

Monilinia fructigena Suppressing and Plant Growth Promoting Endophytic Pseudomonas spp. Bacteria Isolated from Plum
 and
and
Abstract
1. Introduction
2. Materials and Methods
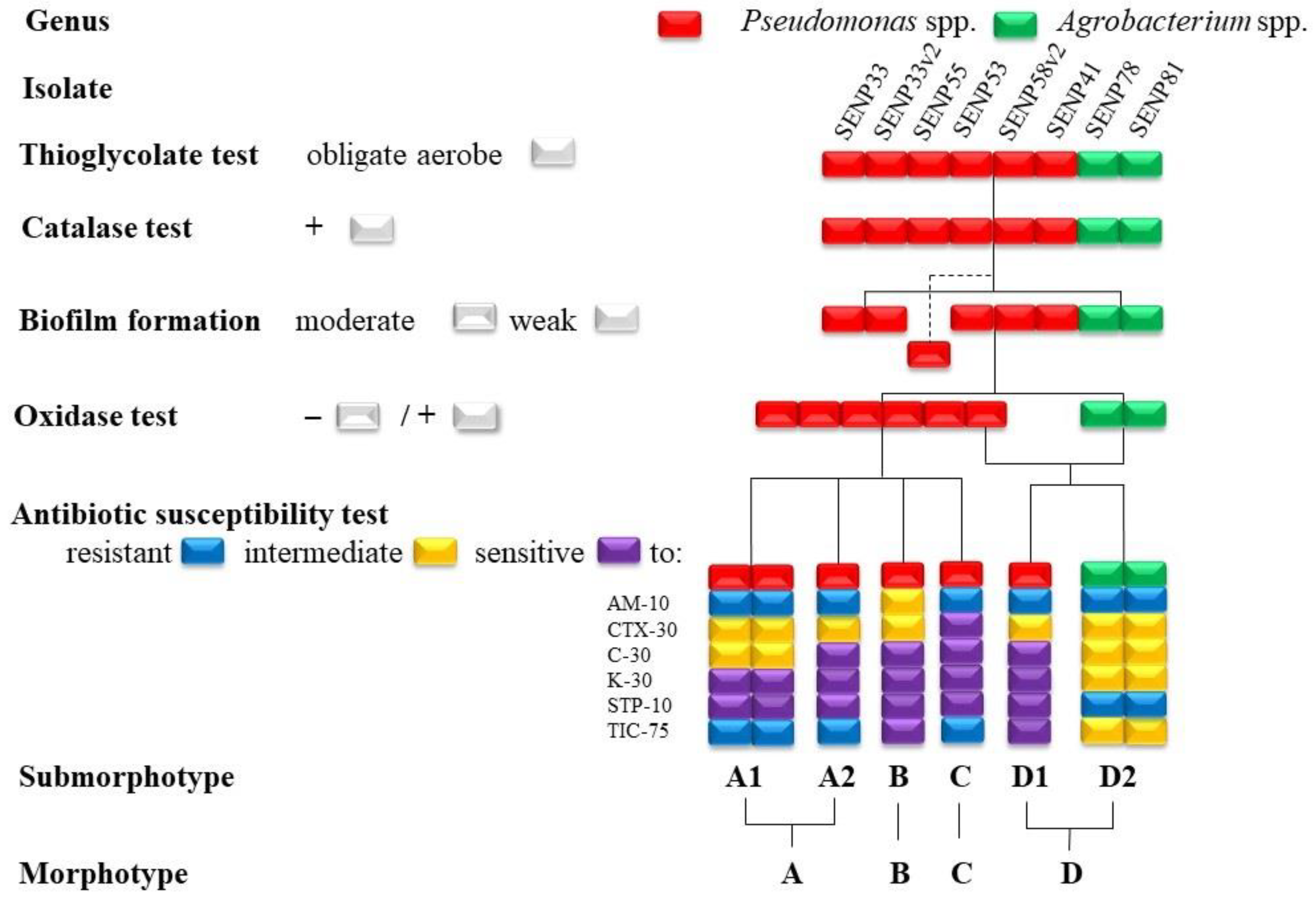
2.1. Isolation of Bacterial Endophytes
2.2. Isolation and Identification of the Fungal Pathogen Monilinia fructigena
2.3. Antagonistic Activity Test In Vitro
2.4. Identification of Potential Antagonists
2.5. Endophyte Morphotyping
2.6. Plant Growth Promoting Traits (PGPTs)
3. Results
3.1. Antagonistic Activity In Vitro
3.2. Endophytic Bacteria Morphotypes
3.3. Plant Growth Promoting Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Identification No. | Closest NCBI Match (Species) | Identity, % | Query Coverage, % | Fragment Length, bp | Accession No. * |
|---|---|---|---|---|---|
| SENP33 | Pseudomonas graminis | 99.79 | 99 | 1449 | NR_026395.1 |
| SENP33v2 | Pseudomonas graminis | 99.86 | 99 | 1400 | NR_026395.1 |
| SENP41 | Pseudomonas amygdali | 99.43 | 100 | 1393 | NR_036999.1 |
| SENP53 | Pseudomonas congelans | 100 | 99 | 1452 | NR_028985.1 |
| SENP55 | Pseudomonas graminis | 99.87 | 100 | 1399 | NR_026395.1 |
| SENP58v2 | Pseudomonas graminis | 99.79 | 99 | 1403 | NR_026395.1 |
| SENP78 | Agrobacterium fabrum | 100 | 99.86 | 1406 | NR_074266.1 |
| SENP81 | Agrobacterium fabrum | 100 | 99 | 1396 | NR_074266.1 |
| Morphotype | Isolate | Color | Form | Margin | Opacity | Appearance | Elevation | Consistency | Texture |
|---|---|---|---|---|---|---|---|---|---|
| A | Pseudomonas graminis, SENP33 | yellowish | circular | entire | translucent | glistening | raised | mucoid | smooth |
| Pseudomonas graminis, SENP33v2 | yellowish | circular | raised | mucoid | |||||
| Pseudomonas graminis, SENP55 | yellowish | circular | raised | mucoid | |||||
| B | Pseudomonas congelans, SENP53 | creamy | irregular | flat | butyrous | ||||
| C | Pseudomonas graminis, SENP58v2 | yellowish | circular | raised | butyrous | ||||
| D | Pseudomonas amygdali, SENP41 | creamy | circular | raised | butyrous | ||||
| Agrobacterium fabrum, SENP78 | creamy | circular | raised | butyrous | |||||
| Agrobacterium fabrum, SENP81 | creamy | circular | raised | butyrous |
References
- Casals, C.; Guijarro, B.; De Cal, A.; Torres, R.; Usall, J.; Perdrix, V.; Hilscher, U.; Ladurner, E. Field validation of biocontrol strategies to control brown rot on stone fruit in several European countries. Pest Manag. Sci. 2021, 77, 2502–2511. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Aksissou, W.; Lyousfi, N.; Ezrari, S.; Blenzar, A.; Tahiri, A.; Ennahli, S.; Hrustić, J.; MacLean, D.; Amiri, S. Biocontrol activity and putative mechanism of Bacillus amyloliquefaciens (SF14 and SP10), Alcaligenes faecalis ACBC1, and Pantoea agglomerans ACBP1 against brown rot disease of fruit. Microb. Pathog. 2020, 139, 103914. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, J.; Xie, J.; Deng, L.; Yao, S.; Zeng, K. Biocontrol efficacy of Pichia membranaefaciens and Kloeckera apiculata against Monilinia fructicola and their ability to induce phenylpropanoid pathway in plum fruit. Biol. Control 2019, 129, 83–91. [Google Scholar] [CrossRef]
