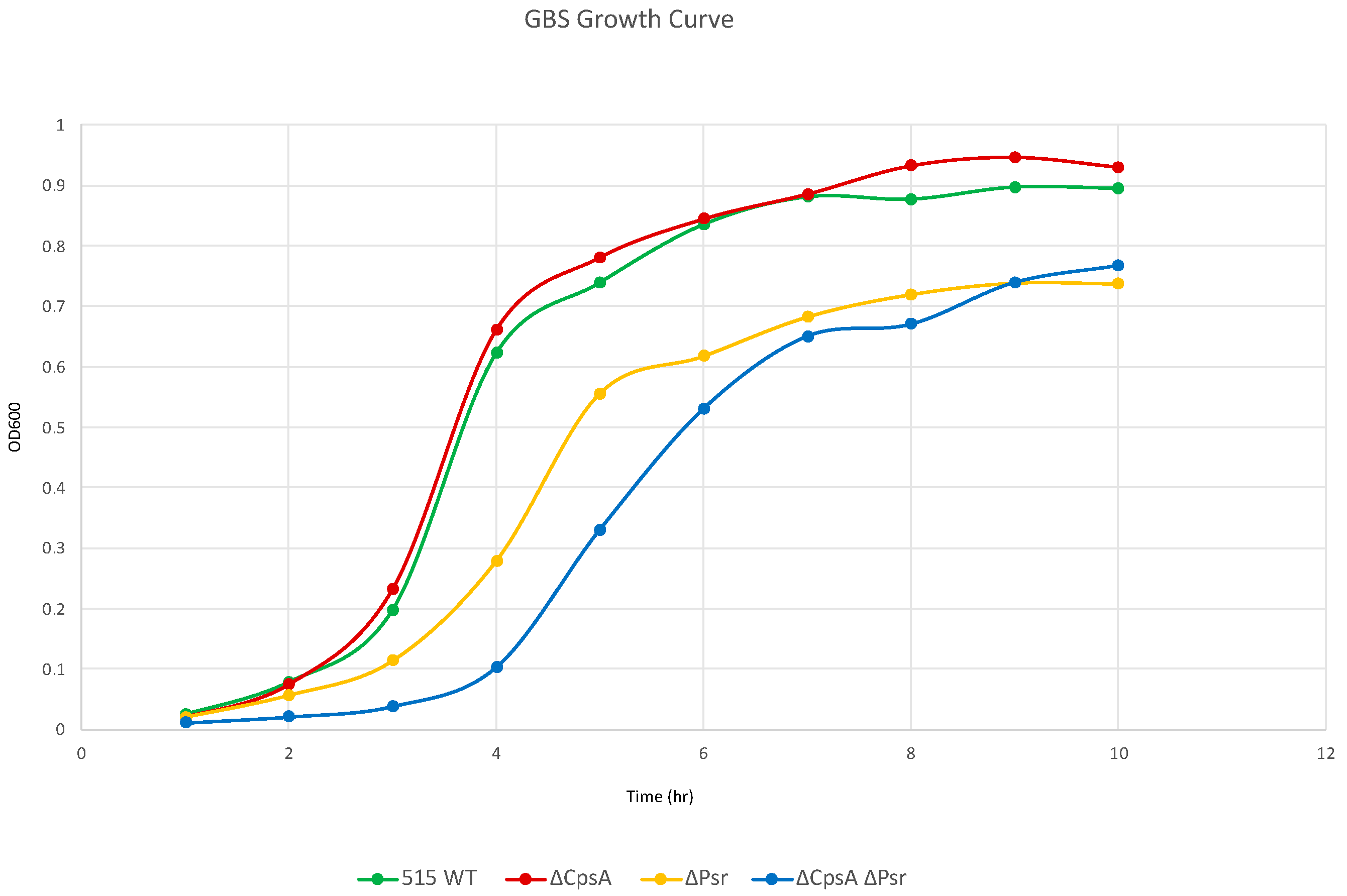
3.1. Loss of Psr Results in a Growth Deficiency
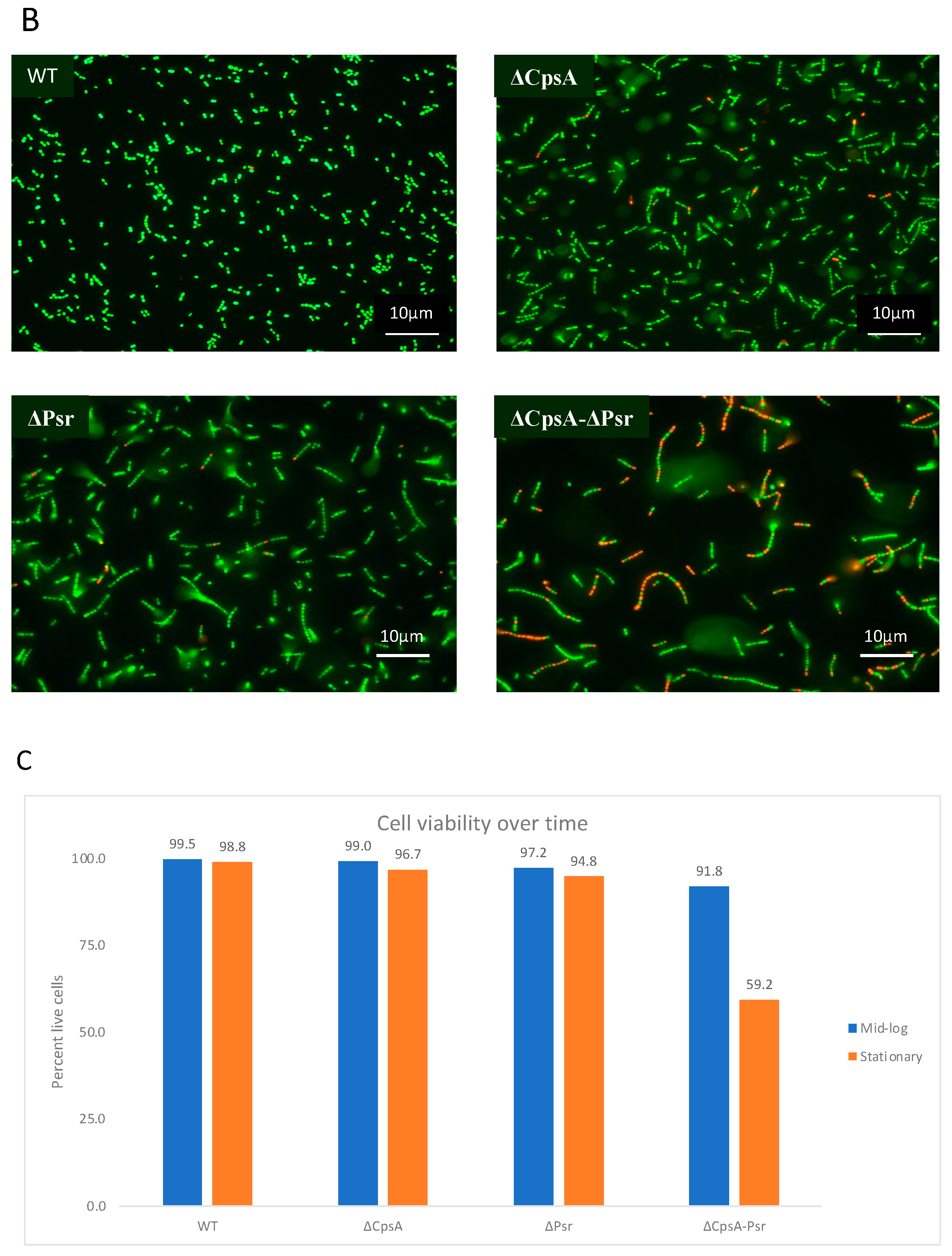
3.2. CpsA-Psr Double Mutant Strain Is Significantly Less Viable over Time
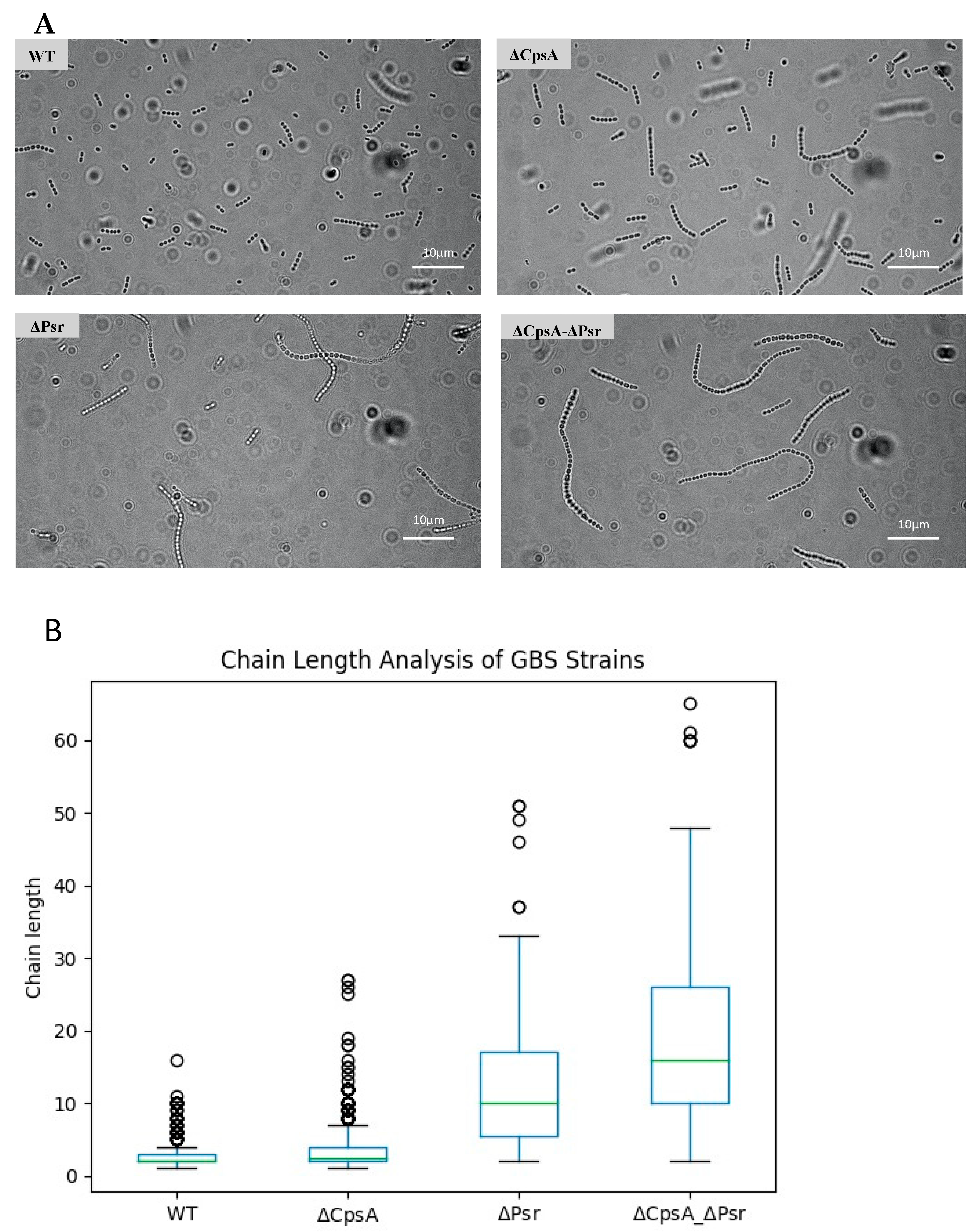
3.3. Loss of CpsA and Psr Results in an Increase in Bacterial Chain Length
3.4. Loss of Psr Results in Changes in Cell Wall Integrity
3.5. Loss of Psr Results in Increased Autolysis
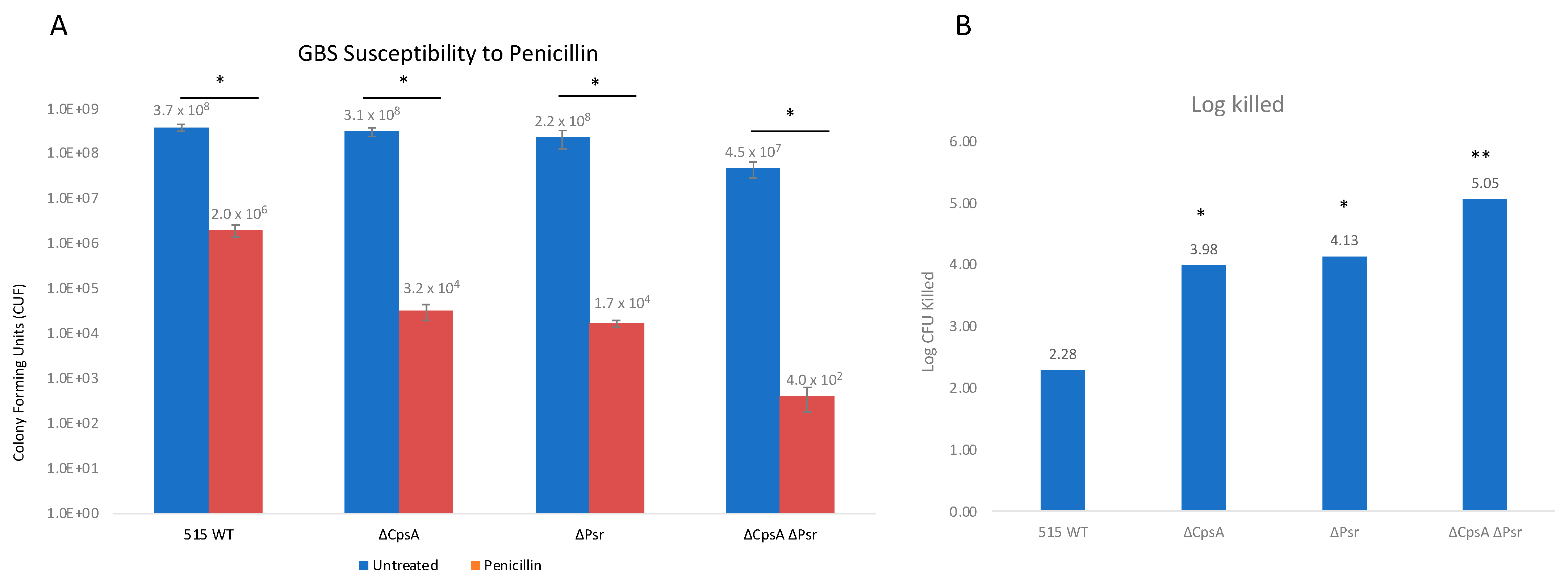
3.6. Absence of CpsA and Psr Result in Increased Susceptibility to Penicillin
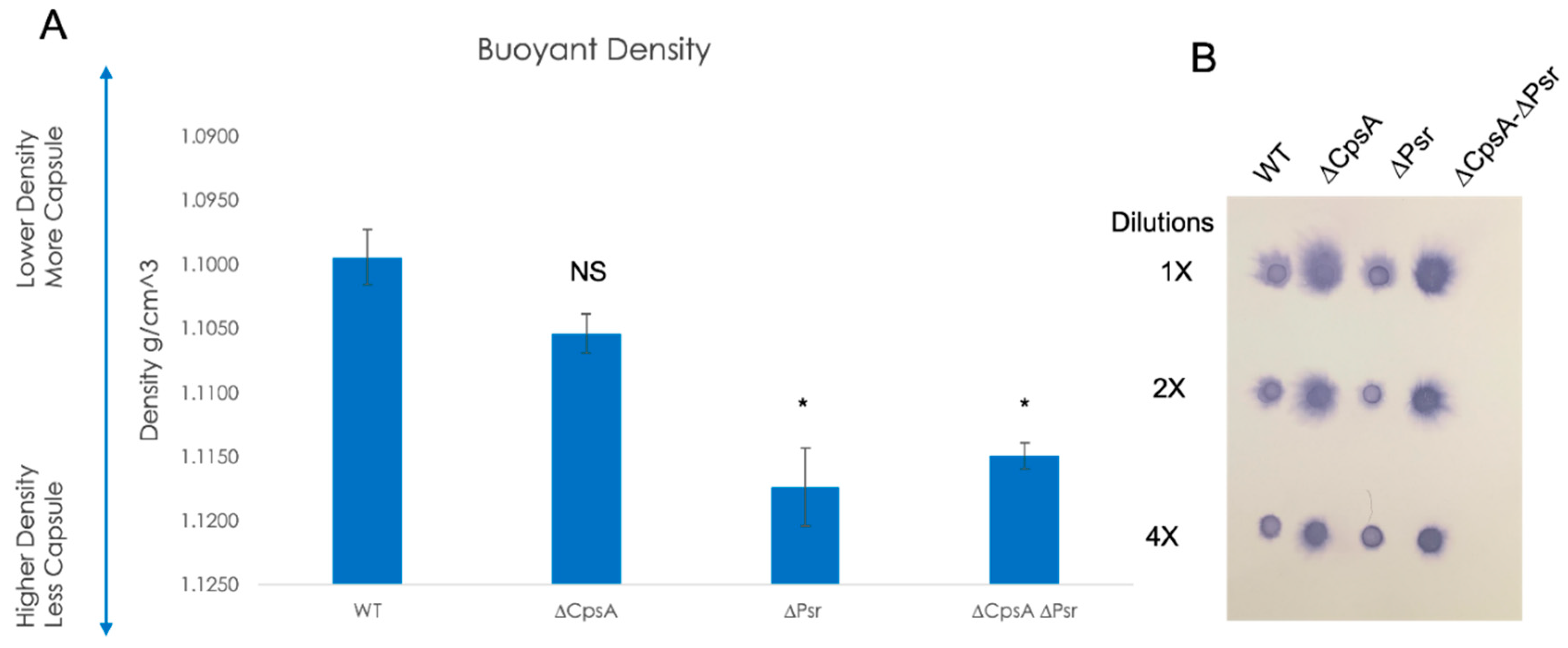
3.7. Absence of CpsA and Psr Results in Decreased Capsule Levels on the Cell
3.8. Absence of CpsA and Psr Results in a Decrease in GBS Virulence In Vivo
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Liakopoulos, A.; Mavroidi, A.; Vourli, S.; Panopoulou, M.; Zachariadou, L.; Chatzipanagiotou, S.; Spiliopoulou, I.; Zerva, L.; Petinaki, E. Molecular characterization of Streptococcus agalactiae from vaginal colonization and neonatal infections: A 4-year multicenter study in Greece. Diagn. Microbiol. Infect. Dis. 2014, 78, 487–490. [Google Scholar] [CrossRef]
- Carl, M.A.; Ndao, I.M.; Springman, A.C.; Manning, S.D.; Johnson, J.R.; Johnston, B.D.; Burnham, C.A.; Weinstock, E.S.; Weinstock, G.M.; Wylie, T.N.; et al. Sepsis from the gut: The enteric habitat of bacteria that cause late-onset neonatal bloodstream infections. Clin. Infect. Dis. 2014, 58, 