


Improvement of the Nutraceutical Profile of Brewer’s Spent Grain after Treatment with Trametes versicolor

 ,
,  ,
,  , ,
, ,  and
and
Abstract

1. Introduction
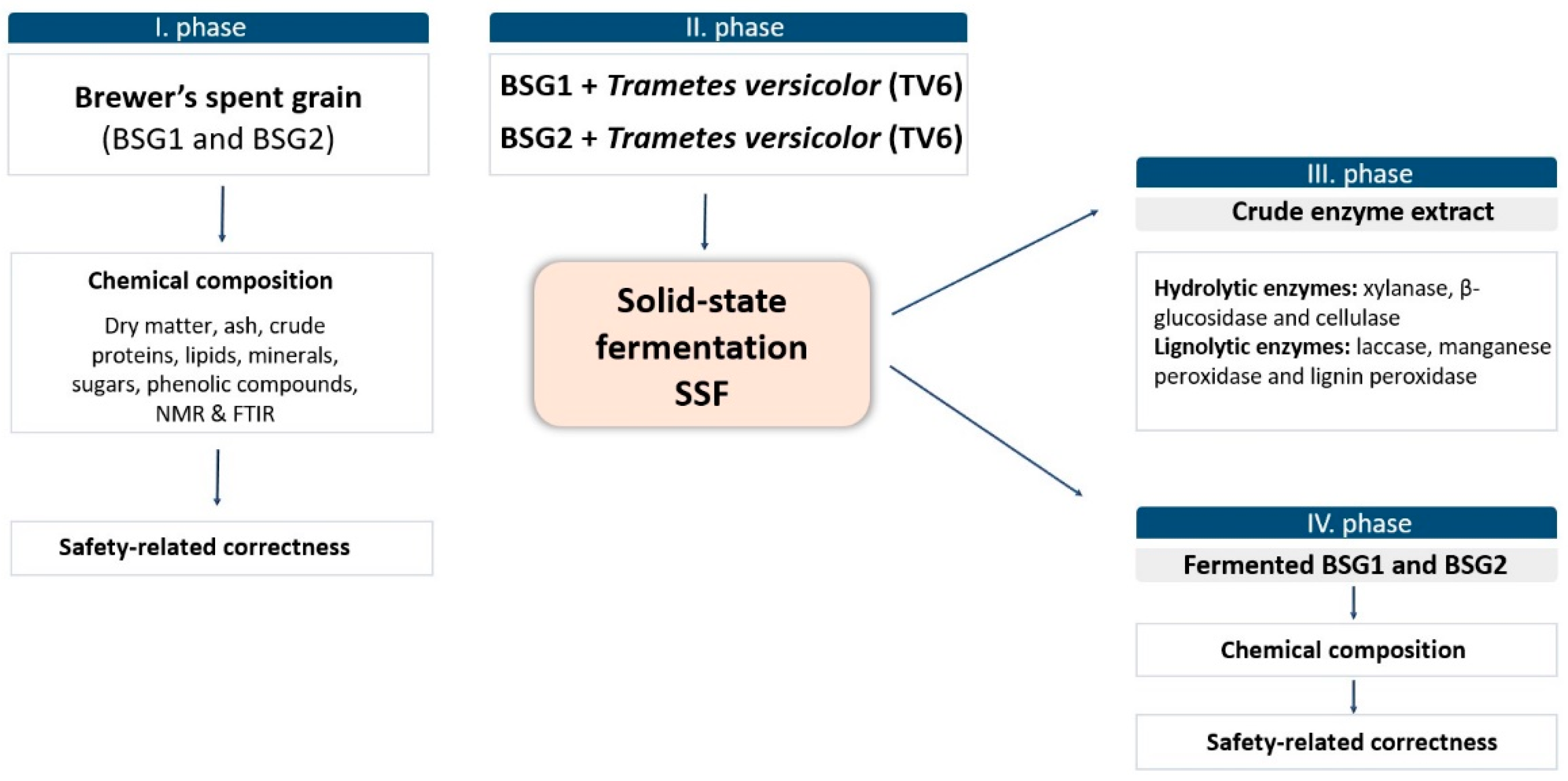
2. Materials and Methods
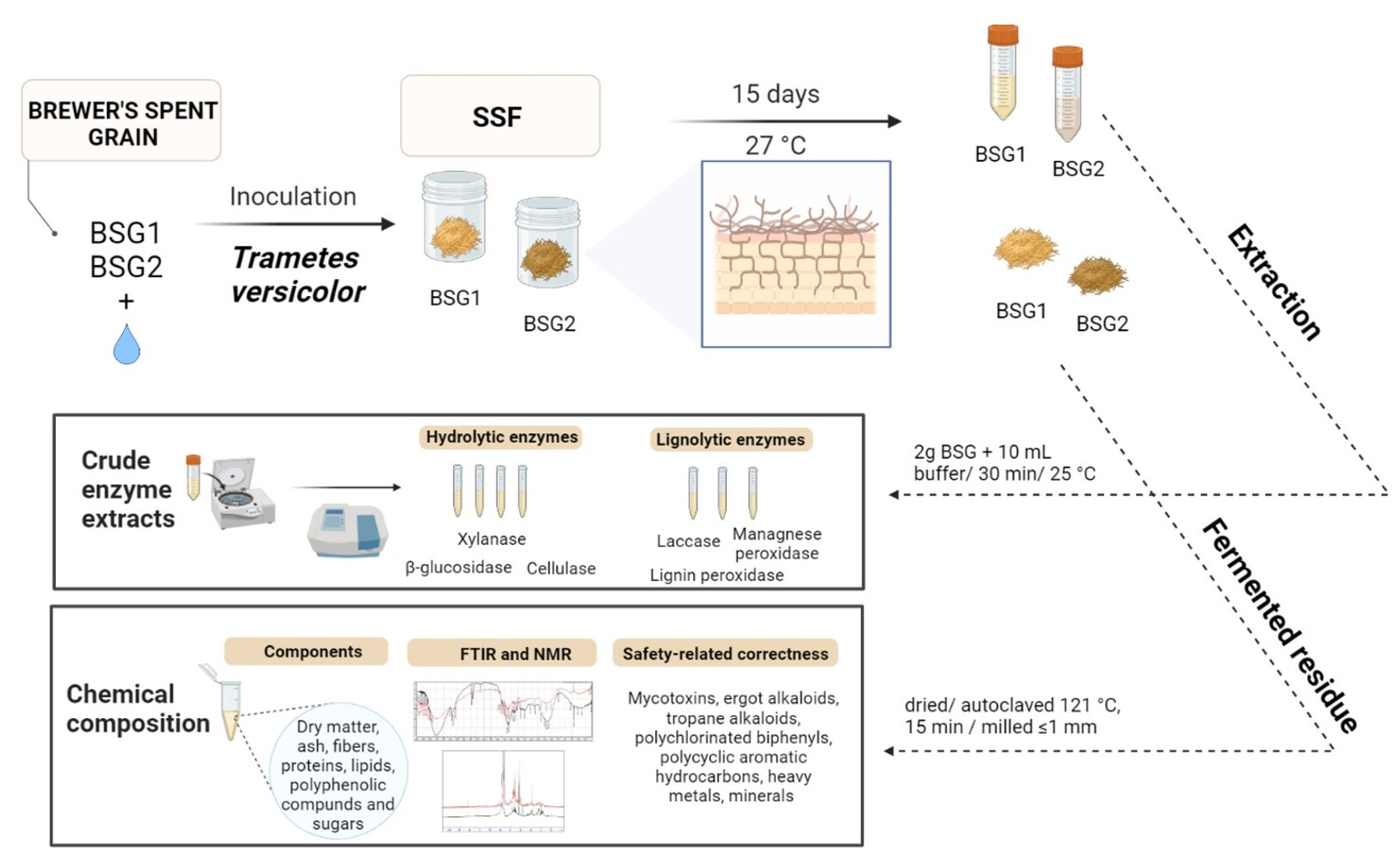
2.1. Substrates and Microorganisms
2.2. Biological Treatment of Brewer’s Spent Grain by Trametes versicolor
2.3. Enzyme Activities Measurements
2.3.1. Preparation of Crude Enzyme Extract
2.3.2. Measurements of Enzyme Activities
2.3.3. pH Measurement
2.4. Chemical Composition of BSG
2.4.1. Dry Matter Content
2.4.2. Ash Content
2.4.3. Crude Proteins
2.4.4. Lipids
2.4.5. Neutral Detergent Fibers (NDF), Acid Detergent Fibers (ADF), and Acid Detergent Lignin (ADL)
2.4.6. Content of Total Reducing Sugars
2.4.7. Content of Total Phenolic Compounds, Total Flavonoids and Total Extractable Proanthocyanidins and Individual Phenolic Compounds
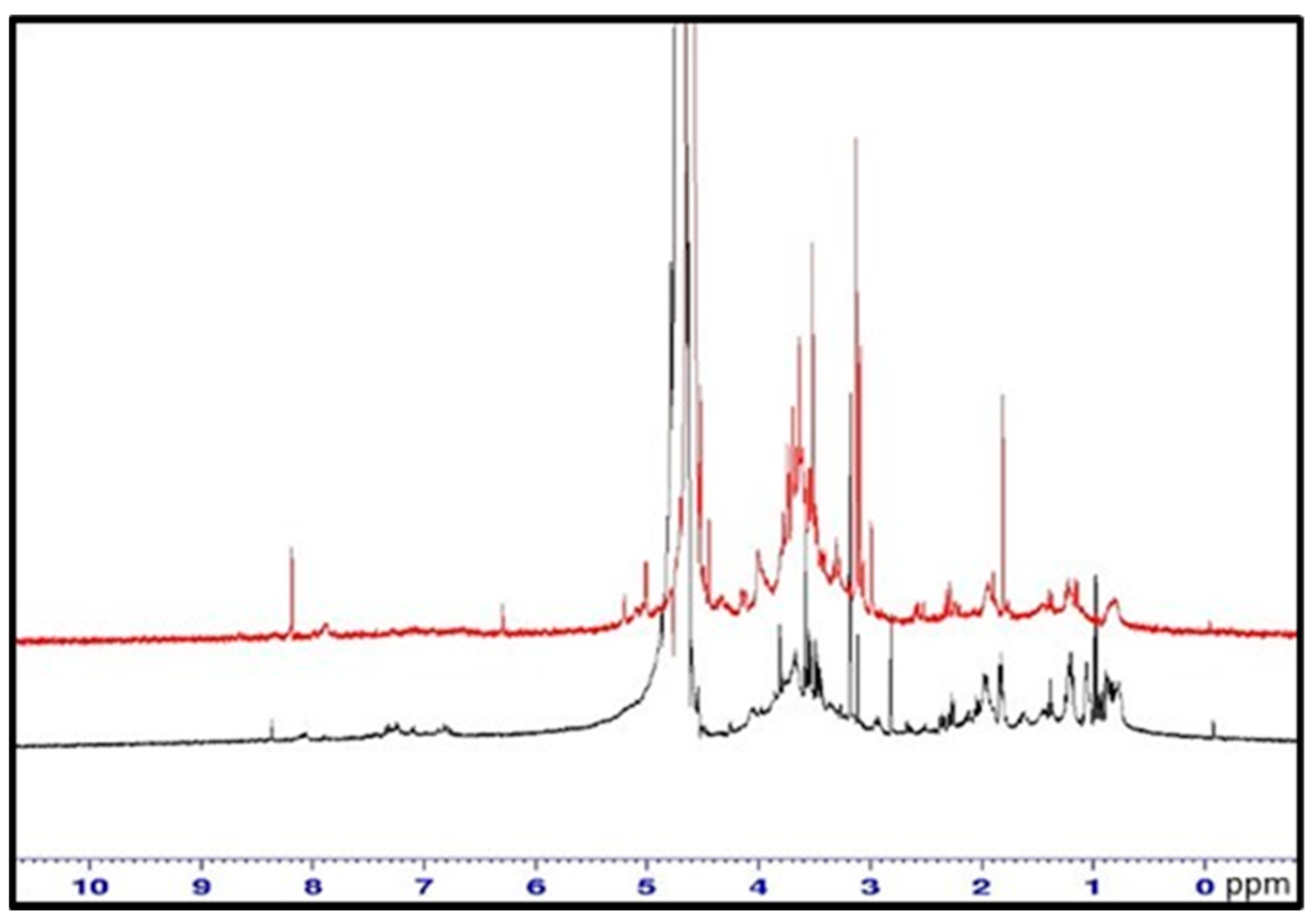
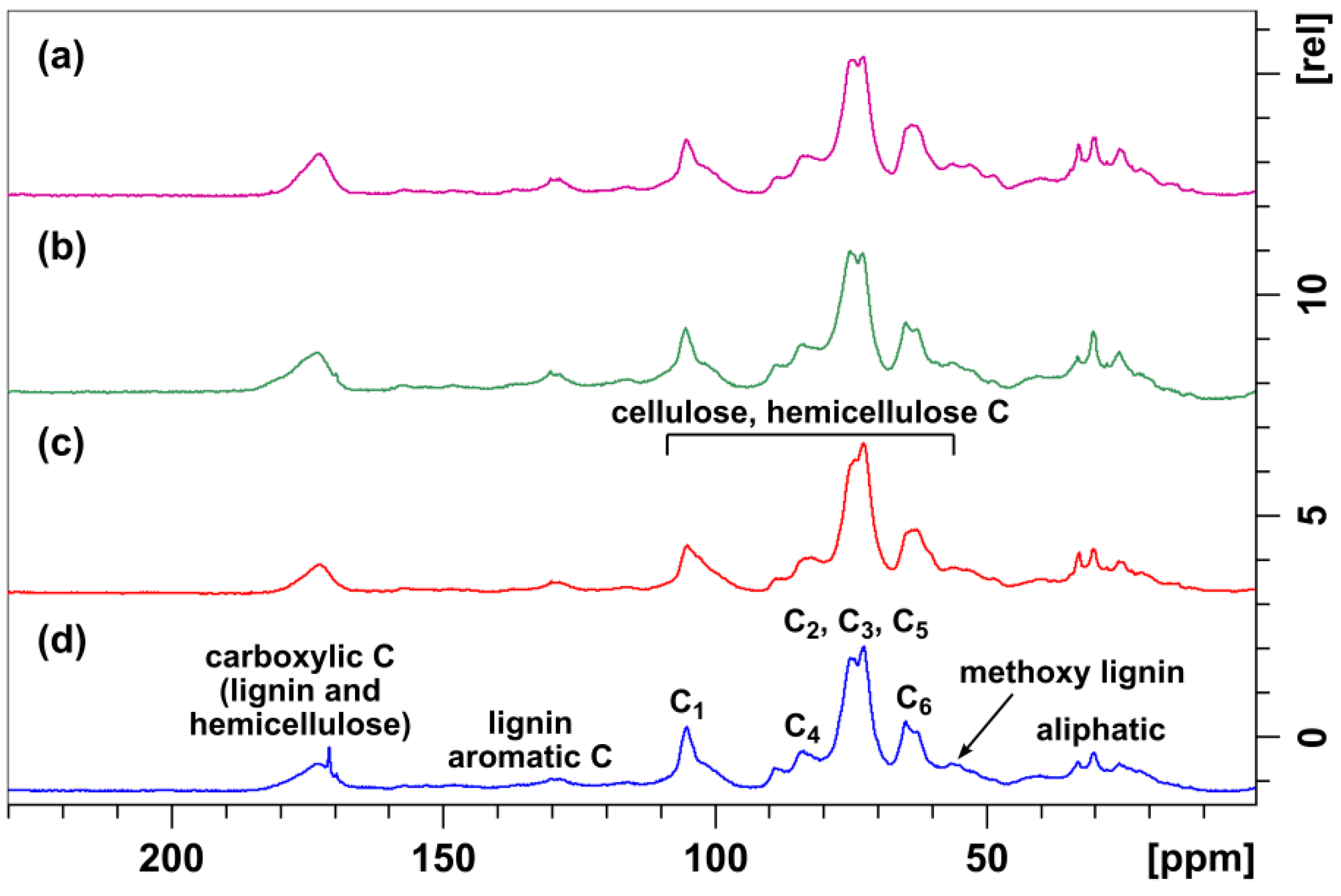
2.4.8. FTIR and NMR Spectra
2.4.9. Analysis and Evaluation of the Safety-Related Correctness of BSG for Feed
3. Results and Discussion
3.1. Cultivation of Trametes versicolor on Brewer’s Spent Grain
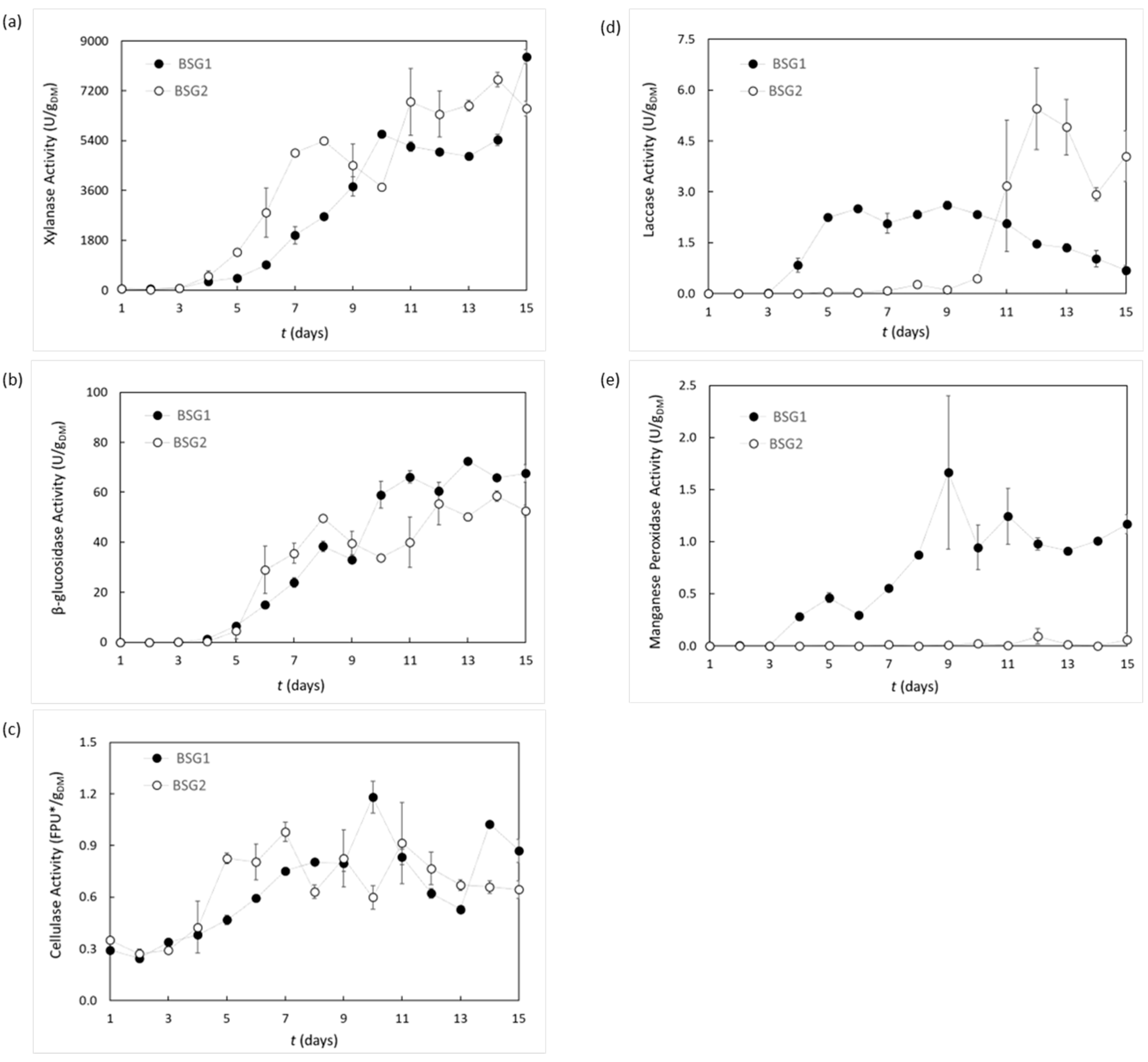
3.2. Trametes versicolor Enzymatic Activities during Cultivation on Brewer’s Spent Grain
3.3. Chemical Composition of Brewer’s Spent Grain
3.3.1. Chemical Analysis of the Fibers, Proteins, Lipids and Ash Content
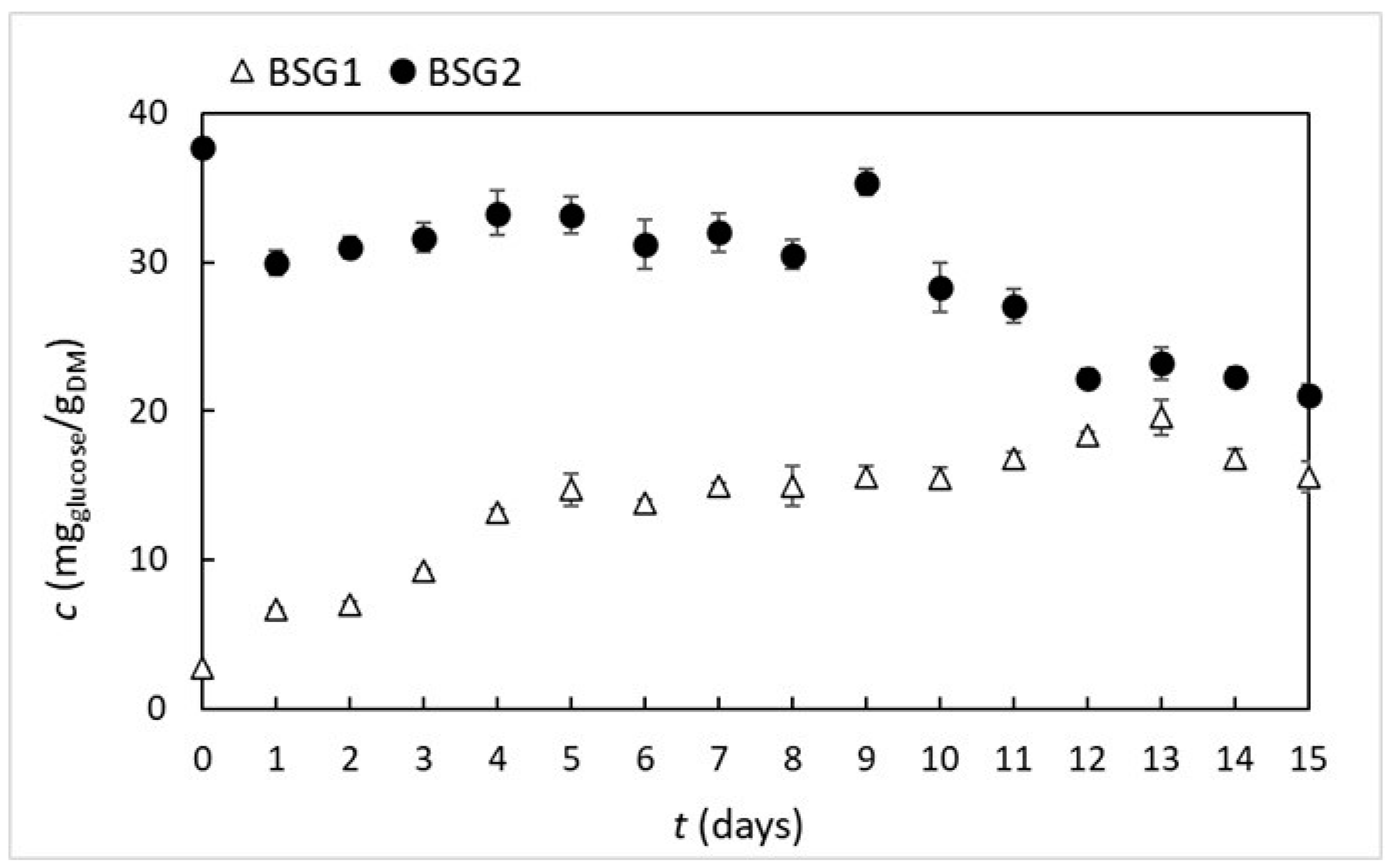