- Grzegorczyk, M.; Żarowska, B.; Restuccia, C.; Cirvilleri, G. Postharvest biocontrol ability of killer yeasts against Monilinia fructigena and Monilinia fructicola on stone fruit. Food Microbiol. 2017, 61, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Mari, M.; Martini, C.; Guidarelli, M.; Neri, F. Postharvest biocontrol of Monilinia laxa, Monilinia fructicola and Monilinia fructigena on stone fruit by two Aureobasidium pullulans strains. Biol. Control 2012, 60, 132–140. [Google Scholar] [CrossRef]
- Miessner, S.; Stammler, G. Monilinia laxa, M. fructigena and M. fructicola: Risk estimation of resistance to QoI fungicides and Identification of Species With Cytochrome B Gene Sequences. J. Plant Dis. Prot. 2010, 117, 162–167. [Google Scholar] [CrossRef]
- Madbouly, A.K.; Abo Elyousr, K.A.M.; Ismail, I.M. Biocontrol of Monilinia fructigena, causal agent of brown rot of apple fruit, by using endophytic yeasts. Biol. Control 2020, 144, 104239. [Google Scholar] [CrossRef]
- Lahlali, R.; McHachti, O.; Radouane, N.; Ezrari, S.; Belabess, Z.; Khayi, S.; Mentag, R.; Tahiri, A.; Barka, E.A. The potential of novel bacterial isolates from natural soil for the control of brown rot disease (Monilinia fructigena) on apple fruits. Agronomy 2020, 10, 1814. [Google Scholar] [CrossRef]
- Bellamy, S.; Xu, X.; Shaw, M. Biocontrol agents to manage brown rot disease on cherry. Eur. J. Plant Pathol. 2021, 161, 493–502. [Google Scholar] [CrossRef]
- Czarnecka, M.; Żarowska, B.; Połomska, X.; Restuccia, C.; Cirvilleri, G. Role of biocontrol yeasts Debaryomyces hansenii and Wickerhamomyces anomalus in plants’ defence mechanisms against Monilinia fructicola in apple fruits. Food Microbiol. 2019, 83, 1–8. [Google Scholar] [CrossRef]
- Rungjindamai, N.; Jeffries, P.; Xu, X.M. Epidemiology and management of brown rot on stone fruit caused by Monilinia laxa. Eur. J. Plant Pathol. 2014, 140, 1–17. [Google Scholar] [CrossRef]
- Xu, X.M.; Robinson, J.D. Epidemiology of brown rot (Monilinia fructigena) on apple: Infection of fruits by conidia. Plant Pathol. 2000, 49, 201–206. [Google Scholar] [CrossRef]
- Xu, X.M.; Guerin, L.; Robinson, J.D. Effects of temperature and relative humidity on conidial germination and viability, colonization and sporulation of Monilinia fructigena. Plant Pathol. 2001, 50, 561–568. [Google Scholar] [CrossRef]
- Hrustić, J.; Mihajlović, M.; Grahovac, M.; Delibašić, G.; Tanović, B. Fungicide sensitivity, growth rate, aggressiveness and frost hardiness of Monilinia fructicola and Monilinia laxa isolates. Eur. J. Plant Pathol. 2018, 151, 389–400. [Google Scholar] [CrossRef]
- Egüen, B.; Melgarejo, P.; De Cal, A. The effect of fungicide resistance on the structure of Monilinia laxa populations in Spanish peach and nectarine orchards. Eur. J. Plant Pathol. 2016, 145, 815–827. [Google Scholar] [CrossRef]
- Larena, I.; Torres, R.; De Cal, A.; Liñán, M.; Melgarejo, P.; Domenichini, P.; Bellini, A.; Mandrin, J.F.; Lichou, J.; De Eribe, X.O.; et al. Biological control of postharvest brown rot (Monilinia spp.) of peaches by field applications of Epicoccum nigrum. Biol. Control 2005, 32, 305–310. [Google Scholar] [CrossRef]