1211–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwatra, G.; Adrian, P.V.; Shiri, T.; Buchmann, E.J.; Cutland, C.L.; Madhi, S.A. Serotype-specific acquisition and loss of group B streptococcus recto-vaginal colonization in late pregnancy. PLoS ONE 2014, 9, e98778. [Google Scholar] [CrossRef] [Green Version]
- Melin, P. Neonatal group B streptococcal disease: From pathogenesis to preventive strategies. Clin. Microbiol. Infect. 2011, 17, 1294–1303. [Google Scholar] [CrossRef] [Green Version]
- Edmond, K.M.; Kortsalioudaki, C.; Scott, S.; Schrag, S.J.; Zaidi, A.K.; Cousens, S.; Heath, P.T. Group B streptococcal disease in infants aged younger than 3 months: Systematic review and meta-analysis. Lancet 2012, 379, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Patras, K.A.; Nizet, V. Group B Streptococcal Maternal Colonization and Neonatal Disease: Molecular Mechanisms and Preventative Approaches. Front. Pediatr. 2018, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Maisey, H.C.; Doran, K.S.; Nizet, V. Recent advances in understanding the molecular basis of group B Streptococcus virulence. Expert Rev. Mol. Med. 2008, 10, e27. [Google Scholar] [CrossRef] [Green Version]
- Nogacka, A.; Salazar, N.; Suarez, M.; Milani, C.; Arboleya, S.; Solis, G.; Fernandez, N.; Alaez, L.; Hernandez-Barranco, A.M.; de Los Reyes-Gavilan, C.G.; et al. Impact of intrapartum antimicrobial prophylaxis upon the intestinal microbiota and the prevalence of antibiotic resistance genes in vaginally delivered full-term neonates. Microbiome 2017, 5, 93. [Google Scholar] [CrossRef] [Green Version]
