
Proteomic Shifts Reflecting Oxidative Stress and Reduced Capacity for Protein Synthesis, and Alterations to Mitochondrial Membranes in Neurospora crassa Lacking VDAC

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. N. crassa Strains and Growth Conditions
2.3. Proteomic Analysis of Enriched Mitochondrial and S100 Cytosolic Fractions
2.4. Stress Analysis
2.5. Fatty Acid Analysis
2.6. Sterol Analysis
2.7. Fluidity Analysis
3. Results and Discussion
3.1. Less Abundant Proteins in ΔPor-1
3.2. More Abundant Proteins in ΔPor-1
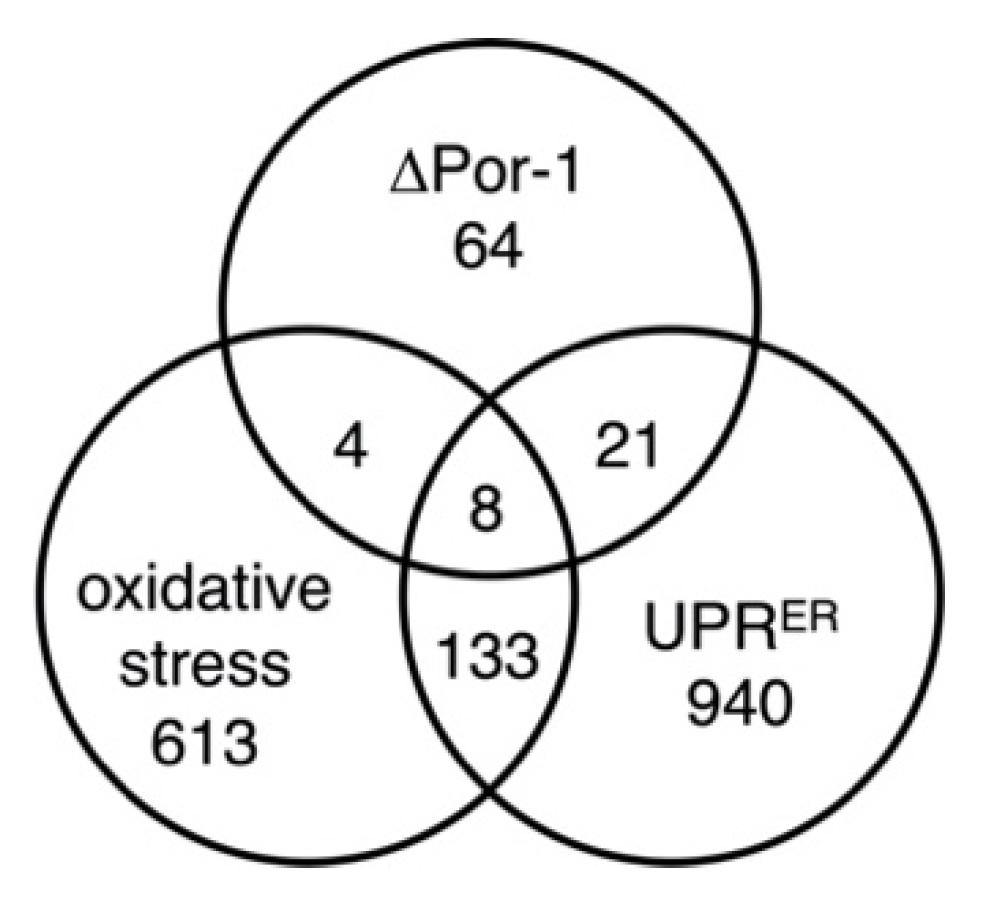
3.3. Unfolded Protein Responses
3.4. Hyphal Morphology
3.5. Mitochondrial Membranes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bay, D.C.; Hafez, M.; Young, M.J.; Court, D.A. Phylogenetic and coevolutionary analysis of the β-barrel protein family comprised of mitochondrial porin (VDAC) and Tom. Biochim. Biophys. Acta (BBA) Biomembr. 2012, 1818, 1502–1519. [Google Scholar] [CrossRef]
- Bayrhuber, M.; Meins, T.; Habeck, M.; Becker, S.; Giller, K.; Villinger, S.; Vonrhein, C.; Griesinger, C.; Zweckstetter, M.; Zeth, K. Structure of the human voltage-dependent anion channel. Proc. Natl. Acad. Sci. USA 2008, 105, 15370–15375. [Google Scholar] [CrossRef]
- Hiller, S.; Garces, R.G.; Malia, T.J.; Orekhov, V.Y.; Colombini, M.; Wagner, G. Solution Structure of the Integral Human Membrane Protein VDAC-1 in Detergent Micelles. Science 2008, 321, 1206–1210. [Google Scholar] [CrossRef]
- Ujwal, R.; Cascio, D.; Colletier, J.-P.; Faham, S.; Zhang, J.; Toro, L.; Ping, P.; Abramson, J. The crystal structure of mouse VDAC1 at 2.3 Å resolution reveals mechanistic insights into metabolite gating. Proc. Natl. Acad. Sci. USA 2008, 105, 17742–17747. [Google Scholar] [CrossRef]
- Rostovtseva, T.; Colombini, M. VDAC channels mediate and gate the flow of ATP: Implications for the regulation of mitochondrial function. Biophys. J. 1997, 72, 1954–1962. [Google Scholar] [CrossRef]
- Benz, R.; Schmid, A.; Dihanich, M. Pores from mitochondrial outer membranes of yeast and a porin-deficient yeast mutant: A comparison. J. Bioenerg. Biomembr. 1989, 21, 439–450. [Google Scholar] [CrossRef]
- Mannella, C.A. Electron microscopy and image analysis of the mitochondrial outer membrane channel, VDAC. J. Bioenerg. Biomembr. 1987, 19, 329–340. [Google Scholar] [CrossRef]
- Shuvo, S.R.; Ferens, F.G.; Court, D.A. The N-terminus of VDAC: Structure, mutational analysis, and a potential role in regulating barrel shape. Biochim. Biophys. Acta (BBA) Biomembr. 2016, 1858, 1350–1361. [Google Scholar] [CrossRef]
- Najbauer, E.E.; Becker, S.; Giller, K.; Zweckstetter, M.; Lange, A.; Steinem, C.; de Groot, B.L.; Griesinger, C.; Andreas, L.B. Structure, gating and interactions of the voltage-dependent anion channel. Eur. Biophys. J. 2021, 50, 159–172. [Google Scholar] [CrossRef]