3.3.2. Total Reducing Sugars
3.3.3. Total and Individual Phenolic Compounds, Total Flavonoids, and Total Extractable Proanthocyanidins
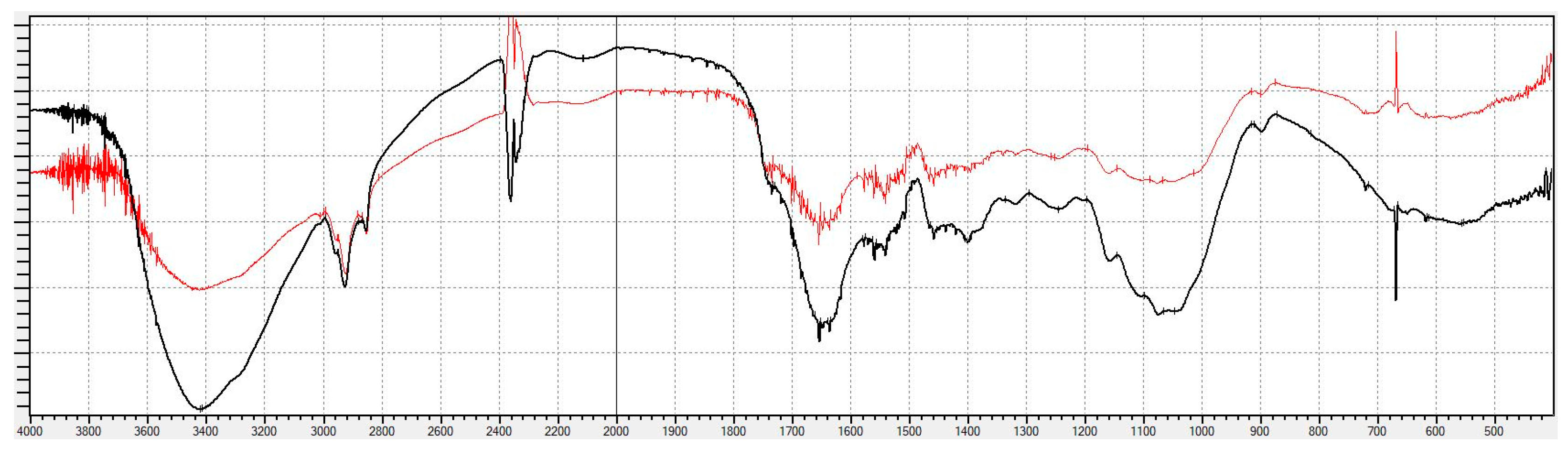
3.3.4. FTIR and NMR
3.3.5. Analysis and Evaluation of the Safety-Related Correctness of BSG for Feed
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zeko-Pivač, A.; Tišma, M.; Žnidaršič-Plazl, P.; Kulisic, B.; Sakellaris, G.; Hao, J.; Planinić, M. The Potential of Brewer’s Spent Grain in the Circular Bioeconomy: State of the Art and Future Perspectives. Front. Bioeng. Biotechnol. 2022, 10, 870744. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.T.; Ton, N.M.N.; Le, N.D.D.; Le, V.V.M. Addition of brewing spent grains from malt and rice adjunct to the formulation of high fiber biscuit: Effects of particle size of brewing spent grains on the product quality. Food Res. 2020, 4, 1480–1486. [Google Scholar] [CrossRef]
- Yitayew, T.; Moges, D.; Satheesh, N. Effect of Brewery Spent Grain Level and Fermentation Time on the Quality of Bread. Int. J. Food Sci. 2022, 2022, 8704684. [Google Scholar] [CrossRef]
- Eliopoulos, C.; Arapoglou, D.; Chorianopoulos, N.; Markou, G.; Haroutounian, S.A. Conversion of brewers’ spent grain into proteinaceous animal feed using solid state fermentation. Environ. Sci. Pollut. Res. 2022, 29, 29562–29569. [Google Scholar] [CrossRef] [PubMed]