- Aiello, D.; Restuccia, C.; Stefani, E.; Vitale, A.; Cirvilleri, G. Postharvest biocontrol ability of Pseudomonas synxantha against Monilinia fructicola and Monilinia fructigena on stone fruit. Postharvest Biol. Technol. 2019, 149, 83–89. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, J.; Zhou, Y.; Deng, L.; Yao, S.; Zeng, K. Inhibitory effect of Pichia membranaefaciens and Kloeckera apiculata against Monilinia fructicola and their biocontrol ability of brown rot in postharvest plum. Biol. Control 2017, 114, 51–58. [Google Scholar] [CrossRef]
- Martini, C.; Mari, M. Monilinia fructicola, Monilinia laxa (Monilinia Rot, Brown Rot); Elsevier: Amsterdam, The Netherlands, 2014; ISBN 9780124115682. [Google Scholar]
- Rodriguez-Pires, S.; Garcia-Companys, M.; Espeso, E.A.; Melgarejo, P.; De Cal, A. Light influence on Monilinia laxa stone fruit interaction. Plant Pathol. 2021, 70, 326–335. [Google Scholar] [CrossRef]
- Tanovic, B.; Gasic, S.; Hrustic, J.; Mihajlovic, M.; Grahovac, M.; Delibasic, G.; Stevanovic, M. Development of a thyme essential oil formulation and its effect on Monilinia ructigena. Pestic. Fitomedicina 2013, 28, 273–280. [Google Scholar] [CrossRef]
- Di Francesco, A.; Ugolini, L.; D’Aquino, S.; Pagnotta, E.; Mari, M. Biocontrol of Monilinia laxa by Aureobasidium pullulans strains: Insights on competition for nutrients and space. Int. J. Food Microbiol. 2017, 248, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K.; Prakash, A.; Johri, B.N. Induced systemic resistance (ISR) in plants: Mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, V.O.; Johnson, K.B.; Sugar, D.; Loper, J.E. Antibiosis contributes to biological control of fire blight by Pantoea agglomerans strain Eh252 in orchards. Phytopathology 2002, 92, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Itkina, D.L.; Suleimanova, A.D.; Sharipova, M.R. Pantoea brenneri AS3 and Bacillus ginsengihumi M2.11 as Potential Biocontrol and Plant Growth-Promoting Agents. Microbiology 2021, 90, 210–218. [Google Scholar] [CrossRef]
- Latz, M.A.C.; Jensen, B.; Collinge, D.B.; Jørgensen, H.J.L. Endophytic fungi as biocontrol agents: Elucidating mechanisms in disease suppression. Plant Ecol. Divers. 2018, 11, 555–567. [Google Scholar] [CrossRef]
- Glick, B.R. Beneficial plant-bacterial interactions. In Biocontrol of Bacteria and Fungi; Springer: Cham, Switzerland, 2020; pp. 181–232. ISBN 9783030443689. [Google Scholar]
- Schena, L.; Nigro, F.; Pentimone, I.; Ligorio, A.; Ippolito, A. Control of postharvest rots of sweet cherries and table grapes with endophytic isolates of Aureobasidium pullulans. Postharvest Biol. Technol. 2003, 30, 209–220. [Google Scholar] [CrossRef]
- Hong, C.E.; Park, J.M. Endophytic bacteria as biocontrol agents against plant pathogens: Current state-of-the-art. Plant Biotechnol. Rep. 2016, 10, 353–357. [Google Scholar] [CrossRef]
- Vaitiekūnaitė, D.; Kuusienė, S.; Beniušytė, E. Oak (Quercus robur) Associated Endophytic Paenibacillus sp. Promotes Poplar (Populus spp.) Root Growth In Vitro. Microorganisms 2021, 9, 1151. [Google Scholar] [CrossRef]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: A treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef]
- White, J.F.; Torres, M.S. Is plant endophyte-mediated defensive mutualism the result of oxidative stress protection? Physiol. Plant. 2010, 138, 440–446. [Google Scholar] [CrossRef]
