
Microbial Communities of Artisanal Fermented Milk Products from Russia
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
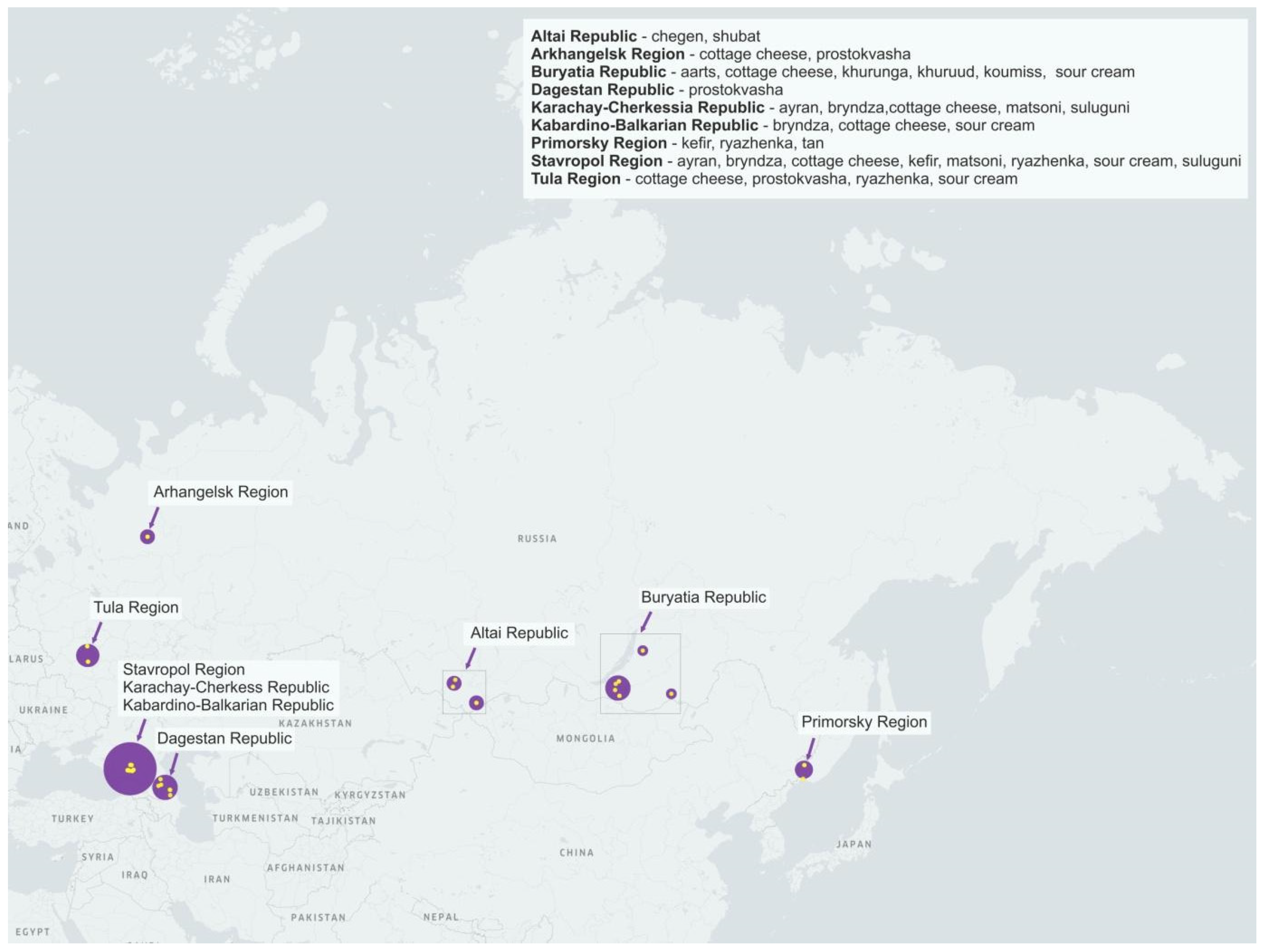
2.1. Collection of Dairy Product Samples
2.2. Volatile Fatty Acids Analysis
2.3. DNA Extraction and Amplicon Sequencing
2.4. Bioinformatics and Statistical Analysis
3. Results and Discussion
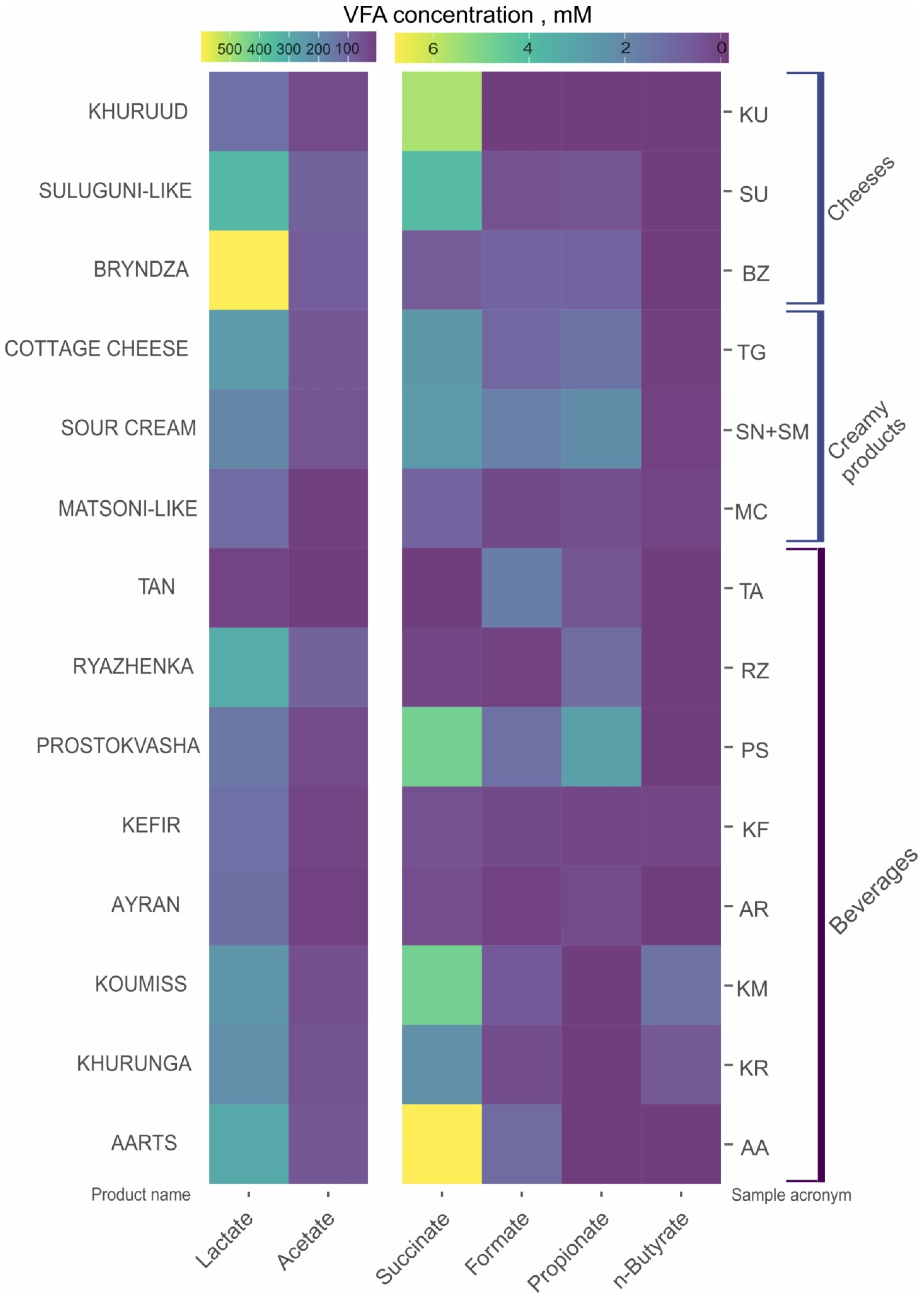
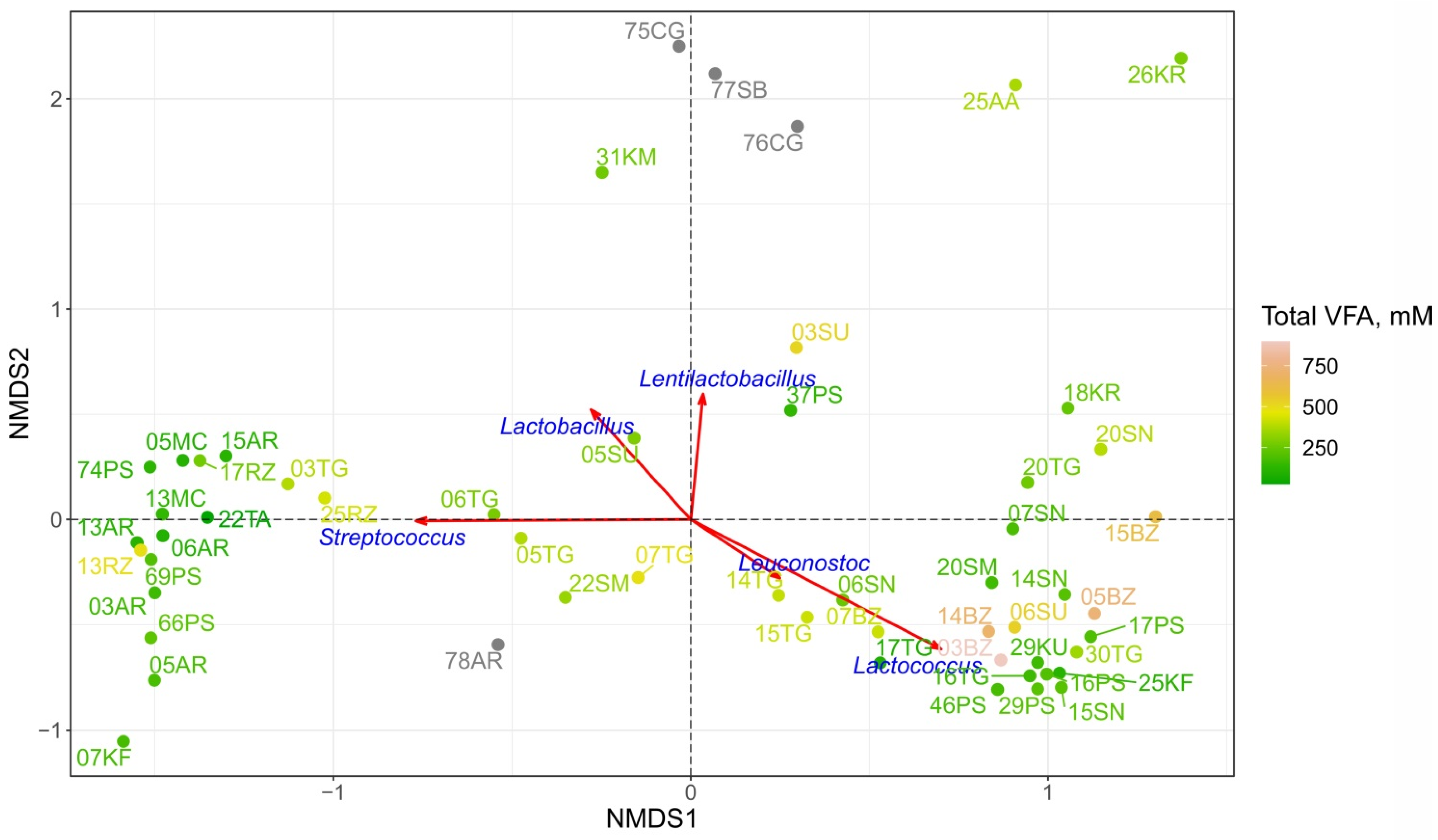
3.1. Volatile Fatty Acids Content
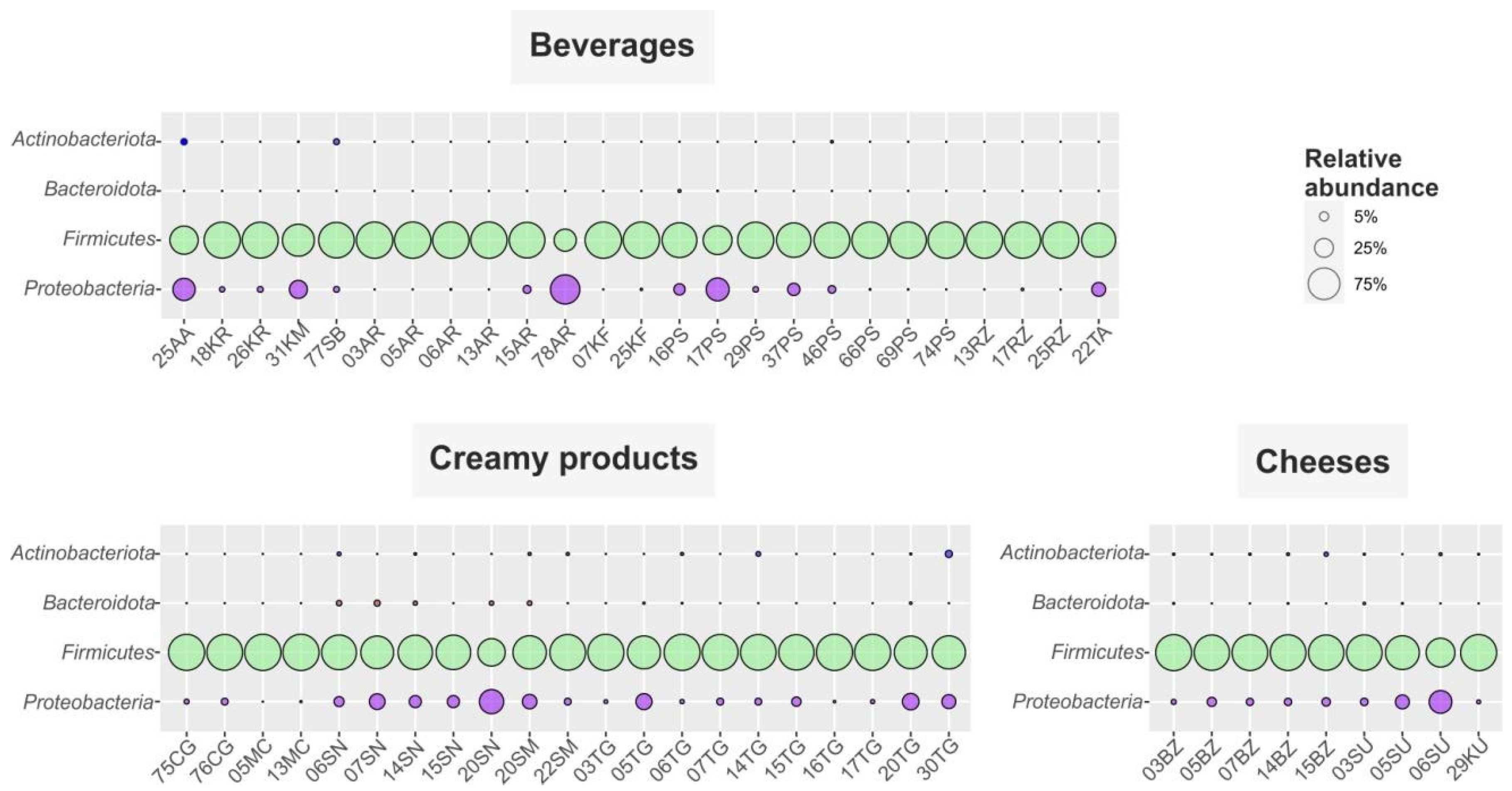
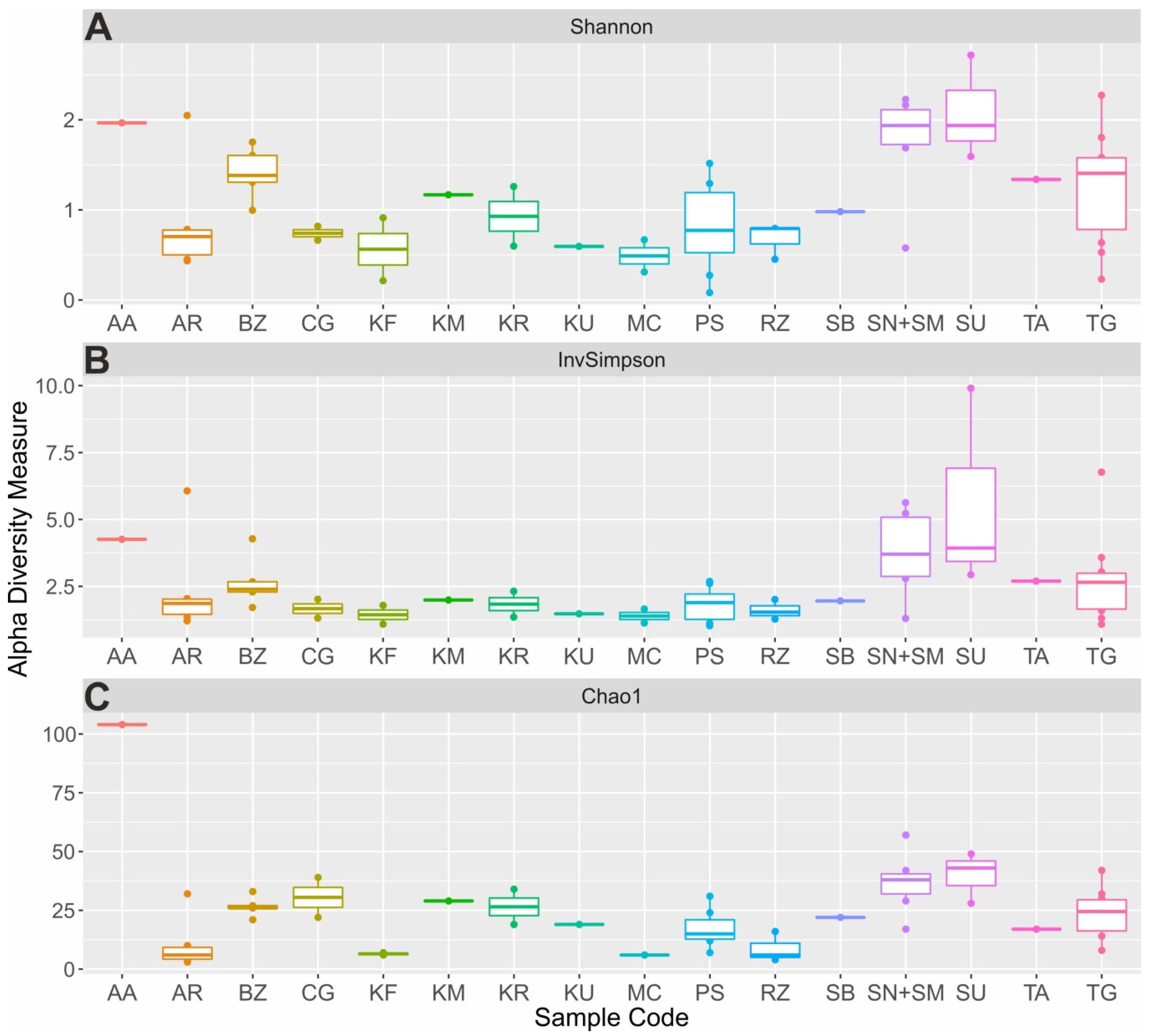
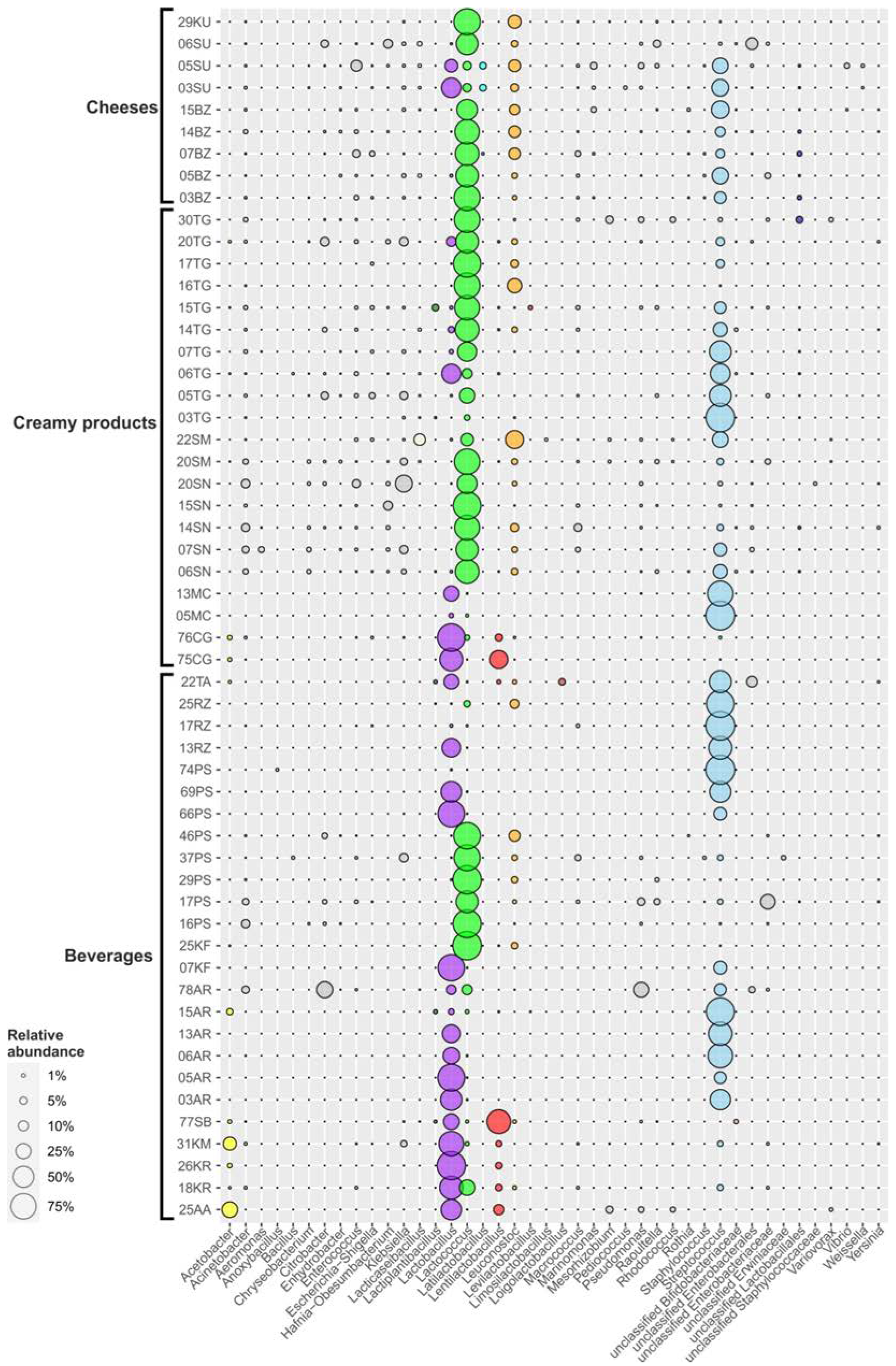
3.2. Biodiversity and Taxonomic Profiling of the Studied Fermented Milk Products
3.2.1. Fermented Milk Beverages
3.2.2. Creamy Fermented Milk Products
3.2.3. Cheeses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kandasamy, S.; Kavitake, D.; Shetty, P.H. Lactic acid bacteria and yeasts as starter cultures for fermented foods and their role in commercialization of fermented foods. In Innovations in Technologies for Fermented Food and Beverage Industries; Springer International Publishing: Cham, Switzerland, 2018; pp. 25–52. [Google Scholar]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Review: Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed]