- Tapiainen, T.; Koivusaari, P.; Brinkac, L.; Lorenzi, H.A.; Salo, J.; Renko, M.; Pruikkonen, H.; Pokka, T.; Li, W.; Nelson, K.; et al. Impact of intrapartum and postnatal antibiotics on the gut microbiome and emergence of antimicrobial resistance in infants. Sci. Rep. 2019, 9, 10635. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z.H.; Liu, D.; Li, H.D.; Zhu, D.P.; He, Y.; Hou, T.; Yu, J.L. Prenatal and postnatal antibiotic exposure influences the gut microbiota of preterm infants in neonatal intensive care units. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 9. [Google Scholar] [CrossRef] [Green Version]
- Mitre, E.; Susi, A.; Kropp, L.E.; Schwartz, D.J.; Gorman, G.H.; Nylund, C.M. Association Between Use of Acid-Suppressive Medications and Antibiotics during Infancy and Allergic Diseases in Early Childhood. JAMA Pediatr. 2018, 172, e180315. [Google Scholar] [CrossRef] [PubMed]
- Arvonen, M.; Virta, L.J.; Pokka, T.; Kroger, L.; Vahasalo, P. Repeated exposure to antibiotics in infancy: A predisposing factor for juvenile idiopathic arthritis or a sign of this group’s greater susceptibility to infections? J. Rheumatol. 2015, 42, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Heelan, J.S.; Hasenbein, M.E.; McAdam, A.J. Resistance of group B streptococcus to selected antibiotics, including erythromycin and clindamycin. J. Clin. Microbiol. 2004, 42, 1263–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simoes, J.A.; Aroutcheva, A.A.; Heimler, I.; Faro, S. Antibiotic resistance patterns of group B streptococcal clinical isolates. Infect Dis. Obstet. Gynecol. 2004, 12, 259690. [Google Scholar] [CrossRef]