- Parchami, M.; Ferreira, J.; Taherzadeh, M. Brewing process development by integration of edible filamentous fungi to upgrade the quality of Brewer’s spent grain (BSG). BioResources 2021, 16, 1686–1701. [Google Scholar] [CrossRef]
- Heffernan, S.M.; Conway, G.E. Nutraceutical Alternatives to Pharmaceutical Analgesics in Osteoarthritis. In Pain Management—Practices, Novel Therapies and Bioactives; Waisundara, V.Y., Banjari, I., Balkić, J., Eds.; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Alexander, W. Hypertension: Is it time to replace drugs with nutrition and nutraceuticals? Pharm. Ther. 2014, 39, 291. [Google Scholar]
- Cano y Postigo, L.O.; Jacobo-Velázquez, D.A.; Guajardo-Flores, D.; Garcia Amezquita, L.E.; García-Cayuela, T. Solid-state fermentation for enhancing the nutraceutical content of agrifood by-products: Recent advances and its industrial feasibility. Food Biosci. 2021, 41, 100926. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; Abd El-Hack, M.E.; Barkat, R.A.; Gabr, A.A.; Foda, A.M.; Noreldin, A.E.; Khafaga, A.F.; El-Sabrout, K.; et al. Potential role of important nutraceuticals in poultry performance and health—A comprehensive review. Res. Vet. Sci. 2021, 137, 9–29. [Google Scholar] [CrossRef]
- Costagliola, G.; Nuzzi, G.; Spada, E.; Comberiati, P.; Verduci, E.; Peroni, D.G. Nutraceuticals in Viral Infections: An Overview of the Immunomodulating Properties. Nutrients 2021, 13, 2410. [Google Scholar] [CrossRef] [PubMed]