- Preto, J.; Krimm, I. The intrinsically disordered N-terminus of the voltage-dependent anion channel. PLoS Comput. Biol. 2021, 17, e1008750. [Google Scholar] [CrossRef]
- Nargang, F.E.; Rapaport, D. Neurospora crassa as a Model Organism for Mitochondrial Biogenesis. Methods Mol. Biol. 2007, 372, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Summers, W.A.; Wilkins, J.A.; Dwivedi, R.C.; Ezzati, P.; Court, D.A. Mitochondrial dysfunction resulting from the absence of mitochondrial porin in Neurospora crassa. Mitochondrion 2012, 12, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Shuvo, S.R.; Wiens, L.M.; Subramaniam, S.; Treberg, J.R.; Court, D.A. Increased reactive oxygen species production and maintenance of membrane potential in VDAC-less Neurospora crassa mitochondria. J. Bioenerg. Biomembr. 2019, 51, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Ferens, F.G.; Spicer, V.; Krokhin, O.V.; Motnenko, A.; Summers, W.A.; Court, D.A. A deletion variant partially complements a porin-less strain of Neurospora crassa. Biochem. Cell Biol. 2017, 95, 318–327. [Google Scholar] [CrossRef]
- Colot, H.V.; Park, G.; Turner, G.E.; Ringelberg, C.; Crew, C.M.; Litvinkova, L.; Weiss, R.L.; Borkovich, K.A.; Dunlap, J.C. A high-throughput gene knockout procedure for Neurospora reveals functions for multiple transcription factors. Proc. Natl. Acad. Sci. USA 2006, 103, 10352–10357. [Google Scholar] [CrossRef]
- McCluskey, K.; Wiest, A.; Plamann, M. The Fungal Genetics Stock Center: A repository for 50 years of fungal genetics research. J. Biosci. 2010, 35, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.H.; de Serres, F.J. Genetic and microbiological research techniques for Neurospora crassa. Methods Methane Metab. Part B Methanotrophy 1970, 17, 79–143. [Google Scholar] [CrossRef]
- Harkness, T.; Nargang, F.E.; Van der Klei, I.J.; Neupert, W.; Lill, R. A crucial role of the mitochondrial protein import receptor MOM19 for the biogenesis of mitochondria. J. Cell Biol. 1994, 124, 637–648. [Google Scholar] [CrossRef]
- Ferens, F.G.; Summers, W.A.T.; Bharaj, A.; Stetefeld, J.; Court, D.A. A C-Terminally Truncated Variant of Neurospora crassa VDAC Assembles into a Partially Functional Form in the Mitochondrial Outer Membrane and Forms Multimers in vitro. Front. Physiol. 2021, 12, 739001. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Dwivedi, R.C.; Spicer, V.; Harder, M.; Antonovici, M.; Ens, W.; Standing, K.G.; Wilkins, J.A.; Krokhin, O.V. Practical Implementation of 2D HPLC Scheme with Accurate Peptide Retention Prediction in Both Dimensions for High-Throughput Bottom-Up Proteomics. Anal. Chem. 2008, 80, 7036–7042. [Google Scholar] [CrossRef]
- Michelsen, U.; von Hagen, J. Chapter 19 Isolation of Subcellular Organelles and Structures. Methods Enzymol. 2009, 463, 305–328. [Google Scholar] [CrossRef] [PubMed]
- Craig, R.; Beavis, R.C. TANDEM: Matching proteins with tandem mass spectra. Bioinformatics 2004, 20, 1466–1467. [Google Scholar] [CrossRef]