- Liu, H.; Carvalhais, L.C.; Crawford, M.; Singh, E.; Dennis, P.G.; Pieterse, C.M.J.; Schenk, P.M. Inner plant values: Diversity, colonization and benefits from endophytic bacteria. Front. Microbiol. 2017, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Boyle, C. What are Endophytes? In Microbial Root Endophytes. Soil Biology; Springer: Berlin/Heidelberg, Germany, 2006; Volume 9, pp. 1–14. ISBN 978-3-540-33525-2. [Google Scholar]
- Hsieh, T.F.; Huang, H.C.; Erickson, R.S. Biological control of bacterial wilt of bean using a bacterial endophyte, Pantoea agglomerans. J. Phytopathol. 2005, 153, 608–614. [Google Scholar] [CrossRef]
- Ren, J.H.; Li, H.; Wang, Y.F.; Ye, J.R.; Yan, A.Q.; Wu, X.Q. Biocontrol potential of an endophytic Bacillus pumilus JK-SX001 against poplar canker. Biol. Control 2013, 67, 421–430. [Google Scholar] [CrossRef]
- Blumenstein, K.; Albrectsen, B.R.; Martín, J.A.; Hultberg, M.; Sieber, T.N.; Helander, M.; Witzell, J. Nutritional niche overlap potentiates the use of endophytes in biocontrol of a tree disease. BioControl 2015, 60, 655–667. [Google Scholar] [CrossRef]
- Strobel, G. The Emergence of Endophytic Microbes and Their Biological Promise. J. Fungi 2018, 4, 57. [Google Scholar] [CrossRef]
- Johnston-Monje, D.; Raizada, M.N. Conservation and diversity of seed associated endophytes in Zea across boundaries of evolution, ethnography and ecology. PLoS ONE 2011, 6, e20396. [Google Scholar] [CrossRef]
- Ait Bahadou, S.; Ouijja, A.; Karfach, A.; Tahiri, A.; Lahlali, R. New potential bacterial antagonists for the biocontrol of fire blight disease (Erwinia amylovora) in Morocco. Microb. Pathog. 2018, 117, 7–15. [Google Scholar] [CrossRef]
- Chang, H.; Yang, H.; Han, T.; Wang, F.; Liu, Y. Study on the optimal antagonistic effect of a bacterial complex against Monilinia fructicola in peach. Open Life Sci. 2020, 15, 890–901. [Google Scholar] [CrossRef]
- Lyousfi, N.; Letrib, C.; Legrifi, I.; Blenzar, A.; El Khetabi, A.; El Hamss, H.; Belabess, Z.; Barka, E.A.; Lahlali, R. Combination of Sodium Bicarbonate (SBC) with Bacterial Antagonists for the Control of Brown Rot Disease of Fruit. J. Fungi 2022, 8, 636. [Google Scholar] [CrossRef]
- Janakiev, T.; Unković, N.; Dimkić, I.; Grbić, M.L.; Stević, T.; Stanković, S.; Berić, T. Susceptibility of Serbian plum cultivars to indigenous bacterial and Monilinia laxa isolates. Bot. Serbica 2020, 44, 203–210. [Google Scholar] [CrossRef]
- Janakiev, T.; Dimkić, I.; Unković, N.; Ljaljević Grbić, M.; Opsenica, D.; Gašić, U.; Stanković, S.; Berić, T. Phyllosphere Fungal Communities of Plum and Antifungal Activity of Indigenous Phenazine-Producing Pseudomonas synxantha against Monilinia laxa. Front. Microbiol. 2019, 10, 2287. [Google Scholar] [CrossRef] [PubMed]
- Janisiewicz, W.J.; Jurick, W.M.; Vico, I.; Peter, K.A.; Buyer, J.S. Culturable bacteria from plum fruit surfaces and their potential for controlling brown rot after harvest. Postharvest Biol. Technol. 2013, 76, 145–151. [Google Scholar] [CrossRef]
- Palmieri, D.; Ianiri, G.; Conte, T.; Castoria, R.; Lima, G.; De Curtis, F. Influence of Biocontrol and Integrated Strategies and Treatment Timing on Plum Brown Rot Incidence and Fungicide Residues in Fruits. Agriculture 2022, 12, 1656. [Google Scholar] [CrossRef]