- Taroncher, M.; Vila-Donat, P.; Tolosa, J.; Ruiz, M.J.; Rodríguez-Carrasco, Y. Biological activity and toxicity of plant nutraceuticals: An overview. Curr. Opin. Food Sci. 2021, 42, 113–118. [Google Scholar] [CrossRef]
- Blaze, J. A Comparison of Current Regulatory Frameworks for Nutraceuticals in Australia, Canada, Japan, and the United States. Innov. Pharm. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Grgić, J.; Šelo, G.; Planinić, M.; Tišma, M.; Bucić-Kojić, A. Role of the Encapsulation in Bioavailability of Phenolic Compounds. Antioxidants 2020, 9, 923. [Google Scholar] [CrossRef] [PubMed]
- Tišma, M.; Žnidaršič-Plazl, P.; Šelo, G.; Tolj, I.; Šperanda, M.; Bucić-Kojić, A.; Planinić, M. Trametes versicolor in lignocellulose-based bioeconomy: State of the art, challenges and opportunities. Bioresour. Technol. 2021, 330, 124997. [Google Scholar] [CrossRef] [PubMed]
- Dulf, F.V.; Vodnar, D.C.; Dulf, E.H.; Pintea, A. Phenolic compounds, flavonoids, lipids and antioxidant potential of apricot (Prunusarmeniaca L.) pomace fermented by two filamentous fungal strains in solid state system. Chem. Cent. J. 2017, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Leite, P.; Silva, C.; Salgado, J.M.; Belo, I. Simultaneous production of lignocellulolytic enzymes and extraction of antioxidant compounds by solid-state fermentation of agro-industrial wastes. Ind. Crops Prod. 2019, 137, 315–322. [Google Scholar] [CrossRef]
- Bianco, A.; Budroni, M.; Zara, S.; Mannazzu, I.; Fancello, F.; Zara, G. The role of microorganisms on biotransformation of brewers’ spent grain. Appl. Microbiol. Biotechnol. 2020, 104, 8661–8678. [Google Scholar] [CrossRef] [PubMed]