- Pokhlebkin, V.V. The Ethnic Cuisines of Our Peoples; Centrpoligraph: Moscow, Russia, 2004; p. 639. [Google Scholar]
- Teuber, M. The genus Lactococcus. In The Genera of Lactic Acid Bacteria; Springer US: Boston, MA, USA, 1995; pp. 173–234. [Google Scholar]
- de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Maske, B.L.; De Dea Lindner, J.; Vale, A.S.; Favero, G.R.; Viesser, J.; de Carvalho, J.C.; Góes-Neto, A.; Soccol, C.R. An updated review on bacterial community composition of traditional fermented milk products: What next-generation sequencing has revealed so far? Crit. Rev. Food Sci. Nutr. 2022, 62, 1870–1889. [Google Scholar] [CrossRef] [PubMed]
- Mayo, B.; Rachid, C.; Alegria, A.; Leite, A.; Peixoto, R.; Delgado, S. Impact of next generation sequencing techniques in food microbiology. Curr. Genom. 2014, 15, 293–309. [Google Scholar] [CrossRef] [PubMed]
- Kamilari, E.; Tomazou, M.; Antoniades, A.; Tsaltas, D. High throughput sequencing technologies as a new toolbox for deep analysis, characterization and potentially authentication of protection designation of origin cheeses? Int. J. Food Sci. 2019, 2019, 5837301. [Google Scholar] [CrossRef]