- Sharma, B. Biotechnology in Agriculture and Forestry. 17. High-Tech and Micropropagation I; Springer Berlin, Heidelberg, 1992; ISBN 9783642764172.
- Kose, M.S.H.; Dogan, M.; Sadi, G. Surface sterilization of Staurogyne repens (Nees) Kuntze with hydrogen peroxide. Bull. Biotechnol. 2020, 1, 6–9. [Google Scholar]
- Davoudpour, Y.; Schmidt, M.; Calabrese, F.; Richnow, H.H.; Musat, N. High resolution microscopy to evaluate the efficiency of surface sterilization of Zea Mays seeds. PLoS ONE 2020, 15, 1–21. [Google Scholar] [CrossRef]
- Merck. Explant Sterilization–Plant Tissue Culture Protocol. Available online: https://www.sigmaaldrich.com/LT/en/technical-documents/protocol/cell-culture-and-cell-culture-analysis/plant-tissue-culture/explant-sterilization (accessed on 25 January 2022).
- Tomaszewska-Sowa, M.; Figas, A.; Keutgen, N.; Keutgen, A.J. Establishing an efficient explant superficial sterilization protocol for in vitro micropropagation of bear’s garlic (Allium ursinum L.). Herba Pol. 2015, 61, 66–77. [Google Scholar] [CrossRef]
- Miliūtė, I.; Buzaitė, O. IAA production and other plant growth promoting traits of endophytic bacteria from apple tree. Biologija 2011, 57, 98–102. [Google Scholar] [CrossRef]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef]
- Amiri, A.; Holb, I.J.; Schnabel, G. A New Selective Medium for the Recovery and Enumeration of Monilinia fructicola, M. fructigena, and M. laxa from Stone Fruits. Mycology 2009, 99, 1199–1208. [Google Scholar] [CrossRef]
- Côté, M.-J.; Tardif, M.-C.; Meldrum, A.J. Identification of Monilinia fructigena, M. fructicola, M. laxa, and Monilia polystroma on Inoculated and Naturally Infected Fruit Using Multiplex PCR. Plant Dis. 2004, 88, 1219–1225. [Google Scholar] [CrossRef]
- Ulrich, K.; Becker, R.; Behrendt, U.; Kube, M.; Ulrich, A. A Comparative Analysis of Ash Leaf-Colonizing Bacterial Communities Identifies Putative Antagonists of Hymenoscyphus fraxineus. Front. Microbiol. 2020, 11, 966. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Heredia, D.; Henríquez-Castillo, C.; Donoso, R.; Lavin, P.; Pavlov, M.S.; FRanchi, O.; Campos, J.L. Complete Genome Sequence of Pseudomonas chilensis Strain ABC1, Isolated from Soil. Microbiol. Resour. Announc. 2020, 9, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Bueno-Gonzalez, V.; Brady, C.; Denman, S.; Allainguillaume, J.; Arnold, D. Pseudomonas kirkiae sp. Nov., a novel species isolated from oak in the united kingdom, and phylogenetic considerations of the genera Pseudomonas, Azotobacter and Azomonas. Int. J. Syst. Evol. Microbiol. 2020, 70, 2426–2434. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: Pluses, perils, and pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef]
- Vaitiekūnaitė, D.; Snitka, V. Differentiation of Closely Related Oak-Associated Gram-Negative Bacteria by Label-Free Surface Enhanced Raman Spectroscopy (SERS). Microorganisms 2021, 9, 1969. [Google Scholar] [CrossRef]
- Bergey, D.H. Bergey’s Manual of Systematic Bacteriology-Vol 4: Bacteroidetes; Springer: New York, NY, USA, 2010. [Google Scholar] [CrossRef]