- Capanna, F.; Emonet, S.P.; Cherkaoui, A.; Irion, O.; Schrenzel, J.; Martinez de Tejada, B. Antibiotic resistance patterns among group B Streptococcus isolates: Implications for antibiotic prophylaxis for early-onset neonatal sepsis. Swiss Med. Wkly. 2013, 143, w13778. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Clindamycin-Resistant Group B Streptococcus; CDC: Atlanta, GA, USA, 2019. [Google Scholar]
- Bitoun, J.P.; Liao, S.; McKey, B.A.; Yao, X.; Fan, Y.; Abranches, J.; Beatty, W.L.; Wen, Z.T. Psr is involved in regulation of glucan production, and double deficiency of BrpA and Psr is lethal in Streptococcus mutans. Microbiology 2013, 159, 493–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhardt, A.; Hoyland, C.N.; Vollmer, D.; Bisle, S.; Cleverley, R.M.; Johnsborg, O.; Havarstein, L.S.; Lewis, R.J.; Vollmer, W. Attachment of capsular polysaccharide to the cell wall in Streptococcus pneumoniae. Microb. Drug Resist. 2012, 18, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.G.; Kim, H.K.; Schneewind, O.; Missiakas, D. The capsular polysaccharide of Staphylococcus aureus is attached to peptidoglycan by the LytR-CpsA-Psr (LCP) family of enzymes. J. Biol. Chem. 2014, 289, 15680–15690. [Google Scholar] [CrossRef] [Green Version]