- McQueen, P.; Spicer, V.; Rydzak, T.; Sparling, R.; Levin, D.; Wilkins, J.A.; Krokhin, O. Information-dependent LC-MS/MS acquisition with exclusion lists potentially generated on-the-fly: Case study using a whole cell digest of Clostridium thermocellum. Proteomics 2012, 12, 1160–1169. [Google Scholar] [CrossRef] [PubMed]
- Gungormusler-Yilmaz, M.; Shamshurin, D.; Grigoryan, M.; Taillefer, M.; Spicer, V.; Krokhin, O.V.; Sparling, R.; Levin, D.B. Reduced catabolic protein expression in Clostridium butyricum DSM 10702 correlate with reduced 1,3-propanediol synthesis at high glycerol loading. AMB Express 2014, 4, 63. [Google Scholar] [CrossRef] [PubMed]
- McQueen, P.; Spicer, V.; Schellenberg, J.; Krokhin, O.; Sparling, R.; Levin, D.; Wilkins, J.A. Whole cell, label free protein quantitation with data independent acquisition: Quantitation at the MS2 level. Proteomics 2015, 15, 16–24. [Google Scholar] [CrossRef]
- Wushke, S.; Spicer, V.; Zhang, X.L.; Fristensky, B.; Krokhin, O.V.; Levin, D.B.; Cicek, N.; Sparling, R. Understanding aerobic/anaerobic metabolism in Caldibacillus debilis through a comparison with model organisms. Syst. Appl. Microbiol. 2017, 40, 245–253. [Google Scholar] [CrossRef]
- Montenegro-Montero, A.; Goity, A.; Larrondo, L.F. The bZIP Transcription Factor HAC-1 Is Involved in the Unfolded Protein Response and Is Necessary for Growth on Cellulose in Neurospora crassa. PLoS ONE 2015, 10, e0131415. [Google Scholar] [CrossRef]
- Fan, F.; Ma, G.; Li, J.; Liu, Q.; Benz, J.P.; Tian, C.; Ma, Y. Genome-wide analysis of the endoplasmic reticulum stress response during lignocellulase production in Neurospora crassa. Biotechnol. Biofuels 2015, 8, 1–17. [Google Scholar] [CrossRef]
- Karaffa, L.; Váczy, K.Z.; Sándor, E.; Biró, S.; Szentirmai, A.; Pócsi, I. Cyanide-resistant alternative respiration is strictly correlated to intracellular peroxide levels in Acremonium chrysogenum. Free. Radic. Res. 2001, 34, 405–416. [Google Scholar] [CrossRef]
- Fernandes, P.N.; Mannarino, S.C.; Silva, C.G.; Pereira, M.D.; Panek, A.D.; Eleutherio, E.C. Oxidative stress response in eukaryotes: Effect of glutathione, superoxide dismutase and catalase on adaptation to peroxide and menadione stresses in Saccharomyces cerevisiae. Redox Rep. 2007, 12, 236–244. [Google Scholar] [CrossRef]
- Loewen, P.C.; Villanueva, J.; Switala, J.; Donald, L.J.; Ivancich, A. Unprecedented access of phenolic substrates to the heme active site of a catalase: Substrate binding and peroxidase-like reactivity of Bacillus pumilus catalase monitored by X-ray crystallography and EPR spectroscopy. Proteins Struct. Funct. Bioinform. 2015, 83, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Sestric, R.; Munch, G.; Cicek, N.; Sparling, R.; Levin, D.B. Growth and neutral lipid synthesis by Yarrowia lipolytica on various carbon substrates under nutrient-sufficient and nutrient-limited conditions. Bioresour. Technol. 2014, 164, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Larsson, L.; Saraf, A. Use of gas chromatography-ion trap tandem mass spectrometry for the detection and characterization of microorganisms in complex samples. Mol. Biotechnol. 1997, 7, 279–287. [Google Scholar] [CrossRef]