- Vaitiekūnaitė, D.; Bružaitė, I.; Snitka, V. Endophytes from blueberry (Vaccinium sp.) fruit: Characterization of yeast and bacteria via label-free surface-enhanced Raman spectroscopy (SERS). Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2022, 275, 121158. [Google Scholar] [CrossRef]
- Setiawati, T.C.; Mutmainnah, L. Solubilization of Potassium Containing Mineral by Microorganisms From Sugarcane Rhizosphere. Agric. Agric. Sci. Procedia 2016, 9, 108–117. [Google Scholar] [CrossRef]
- Fatharani, R.; Rahayu, Y.S. Isolation and Characterization of Potassium-Solubilizing Bacteria from Paddy Rhizosphere (Oryza sativa L.). J. Phys. Conf. Ser. 2018, 1108, 012105. [Google Scholar] [CrossRef]
- Tiwari, G.; Duraivadivel, P.; Sharma, S.; Hariprasad, P. 1-Aminocyclopropane-1-carboxylic acid deaminase producing beneficial rhizobacteria ameliorate the biomass characters of Panicum maximum Jacq. by mitigating drought and salt stress. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Santoyo, G.; del Orozco-Mosqueda, M.C.; Govindappa, M. Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: A review. Biocontrol Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Tamošiūnė, I.; Stanienė, G.; Haimi, P.; Stanys, V.; Rugienius, R.; Baniulis, D. Endophytic Bacillus and Pseudomonas spp. Modulate Apple Shoot Growth, Cellular Redox Balance, and Protein Expression Under in Vitro Conditions. Front. Plant Sci. 2018, 9, 889. [Google Scholar] [CrossRef] [PubMed]
- Mažeikienė, I.; Frercks, B.; Burokienė, D.; Mačionienė, I.; Šalaševičienė, A. Endophytic community composition and genetic-enzymatic features of cultivable bacteria in Vaccinium myrtillus L. In forests of the Baltic-Nordic region. Forests 2021, 12, 1647. [Google Scholar] [CrossRef]
- Rojas-Solís, D.; Zetter-Salmón, E.; Contreras-Pérez, M.; Rocha-Granados, M.d.C.; Macías-Rodríguez, L.; Santoyo, G. Pseudomonas stutzeri E25 and Stenotrophomonas maltophilia CR71 endophytes produce antifungal volatile organic compounds and exhibit additive plant growth-promoting effects. Biocatal. Agric. Biotechnol. 2018, 13, 46–52. [Google Scholar] [CrossRef]
- Marag, P.S.; Suman, A. Growth stage and tissue specific colonization of endophytic bacteria having plant growth promoting traits in hybrid and composite maize (Zea mays L.). Microbiol. Res. 2018, 214, 101–113. [Google Scholar] [CrossRef]
- Gillani, R.A.; Shenaz, N.; Matloob, S.; Haq, F.; Ngah, W.S.W.; Nasim, W.; Munis, M.F.H.; Rehman, A.; Chaudhary, H.J. Biosorption of Cr(III) and Pb(II) by endophytic Agrobacterium tumefaciens 12b3: Equilibrium and kinetic studies. Desalin. Water Treat. 2017, 67, 206–214. [Google Scholar] [CrossRef]
- Yan, J.; Li, Y.; Han, X.Z.; Chen, W.F.; Zou, W.X.; Xie, Z.; Li, M. Agrobacterium deltaense sp. nov., an endophytic bacteria isolated from nodule of Sesbania cannabina. Arch. Microbiol. 2017, 199, 1003–1009. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar, A.; Pandey, K.D.; Roy, B.K. Isolation and characterization of bacterial endophytes from the roots of Cassia tora L. Ann. Microbiol. 2015, 65, 1391–1399. [Google Scholar] [CrossRef]
- Aserse, A.A.; Räsänen, L.A.; Aseffa, F.; Hailemariam, A.; Lindström, K. Diversity of sporadic symbionts and nonsymbiotic endophytic bacteria isolated from nodules of woody, shrub, and food legumes in Ethiopia. Appl. Microbiol. Biotechnol. 2013, 97, 10117–10134. [Google Scholar] [CrossRef]