- Hubscher, J.; Luthy, L.; Berger-Bachi, B.; Stutzmann Meier, P. Phylogenetic distribution and membrane topology of the LytR-CpsA-Psr protein family. BMC Genom. 2008, 9, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toniolo, C.; Balducci, E.; Romano, M.R.; Proietti, D.; Ferlenghi, I.; Grandi, G.; Berti, F.; Ros, I.M.; Janulczyk, R. Streptococcus agalactiae capsule polymer length and attachment is determined by the proteins CpsABCD. J. Biol. Chem. 2015, 290, 9521–9532. [Google Scholar] [CrossRef] [Green Version]
- Hanson, B.R.; Runft, D.L.; Streeter, C.; Kumar, A.; Carion, T.W.; Neely, M.N. Functional analysis of the CpsA protein of Streptococcus agalactiae. J. Bacteriol. 2012, 194, 1668–1678. [Google Scholar] [CrossRef] [Green Version]
- Kawai, Y.; Marles-Wright, J.; Cleverley, R.M.; Emmins, R.; Ishikawa, S.; Kuwano, M.; Heinz, N.; Bui, N.K.; Hoyland, C.N.; Ogasawara, N.; et al. A widespread family of bacterial cell wall assembly proteins. EMBO J. 2011, 30, 4931–4941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patras, K.A.; Derieux, J.; Al-Bassam, M.M.; Adiletta, N.; Vrbanac, A.; Lapek, J.D.; Zengler, K.; Gonzalez, D.J.; Nizet, V. Group B Streptococcus Biofilm Regulatory Protein A Contributes to Bacterial Physiology and Innate Immune Resistance. J. Infect. Dis. 2018, 218, 1641–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieslewicz, M.J.; Kasper, D.L.; Wang, Y.; Wessels, M.R. Functional analysis in type Ia group B Streptococcus of a cluster of genes involved in extracellular polysaccharide production by diverse species of streptococci. J. Biol. Chem. 2001, 276, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.D.; Neely, M.N. Large-scale screen highlights the importance of capsule for virulence in the zoonotic pathogen Streptococcus iniae. Infect. Immun. 