- Marsh, A.J.; O’Sullivan, O.; Hill, C.; Ross, R.P.; Cotter, P.D. Sequencing-based analysis of the bacterial and fungal composition of kefir grains and milks from multiple sources. PLoS ONE 2013, 8, e69371. [Google Scholar] [CrossRef]
- Oki, K.; Dugersuren, J.; Demberel, S.; Watanabe, K. Pyrosequencing analysis of the microbial diversity of airag, khoormog and tarag, traditional fermented dairy products of Mongolia. Biosci. Microbiota Food Health 2014, 33, 53–64. [Google Scholar] [CrossRef]
- Garofalo, C.; Osimani, A.; Milanović, V.; Aquilanti, L.; De Filippis, F.; Stellato, G.; Di Mauro, S.; Turchetti, B.; Buzzini, P.; Ercolini, D.; et al. Bacteria and yeast microbiota in milk kefir grains from different Italian regions. Food Microbiol. 2015, 49, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Shangpliang, H.N.J.; Rai, R.; Keisam, S.; Jeyaram, K.; Tamang, J.P. Bacterial community in naturally fermented milk products of Arunachal Pradesh and Sikkim of India analyzed by high-throughput amplicon sequencing. Sci. Rep. 2018, 8, 1532. [Google Scholar] [CrossRef]
- Martin, N.H.; Trmčić, A.; Hsieh, T.-H.; Boor, K.J.; Wiedmann, M. The evolving role of coliforms as indicators of unhygienic processing conditions in dairy foods. Front. Microbiol. 2016, 7, 1549. [Google Scholar] [CrossRef]