- Mina, D.; Pereira, J.A.; Lino-Neto, T.; Baptista, P. Epiphytic and Endophytic Bacteria on Olive Tree Phyllosphere: Exploring Tissue and Cultivar Effect. Microb. Ecol. 2020, 80, 145–157. [Google Scholar] [CrossRef]
- Beiranvand, M.; Amin, M.; Hashemi-Shahraki, A.; Romani, B.; Yaghoubi, S.; Sadeghi, P. Antimicrobial activity of endophytic bacterial populations isolated from medical plants of Iran. Iran. J. Microbiol. 2017, 9, 11–18. [Google Scholar] [PubMed]
- Padda, K.P.; Puri, A.; Chanway, C. Endophytic nitrogen fixation-a possible “hidden” source of nitrogen for lodgepole pine trees growing at unreclaimed gravel mining sites. FEMS Microbiol. Ecol. 2019, 95, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fones, H.N.; McCurrach, H.; Mithani, A.; Smith, J.A.C.; Preston, G.M. Local adaptation is associated with zinc tolerance in Pseudomonas endophytes of the metal-hyperaccumulator plant Noccaea caerulescens. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160648. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, M.B.; López, M.L.; Echeverría, G.; Viñas, I.; Zudaire, L.; Abadias, M. Evaluation of biocontrol capacity of Pseudomonas graminis CPA-7 against foodborne pathogens on fresh-cut pear and its effect on fruit volatile compounds. Food Microbiol. 2018, 76, 226–236. [Google Scholar] [CrossRef]
- Collazo, C.; Abadias, M.; Aguiló-Aguayo, I.; Alegre, I.; Chenoll, E.; Viñas, I. Studies on the biocontrol mechanisms of Pseudomonas graminis strain CPA-7 against food-borne pathogens in vitro and on fresh-cut melon. LWT-Food Sci. Technol. 2017, 85, 301–308. [Google Scholar] [CrossRef]
- Ramírez-Zapata, D.; Ramos, C.; Aguilera, S.; Bardaji, L.; Martínez-Gil, M.; Murillo, J. Two homologues of the global regulator csr/rsm redundantly control phaseolotoxin biosynthesis and virulence in the plant pathogen Pseudomonas amygdali pv. Phaseolicola 1448a. Microorganisms 2020, 8, 1536. [Google Scholar] [CrossRef]
- Iacobellis, N.S.; Evidente, A.; Surico, G.; Sisto, A.; Gammaldi, G. Production of Phytohormones by Pseudomonas amygdali and their Role in the Hyperplastic Bacterial Canker of Almond. J. Phytopathol. 1990, 129, 177–186. [Google Scholar] [CrossRef]
- Pokojska-Burdziej, A.; Strzelczyk, E.; Dahm, H.; Li, C.Y. Effect of Endophytic Bacterium Pseudomonas fulva on Growth of Pine Seedlings (Pinus sylvestris), Formation of Mycorrhizae and Protection Against Pathogens. Phytopathol. Pol. 2004, 32, 33–47. [Google Scholar]
- Islam, M.A.; Nain, Z.; Alam, M.K.; Banu, N.A.; Islam, M.R. In vitro study of biocontrol potential of rhizospheric Pseudomonas aeruginosa against Fusarium oxysporum f. Sp. cucumerinum. Egypt. J. Biol. Pest Control 2018, 28, 1–11. [Google Scholar] [CrossRef]
- Pellicciaro, M.; Lione, G.; Giordano, L.; Gonthier, P. Biocontrol potential of Pseudomonas protegens against Heterobasidion species attacking conifers in Europe. Biol. Control 2021, 157, 104583. [Google Scholar] [CrossRef]
- Khosro, M.; Yousef, S. Bacterial Biofertilizers for Sustainable Crop Production: A Review. ARPN J. Agric. Bol. Sci. 2012, 7, 307–316. [Google Scholar]