- Chetrariu, A.; Dabija, A. Brewer’s Spent Grains: Possibilities of Valorization, a Review. Appl. Sci. 2020, 10, 5619. [Google Scholar] [CrossRef]
- Mitri, S.; Salameh, S.J.; Khelfa, A.; Leonard, E.; Maroun, R.G.; Louka, N.; Koubaa, M. Valorization of Brewers’ Spent Grains: Pretreatments and Fermentation, a Review. Fermentation 2022, 8, 50. [Google Scholar] [CrossRef]
- Fernandes, H.; Moyano, F.; Castro, C.; Salgado, J.; Martinez, F.; Aznar, M.; Fernandes, N.; Ferreira, P.; Gonçalves, M.; Belo, I.; et al. Solid-state fermented brewer’s spent grain enzymatic extract increases in vitro and in vivo feed digestibility in European seabass. Sci. Rep. 2021, 11, 22946. [Google Scholar] [CrossRef]
- Cooray, S.T.; Chen, W.N. Valorization of brewer’s spent grain using fungi solid-state fermentation to enhance nutritional value. J. Funct. Foods 2018, 42, 85–94. [Google Scholar] [CrossRef]
- Jacobsen, R.M.; Sverdrup-Thygeson, A.; Kauserud, H.; Mundra, S.; Birkemoe, T. Exclusion of invertebrates influences saprotrophic fungal community and wood decay rate in an experimental field study. Funct. Ecol. 2018, 32, 2571–2582. [Google Scholar] [CrossRef]
- Webster, J.; Weber, R. Introduction to Fungi, 3rd ed.; Cambridge University Press: Cambridge, UK, 2007; p. 867. [Google Scholar]
- Index Fungorum. Available online: http://www.indexfungorum.org/ (accessed on 5 November 2022).