- Moghadasian, M.H.; Moghadasian, P.; Le, K.; Hydamaka, A.; Zahradka, P. Lipid analyses of four types of fish from Manitoba lakes. E Cronicon Nutr. 2015, 1, 41–48. [Google Scholar]
- Bari, M.; Battista, N.; Fezza, F.; Finazzi-Agrò, A.; Maccarrone, M. Lipid Rafts Control Signaling of Type-1 Cannabinoid Receptors in Neuronal Cells. J. Biol. Chem. 2005, 280, 12212–12220. [Google Scholar] [CrossRef] [PubMed]
- Ruepp, A.; Zollner, A.; Maier, D.; Albermann, K.; Hani, J.; Mokrejs, M.; Tetko, I.; Güldener, U.; Mannhaupt, G.; Münsterkötter, M.; et al. The FunCat, a functional annotation scheme for systematic classification of proteins from whole genomes. Nucleic Acids Res. 2004, 32, 5539–5545. [Google Scholar] [CrossRef]
- Castilho, B.A.; Shanmugam, R.; Silva, R.; Ramesh, R.; Himme, B.M.; Sattlegger, E. Keeping the eIF2 alpha kinase Gcn2 in check. Biochim. Biophys. Acta 2014, 1843, 1948–1968. [Google Scholar] [CrossRef]
- Carsiotis, M.; Jones, R.F. Cross-Pathway Regulation: Tryptophan-Mediated Control of Histidine and Arginine Biosynthetic Enzymes in Neurospora crassa. J. Bacteriol. 1974, 119, 889–892. [Google Scholar] [CrossRef]
- Barthelmess, I.B. Regulation of amino acid synthetic enzymes in Neurospora crassa in the presence of high concentrations of amino acids. Mol. Genet. Genom. 1986, 203, 533–537. [Google Scholar] [CrossRef]
- Paluh, J.; Orbach, M.J.; Legerton, T.L.; Yanofsky, C. The cross-pathway control gene of Neurospora crassa, cpc-1, encodes a protein similar to GCN4 of yeast and the DNA-binding domain of the oncogene v-jun-encoded protein. Proc. Natl. Acad. Sci. USA 1988, 85, 3728–3732. [Google Scholar] [CrossRef]
- Priebe, S.; Kreisel, C.; Horn, F.; Guthke, R.; Linde, J. FungiFun2: A comprehensive online resource for systematic analysis of gene lists from fungal species. Bioinformatics 2015, 31, 445–446. [Google Scholar] [CrossRef] [PubMed]
- Tükenmez, H.; Magnussen, H.M.; Kovermann, M.; Byström, A.; Wolf-Watz, M. Linkage between Fitness of Yeast Cells and Adenylate Kinase Catalysis. PLoS ONE 2016, 11, e0163115. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed]
- Moraes, C.T.; Diaz, F.; Barrientos, A. Defects in the biosynthesis of mitochondrial heme c and heme a in yeast and mammals. Biochim. Biophys. Acta (BBA) Bioenerg. 2004, 1659, 153–159. [Google Scholar] [CrossRef][Green Version]
- Hillman, G.A.; Henry, M.F. The yeast protein Mam33 functions in the assembly of the mitochondrial ribosome. J. Biol. Chem. 2019, 294, 9813–9829. [Google Scholar] [CrossRef] [PubMed]