2005, 73, 921–934. [Google Scholar] [CrossRef] [Green Version]
- Lowe, B.A.; Miller, J.D.; Neely, M.N. Analysis of the polysaccharide capsule of the systemic pathogen Streptococcus iniae and its implications in virulence. Infect. Immun. 2007, 75, 1255–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Zhai, D.; Wu, Z.; Zhao, Y.; Qiao, D.; Zhao, X. Impairment of the Cell Wall Ligase, LytR-CpsA-Psr Protein (LcpC), in Methicillin Resistant Staphylococcus aureus Reduces Its Resistance to Antibiotics and Infection in a Mouse Model of Sepsis. Front. Microbiol. 2020, 11, 557. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.G.; Frankel, M.B.; Dengler, V.; Schneewind, O.; Missiakas, D. Staphylococcus aureus mutants lacking the LytR-CpsA-Psr family of enzymes release cell wall teichoic acids into the extracellular medium. J. Bacteriol. 2013, 195, 4650–4659. [Google Scholar] [CrossRef] [Green Version]
- Rowe, H.M.; Hanson, B.R.; Runft, D.L.; Lin, Q.; Firestine, S.M.; Neely, M.N. Modification of the CpsA protein reveals a role in alteration of the Streptococcus agalactiae cell envelope. Infect. Immun. 2015, 83, 1497–1506. [Google Scholar] [CrossRef] [Green Version]