- Macalester. Turkey Tail Fungus. Available online: https://www.macalester.edu/ordway/biodiversity/inventory/turkeytailfungus/ (accessed on 7 November 2022).
- Tišma, M.; Jurić, A.; Bucić-Kojić, A.; Panjičko, M.; Planinić, M. Biovalorization of brewers’ spent grain for the production of laccase and polyphenols. J. Inst. Brew. 2018, 124, 182–186. [Google Scholar] [CrossRef]
- Bailey, M.J.; Biely, P.; Poutanen, K. Interlaboratory testing of methods for assay of xylanase activity. J. Biotechnol. 1992, 23, 257–270. [Google Scholar] [CrossRef]
- Adney, B.; Baker, J. Measurement of Cellulase Activities. Laboratory Analytical Procedure; National Renewable Energy Laboratory (NREL): Golden, CO, USA, 1962.
- Karpe, A.V.; Dhamale, V.V.; Morrison, P.D.; Beale, D.J.; Harding, I.H.; Palombo, E.A. Winery biomass waste degradation by sequential sonication and mixed fungal enzyme treatments. Fungal Genet. Biol. 2016, 102, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Lueangjaroenkit, P.L.; Kunitake, E.; Sakkam, M.; Kimura, T.; Teerapatsakul, C.; Sakka, K.; Chitradon, L. Light regulation of two new manganese peroxidase-encoding genes in Trametesolyzona KU-RNW027. Microorganisms 2020, 8, 852. [Google Scholar] [CrossRef] [PubMed]
- Tišma, M.; Žnidaršič-Plazl, P.; Vasić-Rački, Đ.; Zelić, B. Optimization of laccase production by Trametes versicolor cultivated on industrial waste. Appl. Biochem. Biotechnol. 2012, 166, 36–46. [Google Scholar] [CrossRef]
- Tien, M.; Kirk, T.K. Lignin peroxidase of Phanerochaete chrysosporium. Biomass Part B Lignin Pectin Chitin 1988, 161, 238–249. [Google Scholar] [CrossRef]
- Šelo, G.; Planinić, M.; Tišma, M.; Grgić, J.; Perković, G.; Koceva Komlenić, D.; Bucić-Kojić, A. A Comparative Study of the Influence of Various Fungal-Based Pretreatments of Grape Pomace on Phenolic Compounds Recovery. Foods 2022, 11, 1665. [Google Scholar] [CrossRef] [PubMed]
- AOAC. AOAC Official Method 2001.11. In Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- AOAC. OAC Official Method 945.16. In Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Goering, H.K.; Van Soest, P.J. Forage Fiber Analyses (Apparatus, Reagents, Procedures, and Some Applications); Agricultural Research Service, U.S. Department of Agriculture: Washington, DC, USA, 1970; pp. 1–24.
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Marinova, D.; Ribarova, F.; Atanassova, M. Total phenolics and total flavonoids in Bulgarian fruits and vegetables. J. Chem. Technol. Metall. 2005, 40, 255–260. [Google Scholar]
- Planinić, M.; Zelić, B.; Čubel, I.; Bucić-Kojić, A.; Tišma, M. Corn forage biological pretreatment by Trametes Versicolor in a tray bioreactor. Waste Manag. Res. 2016, 34, 802–809. [Google Scholar] [CrossRef]
- Škerget, M.; Kotnik, P.; Hadolin, M.; Hraš, A.R.; Simonič, M.; Knez, Ž. Phenols, proanthocyanidins, flavones and flavonols in some plant materials and their antioxidant activities. Food Chem. 2005, 89, 191–198. [Google Scholar] [CrossRef]
- He, Q.; Peng, H.; Sheng, M.; Hu, S.; Qiu, J.; Gu, J. Humidity Control Strategies for Solid-State Fermentation: Capillary Water Supply by Water-Retention Materials and Negative-Pressure Auto-controlled Irrigation. Front. Bioeng. Biotechnol. 2019, 7, 263. [Google Scholar] [CrossRef] [PubMed]
- Bucić-Kojić, A.; Šelo, G.; Zelić, B.; Planinić, M.; Tišma, M. Recovery of Phenolic Acid and Enzyme Production from Corn Silage Biologically Treated by Trametes versicolor. Appl. Biochem. Biotechnol. 2016, 181, 948–960. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, H.M.N.; Asgher, M.; Bhatti, H.N. Optimization of physical and nutritional factors for synthesis of lignin degrading enzymes by a novel strain of Trametes versicolor. Bioresources 2011, 6, 1273–1278. [Google Scholar]
- Gassara, F.; Brar, S.K.; Tyagi, R.D.; Verma, M.; Surampalli, R.Y. Screening of agro-industrial wastes to produce ligninolytic enzymes by Phanerochaete chrysosporium. Biochem. Eng. J. 2010, 49, 388–394. [Google Scholar] [CrossRef]
- Terrasan, C.R.F.; Carmona, E.C. Solid-state fermentation of brewer’s spent grain for xylanolytic enzymes production by Penicillium janczewskii and analyses of the fermented substrate. Biosci. J. 2015, 31, 1826–1836. [Google Scholar] [CrossRef]
- Moran-Aguilar, M.G.; Costa-Trigo, I.; Calderón-Santoyo, M.; Domínguez, J.M.; Aguilar-Uscanga, M.G. Production of cellulases and xylanases in solid-state fermentation by different strains of Aspergillus niger using sugarcane bagasse and brewery spent grain. Biochem. Eng. J. 2021, 172, 108060. [Google Scholar] [CrossRef]
- Hassan, S.S.; Tiwari, B.K.; Williams, G.A.; Jaiswal, A.K. Bioprocessing of brewers’ spent grain for production of xylanopectinolytic enzymes by Mucor sp. Bioresour. Technol. Rep. 2019, 9, 100371. [Google Scholar] [CrossRef]