- Salazar, D.; Zhang, L.; Degala, G.D.; Frerman, F.E. Expression and Characterization of Two Pathogenic Mutations in Human Electron Transfer Flavoprotein. J. Biol. Chem. 1997, 272, 26425–26433. [Google Scholar] [CrossRef] [PubMed]
- Margolles-Clark, E.; Tenney, K.; Bowman, E.J.; Bowman, B.J. The structure of the vacuolar ATPase in Neurospora crassa. J. Bioenerg. Biomembr. 1999, 31, 29–37. [Google Scholar] [CrossRef]
- Plesofsky, N.; Higgins, L.; Markowski, T.; Brambl, R. Glucose Starvation Alters Heat Shock Response, Leading to Death of Wild Type Cells and Survival of MAP Kinase Signaling Mutant. PLoS ONE 2016, 11, e0165980. [Google Scholar] [CrossRef]
- Reina, S.; Palermo, V.; Guarnera, A.; Guarino, F.; Messina, A.; Mazzoni, C.; De Pinto, V. Swapping of the N-terminus of VDAC1 with VDAC3 restores full activity of the channel and confers anti-aging features to the cell. FEBS Lett. 2010, 584, 2837–2844. [Google Scholar] [CrossRef]
- Madesh, M.; Hajnόczky, G. VDAC-dependent permeabilization of the outer mitochondrial membrane by superoxide induces rapid and massive cytochrome c release. J. Cell Biol. 2001, 155, 1003–1016. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; Mizrachi, D. VDAC1: From structure to cancer therapy. Front. Oncol. 2012, 2, 164. [Google Scholar] [CrossRef]
- Zhu, J.; Yu, X.; Xie, B.; Gu, X.; Zhang, Z.; Li, S. Transcriptomic profiling-based mutant screen reveals three new transcription factors mediating menadione resistance in Neurospora crassa. Fungal Biol. 2013, 117, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, M.W.; Nargund, A.M.; Haynes, C.M. Signaling the mitochondrial unfolded protein response. Biochim. Biophys. Acta (BBA) Bioenerg. 2012, 1833, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Watters, M.K.; Boersma, M.; Johnson, M.; Reyes, C.; Westrick, E.; Lindamood, E. A screen for Neurospora knockout mutants displaying growth rate dependent branch density. Fungal Biol. 2011, 115, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, C.T.; Ebbole, D.J.; Lee, B.U.; Brown, R.E.; Bourland, C.; Madi, L.; Yanofsky, C. Characterization of rco-1 of Neurospora crassa, a pleiotropic gene affecting growth and development that encodes a homolog of Tup1 of Saccharomyces cerevisiae. Mol. Cell. Biol. 1996, 16, 6218–6228. [Google Scholar] [CrossRef] [PubMed]
- Kothe, G.O.; Free, S.J. The Isolation and Characterization of nrc-1 and nrc-2, Two Genes Encoding Protein Kinases That Control Growth and Development in Neurospora crassa. Genetics 1998, 149, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, G.; Rapino, C.; Oddi, S.; Maccarrone, M. Anandamide increases swelling and reduces calcium sensitivity of mitochondria. Biochem. Biophys. Res. Commun. 2009, 388, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Vequi-Suplicy, C.C.; Lamy, M.T.; Marquezin, C.A. The New Fluorescent Membrane Probe Ahba: A Comparative Study with the Largely Used Laurdan. J. Fluoresc. 2013, 23, 479–486. [Google Scholar] [CrossRef]