- Dertli, E.; Çon, A.H. Microbial diversity of traditional kefir grains and their role on kefir aroma. LWT-Food Sci. Technol. 2017, 85, 151–157. [Google Scholar] [CrossRef]
- Issa, A.T.; Tahergorabi, R. Milk bacteria and gastrointestinal tract. In Dietary Interventions in Gastrointestinal Diseases; Elsevier: Amsterdam, The Netherlands, 2019; pp. 265–275. [Google Scholar]
- Shiby, V.K.; Mishra, H.N. Fermented milks and milk products as functional foods—A review. Crit. Rev. Food Sci. Nutr. 2013, 53, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Leech, J.; Cabrera-Rubio, R.; Walsh, A.M.; Macori, G.; Walsh, C.J.; Barton, W.; Finnegan, L.; Crispie, F.; O’Sullivan, O.; Claesson, M.J.; et al. Fermented-food metagenomics reveals substrate-associated differences in taxonomy and health-associated and antibiotic resistance determinants. Msystems 2020, 5, e00522-20. [Google Scholar] [CrossRef] [PubMed]
- Terzić-Vidojević, A.; Veljović, K.; Tolinački, M.; Živković, M.; Lukić, J.; Lozo, J.; Fira, Đ.; Jovčić, B.; Strahinić, I.; Begović, J.; et al. Diversity of non-starter lactic acid bacteria in autochthonous dairy products from Western Balkan Countries-Technological and probiotic properties. Food Res. Int. 2020, 136, 109494. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhao, F.; Hou, Q.; Wang, J.; Li, M.; Sun, Z. PacBio sequencing reveals bacterial community diversity in cheeses collected from different regions. J. Dairy Sci. 2020, 103, 1238–1249. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, Y.; Kwok, L.-Y.; Sun, Z.; Zhang, J.; Guo, Z.; Hou, Q.; Menhe, B.; Zhang, H. High-throughput sequencing for the detection of the bacterial and fungal diversity in Mongolian naturally fermented cow’s milk in Russia. BMC Microbiol. 2015, 15, 45. [Google Scholar] [CrossRef]
- Jin, H.; Mo, L.; Pan, L.; Hou, Q.; Li, C.; Darima, I.; Yu, J. Using PacBio sequencing to investigate the bacterial microbiota of traditional Buryatian cottage cheese and comparison with Italian and Kazakhstan artisanal cheeses. J. Dairy Sci. 2018, 101, 6885–6896. [Google Scholar] [CrossRef]
- Yu, Z.; Peng, C.; Kwok, L.; Zhang, H. The bacterial diversity of spontaneously fermented dairy products collected in Northeast Asia. Foods 2021, 10, 2321. [Google Scholar] [CrossRef]
- Gohl, D.M.; Vangay, P.; Garbe, J.; MacLean, A.; Hauge, A.; Becker, A.; Gould, T.J.; Clayton, J.B.; Johnson, T.J.; Hunter, R.; et al. Systematic improvement of amplicon marker gene methods for increased accuracy in microbiome studies. Nat. Biotechnol. 2016, 34, 942–949. [Google Scholar] [CrossRef]
- Hugerth, L.W.; Wefer, H.A.; Lundin, S.; Jakobsson, H.E.; Lindberg, M.; Rodin, S.; Engstrand, L.; Andersson, A.F. DegePrime, a program for degenerate primer design for broad-taxonomic-range PCR in microbial ecology studies. Appl. Environ. Microbiol. 2014, 80, 5116–5123. [Google Scholar] [CrossRef]
- Merkel, A.Y.; Tarnovetskii, I.Y.; Podosokorskaya, O.A.; Toshchakov, S.V. Analysis of 16S rRNA Primer systems for profiling of thermophilic microbial communities. Microbiology 2019, 88, 671–680. [Google Scholar] [CrossRef]
- Vortsepneva, E.; Chevaldonné, P.; Klyukina, A.; Naduvaeva, E.; Todt, C.; Zhadan, A.; Tzetlin, A.; Kublanov, I. Microbial associations of shallow-water Mediterranean marine cave Solenogastres (Mollusca). PeerJ 2021, 9, e12655. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Wagner, H. R Package, Version 2. In Vegan: Community Ecology Package; 2019; p. 5. Available online: https://github.com/vegandevs/vegan (accessed on 19 September 2022).