- Outeiriño, D.; Costa-Trigo, I.; de Souza Oliveira, R.P.; Pérez Guerra, N.; Domínguez, J.M. A novel approach to the biorefinery of brewery spent grain. Process Biochem. 2019, 85, 135–142. [Google Scholar] [CrossRef]
- Ilić, N.; Davidović, S.; Milić, M.; Rajilić-Stojanović, M.; Pecarski, D.; Ivančić-Šantek, M.; Mihajlovski, K.; Dimitrijević-Branković, S. The agricultural waste utilization for exploration of the extracellular enzyme potential of three novel white-rot Basidiomycetes: Fomesfomentarius TMF2, Schizophyllum commune TMF3 and Bjerkandera adusta TMF1. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Liguori, R.; Soccol, R.C.; Vandenberghe, P.L.; Woiciechowski, L.A.; Ionata, E.; Marcolongo, L.; Faraco, V. Selection of the strain Lactobacillus acidophilus ATCC 43121 and its application to brewers’ spent grain conversion into lactic acid. BioMed Res. Int. 2015, 2015, 240231. [Google Scholar] [CrossRef] [PubMed]
- Ibarruri, J.; Cebrián, M.; Hernández, I. Solid State Fermentation of Brewer’s Spent Grain Using Rhizopus sp. to Enhance Nutritional Value. Waste Biomass Valorization 2019, 10, 3687–3700. [Google Scholar] [CrossRef]
- Aftab, M.N.; Iqbal, I.; Riaz, F.; Karadag, A.; Tabatabaei, M. Different Pretreatment Methods of Lignocellulosic Biomass for Use in Biofuel Production. In Biomass for Bioenergy—Recent Trends and Future Challenges; Abomohra, A.E., Ed.; IntechOpen: London, UK, 2019; pp. 1–24. [Google Scholar] [CrossRef]
- Llimós, J.; Martínez-Avila, O.; Marti, E.; Corchado-Lopo, C.; Llenas, L.; Gea, T.; Ponsá, S. Brewer’s spent grain biotransformation to produce lignocellulolytic enzymes and polyhydroxyalkanoates in a two-stage valorization scheme. Biomass Convers. Biorefinery 2020, 12, 3921–3932. [Google Scholar] [CrossRef]
- Da Costa Maia, I.; Thomaz dos Santos D’Almeida, C.; Guimarães Freire, D.M.; Cavalcanti, E.; d’Avila, C.; Cameron, L.C.; Dias, J.F.; Larraz Ferreira, M.S. Effect of solid-state fermentation over the release of phenolic compounds from brewer’s spent grain revealed by UPLC-MSE. LWT—Food Sci. Technol. 2020, 133, 110136. [Google Scholar] [CrossRef]
- Birsan, R.I.; Wilde, P.; Waldron, K.W.; Rai, D.K. Recovery of polyphenols from brewer’s spent grains. Antioxidants 2019, 8, 380. [Google Scholar] [CrossRef] [PubMed]
- Macias-Garbett, R.; Serna-Hernández, S.O.; Sosa-Hernández, J.E.; Parra-Saldívar, R. Phenolic compounds from brewer’s spent grains: Toward green recovery methods and applications in the cosmetic industry. Front. Sustain. Food Syst. 2021, 5, 681684. [Google Scholar] [CrossRef]
- Su, Y.; Wenzel, M.; Paasch, S.; Seifert, M.; Böhm, W.; Doert, T.; Weigand, J.J. Recycling of Brewer’s Spent Grain as a Biosorbent by Nitro-Oxidation for Uranyl Ion Removal from Wastewater. ACS Omega 2021, 6, 19364–19377. [Google Scholar] [CrossRef]
- El Hariri El Nokab, M.; Habib, M.H.; Alassmy, Y.A.; Abduljawad, M.M.; Alshamrani, K.M.; Sebakhy, K.O. Solid State NMR a Powerful Technique for Investigating Sustainable/Renewable Cellulose-Based Materials. Polymers 2022, 14, 1049. [Google Scholar] [CrossRef]
- Sparrman, T.; Svenningsson, L.; Sahlin-Sjövold, K.; Nordstierna, L.; Westman, G.; Bernin, D. A revised solid-state NMR method to assess the crystallinity of cellulose. Cellulose 2019, 26, 8993–9003. [Google Scholar] [CrossRef]
- Cipriano, D.F.; Chinelatto, L.S.; Nascimento, S.A.; Rezende, C.A.; de Menezes, S.M.C.; Freitas, J.C.C. Potential and limitations of 13C CP/MAS NMR spectroscopy to determine the lignin content of lignocellulosic feedstock. Biomass Bioenergy 2020, 142, 105792. [Google Scholar] [CrossRef]
- Allegretti, C.; Bellinetto, E.; D’Arrigo, P.; Griffini, G.; Marzorati, S.; Rossato, L.A.M.; Ruffini, E.; Schiavi, L.; Serra, S.; Strini, A.; et al. Towards a Complete Exploitation of Brewers’ Spent Grain from a Circular Economy Perspective. Fermentation 2022, 8, 151. [Google Scholar] [CrossRef]
- Mastanjević, K.; Šarkanj, B.; Warth, B.; Krska, R.; Sulyok, M.; Mastanjević, K.; Šantek, B.; Krstanović, V. Fusarium culmorum multi-toxin screening in malting and brewing by-products. LWT—Food Sci. Technol. 2018, 98, 642–645. [Google Scholar] [CrossRef]
- Lao, E.J. The implications and management strategies of animal feed mycotoxins. Rev. Environ. Contam. Toxicol. 2020, 3, 53–61. [Google Scholar] [CrossRef]
- Santos Pereira, C.C.; Cunha, S.; Fernandes, J.O. Prevalent Mycotoxins in Animal Feed: Occurrence and Analytical Methods. Toxins 2019, 11, 290. [Google Scholar] [CrossRef]
- Babič, J.; Tavčar-Kalcher, G.; Celar, F.A.; Kos, K.; Červek, M.; Jakovac-Strajn, B. Ergot and ergot alkaloids in cereal grains intended for animal feeding collected in Slovenia: Occurrence, pattern and correlations. Toxins 2020, 12, 730. [Google Scholar] [CrossRef]
- Romera-Torres, A.; Romero-González, R.; Martínez Vidal, J.L.; Frenich, A.G. Study of the occurrence of tropane alkaloids in animal feed using LC-HRMS. Anal. Methods 2018, 10, 3340–3346. [Google Scholar] [CrossRef]