- Colell, A.; García-Ruiz, C.; Lluis, J.M.; Coll, O.; Mari, M.; Fernández-Checa, J.C. Cholesterol Impairs the Adenine Nucleotide Translocator-mediated Mitochondrial Permeability Transition through Altered Membrane Fluidity. J. Biol. Chem. 2003, 278, 33928–33935. [Google Scholar] [CrossRef] [PubMed]
- Senault, C.; Yazbeck, J.; Goubern, M.; Portet, R.; Vincent, M.; Gallay, J. Relation between membrane phospholipid composition, fluidity and function in mitochondria of rat brown adipose tissue. Effect of thermal adaptation and essential fatty acid deficiency. Biochim. Biophys. Acta (BBA) Biomembr. 1990, 1023, 283–289. [Google Scholar] [CrossRef]
- Maccarrone, M.; Bellincampi, L.; Melino, G.; Agro, A.F. Cholesterol, but not its esters, triggers programmed cell death in human erythroleukemia K562 cells. JBIC J. Biol. Inorg. Chem. 1998, 253, 107–113. [Google Scholar] [CrossRef]
- Solsona-Vilarrasa, E.; Fucho, R.; Torres, S.; Núñez, S.T.; Lámbarri, N.N.; Enrich, C.; García-Ruiz, C.; Fernández-Checa, J.C. Cholesterol enrichment in liver mitochondria impairs oxidative phosphorylation and disrupts the assembly of respiratory supercomplexes. Redox Biol. 2019, 24, 101214. [Google Scholar] [CrossRef] [PubMed]
- Cantor, R.S. Lipid Composition and the Lateral Pressure Profile in Bilayers. Biophys. J. 1999, 76, 2625–2639. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Sample (Method) | Proteins Detected | Proteins More Abundant in ΔPor-1 2 | Proteins Less Abundant ΔPor-1 2 | Reference |
|---|---|---|---|---|
| S100 cytosol-enriched (1D LC-MS/MS 1) | 1031 | 74 | 96 | This work |
| Mitochondria-enriched (2D LC-MS/MS) | 867 3 | 23 | 37 | This work |
| Mitochondria-enriched (1D LC-MS/MS) | 542 | 10 | 13 | [14] |
| Mitochondria-enriched (iTRAQ) | 489 | 12 | 7 | [12] |
| Measurement | WT | ΔPor-1 |
|---|---|---|
| 1 Catalase (cytoplasm, U/mg) | 45.8 +/− 3.3 | 95.5 +/− 21.9 |
| 2 Intracellular ROS relative to WT (%) | 100 | 150 +/− 50 |
| 3 Growth rate in the absence of tunicamycin or with 2.5 μg/mL tunicamycin (cm/day) | 11.7 +/− 0.1 2.7 +/− 0.8 | 4.1 +/− 0.7 no growth |
| 4 Ergosterol content (% relative to WT) | 100 +/− 16.8 | 59.6 +/− 9.4 |
| A. More Abundant Proteins in ΔPor-1 and Cells Undergoing UPRER 1 | B. More Abundant Proteins in ΔPor-1 and Menadione-Treated Cells 2 | ||
|---|---|---|---|
| NCU05780 | glutathione S-transferase-1 | NCU05780 | glutathione S-transferase-1 |
| NCU09519 | 2,5-diketo-D-gluconic acid reductase A | NCU09559 | clock-controlled gene-9 |
| NCU10572 | short-chain oxidoreductase | NCU05770 | catalase-2 |
| NCU01272 | mitochondrial presequence protease | NCU04930 | hypothetical protein |
| NCU02549 | processing enhancing protein | NCU00355 | catalase-3 |
| NCU09559 | clock-controlled gene-9 | NCU09674 | O-methyltransferase family 3 |
| NCU09560 | superoxide dismutase | NCU02812 | uridylate kinase |