- Hu, J.; Wei, Z.; Weidner, S.; Friman, V.P.; Xu, Y.C.; Shen, Q.R.; Jousset, A. Probiotic Pseudomonas communities enhance plant growth and nutrient assimilation via diversity-mediated ecosystem functioning. Soil Biol. Biochem. 2017, 113, 122–129. [Google Scholar] [CrossRef]
- Wallace, R.L.; Hirkala, D.L.; Nelson, L.M. Mechanisms of action of three isolates of Pseudomonas fluorescens active against postharvest grey mold decay of apple during commercial storage. Biol. Control 2018, 117, 13–20. [Google Scholar] [CrossRef]
- Escobar, M.A.; Dandekar, A.M. Agrobacterium tumefaciens as an agent of disease. Trends Plant Sci. 2003, 8, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Huo, N.; Gu, Y.; McCue, K.F.; Alabed, D.; Thomson, J.G. Complete Genome Sequence of Agrobacterium fabrum Strain 1D159. Microbiol. Resour. Announc. 2019, 8, 59–60. [Google Scholar] [CrossRef]
- Penyalver, R.; Vicedo, B.; López, M.M. Use of the genetically engineered Agrobacterium strain K1026 for biological control of crown gall. Eur. J. Plant Pathol. 2000, 106, 801–810. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, Y.; Liu, B.; Zhang, Z.; Yue, T. Biocontrol activity and patulin-removal effects of Bacillus subtilis Rhodobacter sphaeroides and Agrobacterium tumefaciens against Penicillium expansum. J. Appl. Microbiol. 2016, 121, 1384–1393. [Google Scholar] [CrossRef]




| Method | Hydrogen Peroxide [47] | Sodium Hypochlorite [48] | Ethanol [49] | Mercuric Chloride [47,50] | Bleach [51] | Combined Method (Modified [52]) | |
|---|---|---|---|---|---|---|---|
| Step | |||||||
| 1. | Sterilization agent, concentra-tion, treatment duration | H2O2, 3%, 10 min | NaClO, 5%, 5 min | C2H5OH, 70%, 3 min | HgCl2, 0,1%, 10 min | Bleach, 5 min | C2H5OH, 70%, 5 min; H2O2, 3%, 20 min; C2H5OH, 70%, 1 min |
| 2. | Washing with sterile distilled water | 4 times, 2 min | Once, 10 min | 4 times, 1 min | 7 times, 1 min | 4 times, 1 min | 5 times, 1 min |
| 3. | Macerate buds in 5 mL of sterile distilled water for 30 min [49] | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolytaitė, A.; Vaitiekūnaitė, D.; Antanynienė, R.; Baniulis, D.; Frercks, B. Monilinia fructigena Suppressing and Plant Growth Promoting Endophytic Pseudomonas spp. Bacteria Isolated from Plum. Microorganisms 2022, 10, 2402. https://doi.org/10.3390/microorganisms10122402
Kolytaitė A, Vaitiekūnaitė D, Antanynienė R, Baniulis D, Frercks B. Monilinia fructigena Suppressing and Plant Growth Promoting Endophytic Pseudomonas spp. Bacteria Isolated from Plum. Microorganisms. 2022; 10(12):2402. https://doi.org/10.3390/microorganisms10122402
Chicago/Turabian StyleKolytaitė, Augustina, Dorotėja Vaitiekūnaitė, Raminta Antanynienė, Danas Baniulis, and Birutė Frercks. 2022. "Monilinia fructigena Suppressing and Plant Growth Promoting Endophytic Pseudomonas spp. Bacteria Isolated from Plum" Microorganisms 10, no. 12: 2402. https://doi.org/10.3390/microorganisms10122402
APA StyleKolytaitė, A., Vaitiekūnaitė, D., Antanynienė, R., Baniulis, D., & Frercks, B. (2022). Monilinia fructigena Suppressing and Plant Growth Promoting Endophytic Pseudomonas spp. Bacteria Isolated from Plum. Microorganisms, 10(12), 2402. https://doi.org/10.3390/microorganisms10122402