- Toman, R.; Pšenková, M.; Tančin, V. Polychlorinated biphenyls in cow’s milk, feed and soil in selected areas of Slovakia. Acta Fytotech. Zootech. 2020, 23, 4. [Google Scholar] [CrossRef]
- Şahin, T.; Dalğa, S.; Ölmez, M. Polycyclic Aromatic Hydrocarbons (PAHs) and Their Importance in Animal Nutrition. In Animal Husbandry; Kukovics, S., Ed.; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Hejna, M.; Gottardo, D.; Baldi, A.; Dell’Orto, V.; Cheli, F.; Zaninelli, M.; Rossi, L. Nutritional ecology of heavy metals. Animal 2018, 12, 2156–2170. [Google Scholar] [CrossRef]
- Eur-LEX. Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on Undesirable Substances in Animal Feed. Available online: http://data.europa.eu/eli/dir/2002/32/2019-11-28 (accessed on 20 September 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component [%] | Raw BSG1 | Fermented BSG1 | Raw BSG2 | Fermented BSG2 |
|---|---|---|---|---|
| Cellulose | 21.47 ± 1.22 | 17.82 ± 1.57 | 28.34 ± 0.46 | 26.59 ± 0.75 |
| Hemicellulose | 30.95 ± 1.55 | 27.87 ± 0.87 | 14.89 ± 1.25 | 20.26 ± 2.39 |
| Lignin | 6.94 ± 0.95 | 5.75 ± 0.34 | 3.81 ± 0.26 | 6.90 ± 1.97 |
| Crude Proteins | 23.07 ± 0.13 | 26.42 ± 0.11 | 18.34 ± 0.01 | 25.29 ± 0.24 |
| Lipids | 8.09 ± 0.17 | 3.82 ± 0.19 | 8.85 ± 0.62 | 3.99 ± 0.55 |
| Ash | 3.86 ± 0.03 | 5.00 ± 0.00 | 4.24 ± 0.06 | 5.73 ± 0.11 |
| cmax [mg/gDM] * | Raw BSG1 | Fermented BSG1 | SSF Day | Raw BSG2 | Fermented BSG2 | SSF Day |
|---|---|---|---|---|---|---|
| Total Polyphenols | 4.495 ± 0.270 | 16.867 ± 0.126 | 14 | 9.242 ± 0.068 | 15.198 ± 0.384 | 7 |
| Total Flavonoids | 1.268 ± 0.025 | 2.056 ± 0.029 | 11 | 1.971 ± 0.044 | 2.246 ± 0.139 | 7 |
| Total Proanthocyanidins | 0.517 ± 0.431 | 0.287 ± 0.030 | 15 | 0.494 ± 0.009 | 0.456 ± 0.009 | 15 |
| Individual Phenols | ||||||
| p-hydroxybenzoic acid | 0.022 ± 0.001 | 0.105 ± 0.003 | 3 | 0.203 ± 0.047 | 0.324 ± 0.008 | 6 |
| Catechin | 0.148 ± 0.009 | 1.301 ± 0.025 | 15 | 0.188 ± 0.011 | 1.152 ± 0.047 | 7 |
| Epicatechin | 0.092 ± 0.005 | 0.137 ± 0.010 | 11 | 0.035 ± 0.000 | 0.149 ± 0.001 | 3 |
| Vanillic acid | 0.002 ± 0.00 | 0.013 ± 0.000 | 15 | n.d | n.d. | - |
| Syringic acid | 0.031 ± 0.001 | 0.281 ± 0.000 | 9 | 0.015 ± 0.000 | 0.218 ± 0.000 | 7 |
| p-coumaric acid | 0.008 ± 0.000 | 0.019 ± 0.000 | 12 | 0.004 ± 0.000 | 0.024 ± 0.001 | 5 |
| Ferulic acid | n.d. | 0.139 ± 0.000 | 12 | 0.166 ± 0.027 | 0.164 ± 0.006 | 5 |
| Kaempferol | 0.023 ± 0.000 | 0.0230 ± 0.003 | 10 | 0.016 ± 0.000 | 0.040 ± 0.002 | 7 |
| 3,4-dihydroxybenzoic acid | 0.462 ± 0.001 | 6.823 ± 2.159 | 3 | 0.025 ± 0.000 | 0.141 ± 0.008 | 12 |
| o-coumaric acid | 0.012 ± 0.001 | 0.019 ± 0.001 | 8 | 0.012 ± 0.001 | 0.019 ± 0.001 | 8 |
| p-hydroxyphenylacetic acid | n.d. | n.d. | - | 0.203 ± 0.047 | 0.324 ± 0.008 | 6 |
| /ppm | H-Atom | Raw BSG1/% * | Fermented BSG1/% * | Raw BSG2/% | Fermented BSG2/% |
|---|---|---|---|---|---|
| 0.50–3.00 | Aliphatic | 29.17 | 31.33 | 25.71 | 30.82 |
| 3.00–4.40 | Carbohydrate, methoxy | 69.45 | 66.77 | 71.42 | 62.89 |
| 6.70–7.65 | Aromatic | 0.70 | 1.20 | 2.14 | 5.66 |
| 7.94–8.50 | Aromatic, amino acid, formic acid | 0.70 | 0.70 | 0.73 | 0.63 |
| Compound | Raw BSG1 | Fermented BSG1 | Raw BSG2 | Fermented BSG2 |
|---|---|---|---|---|
| Phosphorus [%] | 0.48 | 0.60 | 0.66 | 0.80 |
| Calcium [mg/kg] | 2073 | 2456 | 2763 | 2994 |
| Potassium [mg/kg] | 586 | 499 | 722 | 936 |
| Iron [mg/kg] | 173 | 220 | 103 | 113 |
| Lead [mg/kg] | 0.049 | 0.064 | 0.025 | 0.033 |
| Cadmium [mg/kg] | 0.039 | 0.048 | 0.044 | 0.057 |
| Arsenic [mg/kg] | 0.032 | 0.039 | <0.010 | 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeko-Pivač, A.; Bošnjaković, A.; Planinić, M.; Parlov Vuković, J.; Novak, P.; Jednačak, T.; Tišma, M. Improvement of the Nutraceutical Profile of Brewer’s Spent Grain after Treatment with Trametes versicolor. Microorganisms 2022, 10, 2295. https://doi.org/10.3390/microorganisms10112295
Zeko-Pivač A, Bošnjaković A, Planinić M, Parlov Vuković J, Novak P, Jednačak T, Tišma M. Improvement of the Nutraceutical Profile of Brewer’s Spent Grain after Treatment with Trametes versicolor. Microorganisms. 2022; 10(11):2295. https://doi.org/10.3390/microorganisms10112295
Chicago/Turabian StyleZeko-Pivač, Anđela, Anja Bošnjaković, Mirela Planinić, Jelena Parlov Vuković, Predrag Novak, Tomislav Jednačak, and Marina Tišma. 2022. "Improvement of the Nutraceutical Profile of Brewer’s Spent Grain after Treatment with Trametes versicolor" Microorganisms 10, no. 11: 2295. https://doi.org/10.3390/microorganisms10112295
APA StyleZeko-Pivač, A., Bošnjaković, A., Planinić, M., Parlov Vuković, J., Novak, P., Jednačak, T., & Tišma, M. (2022). Improvement of the Nutraceutical Profile of Brewer’s Spent Grain after Treatment with Trametes versicolor. Microorganisms, 10(11), 2295. https://doi.org/10.3390/microorganisms10112295