| NCU05770 | catalase-2 | NCU08402 | zinc-binding alcohol dehydrogenase |
| NCU03739 | ERP38 protein | NCU02727 | glycine cleavage system T protein |
| NCU04930 | hypothetical protein | NCU08004 | electron transfer flavoprotein alpha-subunit |
| NCU01589 | heat-shock protein 60 | NCU06974 | histidinol-phosphatase |
| NCU09674 | O-methyltransferase family 3 | NCU00685 | casein kinase I isoform delta |
| NCU03611 | chitin synthase-1 | ||
| NCU03795 | cell division control protein 12 | ||
| NCU03949 | nitropropane dioxygenase-1 | C. More abundant proteins in ΔPor-1, menadione-treated cells and cells undergoing UPRER | |
| NCU05881 | DUF500 and UBA/TS-N domain-containing protein | NCU05780 | glutathione S-transferase-1 |
| NCU10810 | mRNA-splicing protein | NCU09559 | clock-controlled gene-9 |
| NCU02727 | glycine cleavage system T protein | NCU05770 | catalase-2 |
| NCU06738 | protein transporter sec-31 | NCU04930 | hypothetical protein |
| NCU01166 | microcycle blastoconidiation | NCU09674 | O-methyltransferase family 3 |
| NCU03596 | CRAL/TRIO domain-containing protein | NCU02727 | glycine cleavage system T protein |
| NCU06974 | histidinol-phosphatase | NCU06974 | histidinol-phosphatase |
| NCU00864 | TIM-barrel enzyme family protein | NCU00685 | casein kinase I isoform delta |
| NCU09223 | protein disulfide-isomerase | ||
| NCU10360 | hypothetical protein | ||
| NCU05495 | clock-controlled gene-16 | ||
| NCU01004 | phosphatidylserine decarboxylase proenzyme | ||
| NCU00685 | casein kinase I isoform delta | ||
| NCU00350 | epoxide hydrolase | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shuvo, S.R.; Motnenko, A.; Krokhin, O.V.; Spicer, V.; Court, D.A. Proteomic Shifts Reflecting Oxidative Stress and Reduced Capacity for Protein Synthesis, and Alterations to Mitochondrial Membranes in Neurospora crassa Lacking VDAC. Microorganisms 2022, 10, 198. https://doi.org/10.3390/microorganisms10020198
Shuvo SR, Motnenko A, Krokhin OV, Spicer V, Court DA. Proteomic Shifts Reflecting Oxidative Stress and Reduced Capacity for Protein Synthesis, and Alterations to Mitochondrial Membranes in Neurospora crassa Lacking VDAC. Microorganisms. 2022; 10(2):198. https://doi.org/10.3390/microorganisms10020198
Chicago/Turabian StyleShuvo, Sabbir R., Anna Motnenko, Oleg V. Krokhin, Victor Spicer, and Deborah A. Court. 2022. "Proteomic Shifts Reflecting Oxidative Stress and Reduced Capacity for Protein Synthesis, and Alterations to Mitochondrial Membranes in Neurospora crassa Lacking VDAC" Microorganisms 10, no. 2: 198. https://doi.org/10.3390/microorganisms10020198
APA StyleShuvo, S. R., Motnenko, A., Krokhin, O. V., Spicer, V., & Court, D. A. (2022). Proteomic Shifts Reflecting Oxidative Stress and Reduced Capacity for Protein Synthesis, and Alterations to Mitochondrial Membranes in Neurospora crassa Lacking VDAC. Microorganisms, 10(2), 198. https://doi.org/10.3390/microorganisms10020198