- Over, B.; Heusser, R.; McCallum, N.; Schulthess, B.; Kupferschmied, P.; Gaiani, J.M.; Sifri, C.D.; Berger-Bachi, B.; Stutzmann Meier, P. LytR-CpsA-Psr proteins in Staphylococcus aureus display partial functional redundancy and the deletion of all three severely impairs septum placement and cell separation. FEMS Microbiol. Lett. 2011, 320, 142–151. [Google Scholar] [CrossRef]
- Bitoun, J.P.; Liao, S.; Yao, X.; Ahn, S.J.; Isoda, R.; Nguyen, A.H.; Brady, L.J.; Burne, R.A.; Abranches, J.; Wen, Z.T. BrpA is involved in regulation of cell envelope stress responses in Streptococcus mutans. Appl. Environ. Microbiol. 2012, 78, 2914–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitoun, J.P.; Liao, S.; Xie, G.G.; Beatty, W.L.; Wen, Z.T. Deficiency of BrpB causes major defects in cell division, stress responses and biofilm formation by Streptococcus mutans. Microbiology 2014, 160, 67–78. [Google Scholar] [CrossRef] [Green Version]
- De, A.; Liao, S.; Bitoun, J.P.; Roth, R.; Beatty, W.L.; Wu, H.; Wen, Z.T. Deficiency of RgpG Causes Major Defects in Cell Division and Biofilm Formation, and Deficiency of LytR-CpsA-Psr Family Proteins Leads to Accumulation of Cell Wall Antigens in Culture Medium by Streptococcus mutans. Appl. Environ. Microbiol. 2017, 83, e00928-17. [Google Scholar] [CrossRef] [Green Version]
- Dengler, V.; Meier, P.S.; Heusser, R.; Kupferschmied, P.; Fazekas, J.; Friebe, S.; Staufer, S.B.; Majcherczyk, P.A.; Moreillon, P.; Berger-Bachi, B.; et al. Deletion of hypothetical wall teichoic acid ligases in Staphylococcus aureus activates the cell wall stress response. FEMS Microbiol. Lett. 2012, 333, 