- Mather, I.H. A review and proposed nomenclature for major proteins of the milk-fat globule membrane. J. Dairy Sci. 2000, 83, 203–247. [Google Scholar] [CrossRef]
- Watanabe, K.; Fujimoto, J.; Sasamoto, M.; Dugersuren, J.; Tumursuh, T.; Demberel, S. Diversity of lactic acid bacteria and yeasts in Airag and Tarag, traditional fermented milk products of Mongolia. World J. Microbiol. Biotechnol. 2008, 24, 1313–1325. [Google Scholar] [CrossRef]
- Kozhakhmetov, S.; Tynybayeva, I.; Baikhanova, D.; Saduakhasova, S.; Shakhabayeva, G.; Kushugulova, A.; Nurgozhin, T.; Zhumadilov, Z. Metagenomic analysis of koumiss in Kazakhstan. Cent. Asian J. Glob. Health 2014, 3, 163. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Yang, C.; Jin, H.; Kwok, L.-Y.; Sun, Z.; Zhang, H. Metagenomic features of traditional fermented milk products. LWT 2022, 155, 112945. [Google Scholar] [CrossRef]
- Watanabe, K.; Makino, H.; Sasamoto, M.; Kudo, Y.; Fujimoto, J.; Demberel, S. Bifidobacterium mongoliense sp. nov., from airag, a traditional fermented mare’s milk product from Mongolia. Int. J. Syst. Evol. Microbiol. 2009, 59, 1535–1540. [Google Scholar] [CrossRef]
- Soleymanzadeh, N.; Mirdamadi, S.; Kianirad, M. Antioxidant activity of camel and bovine milk fermented by lactic acid bacteria isolated from traditional fermented camel milk (Chal). Dairy Sci. Technol. 2016, 96, 443–457. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Ibacache-Quiroga, C.; González-Pizarro, K.; Charifeh, M.; Canales, C.; Díaz-Viciedo, R.; Schmachtenberg, O.; Dinamarca, M.A. Metagenomic and functional characterization of two Chilean kefir beverages reveals a dairy beverage containing active enzymes, short-chain fatty acids, microbial β-amyloids, and bio-film inhibitors. Foods 2022, 11, 900. [Google Scholar] [CrossRef]
- Walsh, A.M.; Crispie, F.; Kilcawley, K.; O’Sullivan, O.; O’Sullivan, M.G.; Claesson, M.J.; Cotter, P.D. Microbial succession and flavor production in the fermented dairy beverage kefir. Msystems 2016, 1, e00052-16. [Google Scholar] [CrossRef]
- He, G.; Liu, T.; Sadiq, F.A.; Gu, J.; Zhang, G. Insights into the microbial diversity and community dynamics of Chinese traditional fermented foods from using high-throughput sequencing approaches. J. Zhejiang Univ.-Sci. B 2017, 18, 289–302. [Google Scholar] [CrossRef]
- Dobson, A.; O’Sullivan, O.; Cotter, P.D.; Ross, P.; Hill, C. High-throughput sequence-based analysis of the bacterial composition of kefir and an associated kefir grain. FEMS Microbiol. Lett. 2011, 320, 56–62. [Google Scholar] [CrossRef]
- Korsak, N.; Taminiau, B.; Leclercq, M.; Nezer, C.; Crevecoeur, S.; Ferauche, C.; Detry, E.; Delcenserie, V.; Daube, G. Short communication: Evaluation of the microbiota of kefir samples using metagenetic analysis targeting the 16S and 26S ribosomal DNA fragments. J. Dairy Sci. 2015, 98, 3684–3689. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wang, H.M.; Zha, M.S.; Qing, Y.T.; Bai, N.; Ren, Y.; Xi, X.X.; Liu, W.J.; Menghe, B.L.G.; Zhang, H.P. Molecular identification and quantification of lactic acid bacteria in traditional fermented dairy foods of Russia. J. Dairy Sci. 2015, 98, 5143–5154. [Google Scholar] [CrossRef] [PubMed]
- Moiseenko, K.V.; Glazunova, O.A.; Savinova, O.S.; Ajibade, B.O.; Ijabadeniyi, O.A.; Fedorova, T.V. Analytical characterization of the widely consumed commercialized fermented beverages from Russia (kefir and ryazhenka) and South Africa (amasi and mahewu): Potential functional properties and profiles of volatile organic compounds. Foods 2021, 10, 3082. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Urashima, T.; Chanishvili, N.; Arai, I.; Motoshima, H. Major microbiota of lactic acid bacteria from Matsoni, a traditional Georgian fermented milk. Anim. Sci. J. 2007, 78, 85–91. [Google Scholar] [CrossRef]
- Yu, J.; Mo, L.; Pan, L.; Yao, C.; Ren, D.; An, X.; Tsogtgerel, T.; Zhang, H.; Liu, W. Bacterial microbiota and metabolic character of traditional sour cream and butter in Buryatia, Russia. Front. Microbiol. 2018, 9, 2496. [Google Scholar] [CrossRef] [PubMed]
- Chebeňová-Turcovská, V.; Ženišová, K.; Kuchta, T.; Pangallo, D.; Brežná, B. Culture-independent detection of microorganisms in traditional Slovakian bryndza cheese. Int. J. Food Microbiol. 2011, 150, 73–78. [Google Scholar] [CrossRef]
- Kačániová, M.; Terentjeva, M.; Kunová, S.; Haščík, P.; Kowalczewski, P.Ł.; Štefániková, J. Diversity of microbiota in Slovak summer ewes’ cheese “Bryndza”. Open Life Sci. 2021, 16, 277–286. [Google Scholar] [CrossRef]
- Tillisch, K.; Labus, J.; Kilpatrick, L.; Jiang, Z.; Stains, J.; Ebrat, B.; Guyonnet, D.; Legrain–Raspaud, S.; Trotin, B.; Naliboff, B.; et al. Consumption of Fermented Milk Product With Probiotic Modulates Brain Activity. Gastroenterology 2013, 144, 1394–1401.e4. [Google Scholar] [CrossRef]
- Ghoddusi, H.B.; Tamime, A.Y. Microflora of the intestine. In Encyclopedia of Food Microbiology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 639–645. [Google Scholar]
- Ghosh, T.; Beniwal, A.; Semwal, A.; Navani, N.K. Mechanistic Insights into probiotic properties of lactic acid bacteria associated with ethnic fermented dairy products. Front. Microbiol. 2019, 10, 502. [Google Scholar] [CrossRef]
- Taylor, B.C.; Lejzerowicz, F.; Poirel, M.; Shaffer, J.P.; Jiang, L.; Aksenov, A.; Litwin, N.; Humphrey, G.; Martino, C.; Miller-Montgomery, S.; et al. Consumption of fermented foods is associated with systematic differences in the gut microbiome and metabolome. Msystems 2020, 5, e00901-19. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Type | Sample Acronym | Location | # Of Samples | Sample Designation | pH |
|---|---|---|---|---|---|
| Beverages | |||||
| Aarts | AA | Buryatia Republic | 1 | 25AA | 3.5 |
| Khurunga | KR | Buryatia Republic | 2 | 18KR, 26KR | 3.5–4 |
| Koumiss | KM | Buryatia Republic | 1 | 31KM | 3.5 |
| Shubat | SB | Altai Republic | 1 | 77SB | ND |
| Ayran | AR | Karachay-Cherkessia Republic | 2 | 03AR, 05AR | 4 |
| Stavropol Region | 3 | 06AR, 13AR, 15AR | 4 | ||
| Altai Republic | 1 | 78AR | ND | ||
| Kefir | KF | Stavropol Region | 1 | 07KF | 4 |
| Primorsky Region | 1 | 25KF | 4.5 | ||
| Prostokvasha | PS | Arkhangelsk Region | 1 | 16PS | 4.5 |
| Tula Region | 1 | 17PS | 4.5 | ||
| Dagestan Republic | 6 | 29PS, 37PS, 46PS, 66PS, 69PS, 74PS | 3.5–4.5 | ||
| Ryazhenka | RZ | Stavropol Region | 1 | 13RZ | 4.5 |
| Tula Region | 1 | 17RZ | 5 | ||
| Primorsky Region | 1 | 25RZ | 5 | ||
| Tan | TA | Primorsky Region | 1 | 22TA | 5 |
| Creamy products | |||||
| Chegen | CG | Altai Republic | 2 | 75CG, 76CG | ND |
| Matsoni-like product | MC | Karachay-Cherkessia Republic | 1 | 05MC | 4.5 |
| Stavropol Region | 1 | 13MC | 4 | ||
| Sour cream | SN+SM | Stavropol Region | 3 | 06SN, 07SN, 15SN | 5 |
| Kabardino-Balkarian Republic | 1 | 14SN | 4.5 | ||
| Tula Region | 1 | 20SN | 5 | ||
| Buryatia Republic | 2 | 20SM, 22SM | 4 | ||
| Cottage cheese | TG | Karachay-Cherkessia Republic | 2 | 03TG, 05TG | 4–5 |
| Stavropol Region | 3 | 06TG, 07TG, 15TG | 4–5.5 | ||
| Kabardino-Balkarian Republic | 1 | 14TG | 4 | ||
| Arkhangelsk Region | 1 | 16TG | 5.5 | ||
| Tula Region | 2 | 17TG, 20TG | 4–5 | ||
| Buryatia Republic | 1 | 30TG | 4.5 | ||
| Cheeses | |||||
| Bryndza | BZ | Karachay-Cherkessia Republic | 2 | 03BZ, 05BZ | 5 |
| Stavropol Region | 2 | 07BZ, 15BZ | 5 | ||
| Kabardino-Balkarian Republic | 1 | 14BZ | 5 | ||
| Salt cheese (suluguni-like) | SU | Karachay-Cherkessia Republic | 2 | 03SU, 05SU | 5 |
| Stavropol Region | 1 | 06SU | 5.5 | ||
| Khuruud | KU | Buryatia Republic | 1 | 29KU | 6.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kochetkova, T.V.; Grabarnik, I.P.; Klyukina, A.A.; Zayulina, K.S.; Elizarov, I.M.; Shestakova, O.O.; Gavirova, L.A.; Malysheva, A.D.; Shcherbakova, P.A.; Barkhutova, D.D.; et al. Microbial Communities of Artisanal Fermented Milk Products from Russia. Microorganisms 2022, 10, 2140. https://doi.org/10.3390/microorganisms10112140
Kochetkova TV, Grabarnik IP, Klyukina AA, Zayulina KS, Elizarov IM, Shestakova OO, Gavirova LA, Malysheva AD, Shcherbakova PA, Barkhutova DD, et al. Microbial Communities of Artisanal Fermented Milk Products from Russia. Microorganisms. 2022; 10(11):2140. https://doi.org/10.3390/microorganisms10112140
Chicago/Turabian StyleKochetkova, Tatiana V., Ilya P. Grabarnik, Alexandra A. Klyukina, Kseniya S. Zayulina, Ivan M. Elizarov, Oksana O. Shestakova, Liliya A. Gavirova, Anastasia D. Malysheva, Polina A. Shcherbakova, Darima D. Barkhutova, and et al. 2022. "Microbial Communities of Artisanal Fermented Milk Products from Russia" Microorganisms 10, no. 11: 2140. https://doi.org/10.3390/microorganisms10112140
APA StyleKochetkova, T. V., Grabarnik, I. P., Klyukina, A. A., Zayulina, K. S., Elizarov, I. M., Shestakova, O. O., Gavirova, L. A., Malysheva, A. D., Shcherbakova, P. A., Barkhutova, D. D., Karnachuk, O. V., Shestakov, A. I., Elcheninov, A. G., & Kublanov, I. V. (2022). Microbial Communities of Artisanal Fermented Milk Products from Russia. Microorganisms, 10(11), 2140. https://doi.org/10.3390/microorganisms10112140