109–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, N.; Waldow, F.; Kohler, T.P.; Rohde, M.; Kreikemeyer, B.; Gomez-Mejia, A.; Hain, T.; Schwudke, D.; Vollmer, W.; Hammerschmidt, S.; et al. Lipoteichoic acid deficiency permits normal growth but impairs virulence of Streptococcus pneumoniae. Nat. Commun. 2017, 8, 2093. [Google Scholar] [CrossRef]
- Primo, E.D.; Otero, L.H.; Ruiz, F.; Klinke, S.; Giordano, W. The disruptive effect of lysozyme on the bacterial cell wall explored by an in-silico structural outlook. Biochem. Mol. Biol. Educ. 2018, 46, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajaei, A.; Rowe, H.M.; Neely, M.N. The LCP Family Protein, Psr, Is Required for Cell Wall Integrity and Virulence in Streptococcus agalactiae. Microorganisms 2022, 10, 217. https://doi.org/10.3390/microorganisms10020217
Rajaei A, Rowe HM, Neely MN. The LCP Family Protein, Psr, Is Required for Cell Wall Integrity and Virulence in Streptococcus agalactiae. Microorganisms. 2022; 10(2):217. https://doi.org/10.3390/microorganisms10020217
Chicago/Turabian StyleRajaei, Atefeh, Hannah M. Rowe, and Melody N. Neely. 2022. "The LCP Family Protein, Psr, Is Required for Cell Wall Integrity and Virulence in Streptococcus agalactiae" Microorganisms 10, no. 2: 217. https://doi.org/10.3390/microorganisms10020217
APA StyleRajaei, A., Rowe, H. M., & Neely, M. N. (2022). The LCP Family Protein, Psr, Is Required for Cell Wall Integrity and Virulence in Streptococcus agalactiae. Microorganisms, 10(2), 217. https://doi.org/10.3390/microorganisms10020217