Circadian Rhythms in Dinoflagellates: What Is the Purpose of Synthesis and Destruction of Proteins?

Abstract
:1. Introduction
2. Many Quasi-Independent Oscillators in Single Cells
3. Protein Synthesis and Destruction
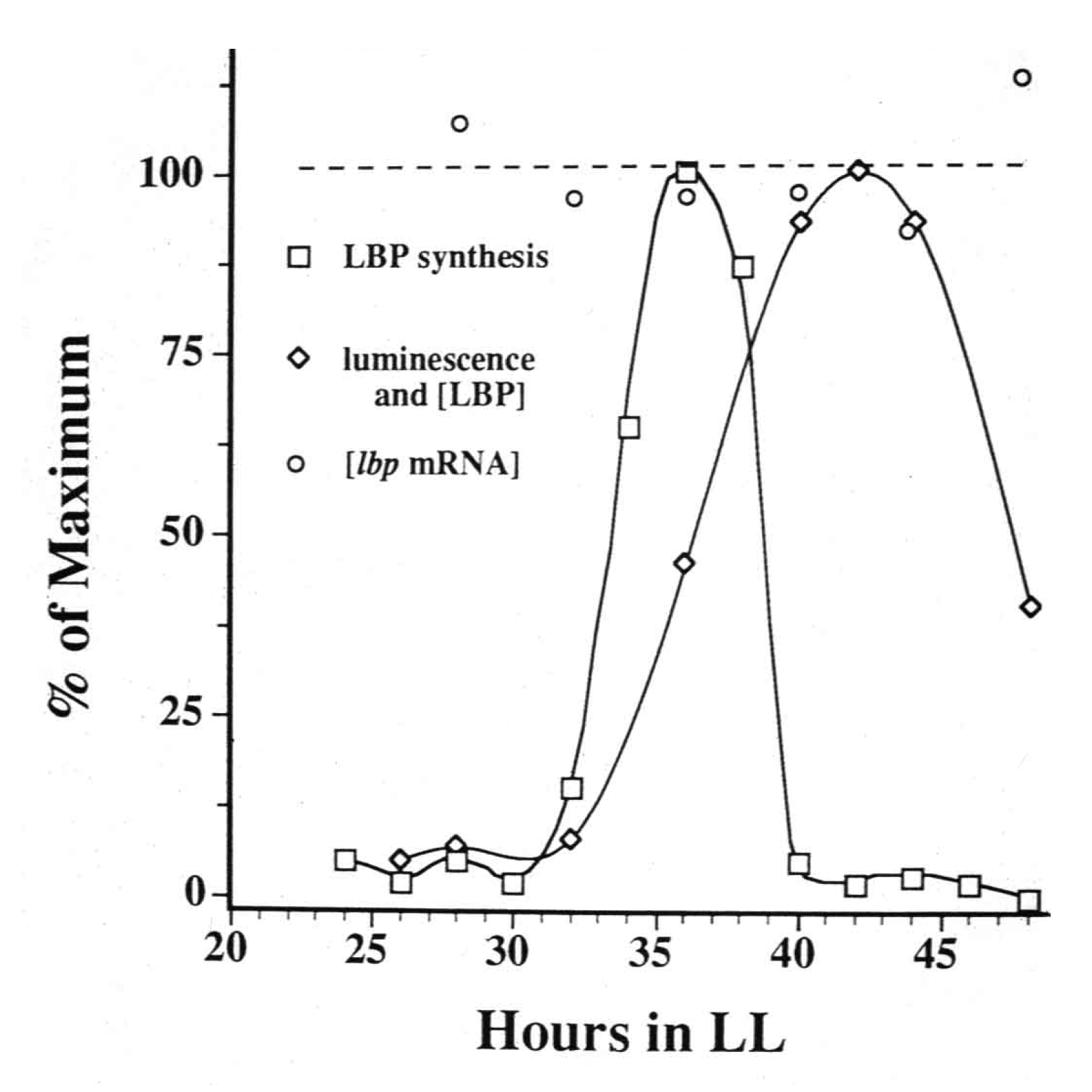
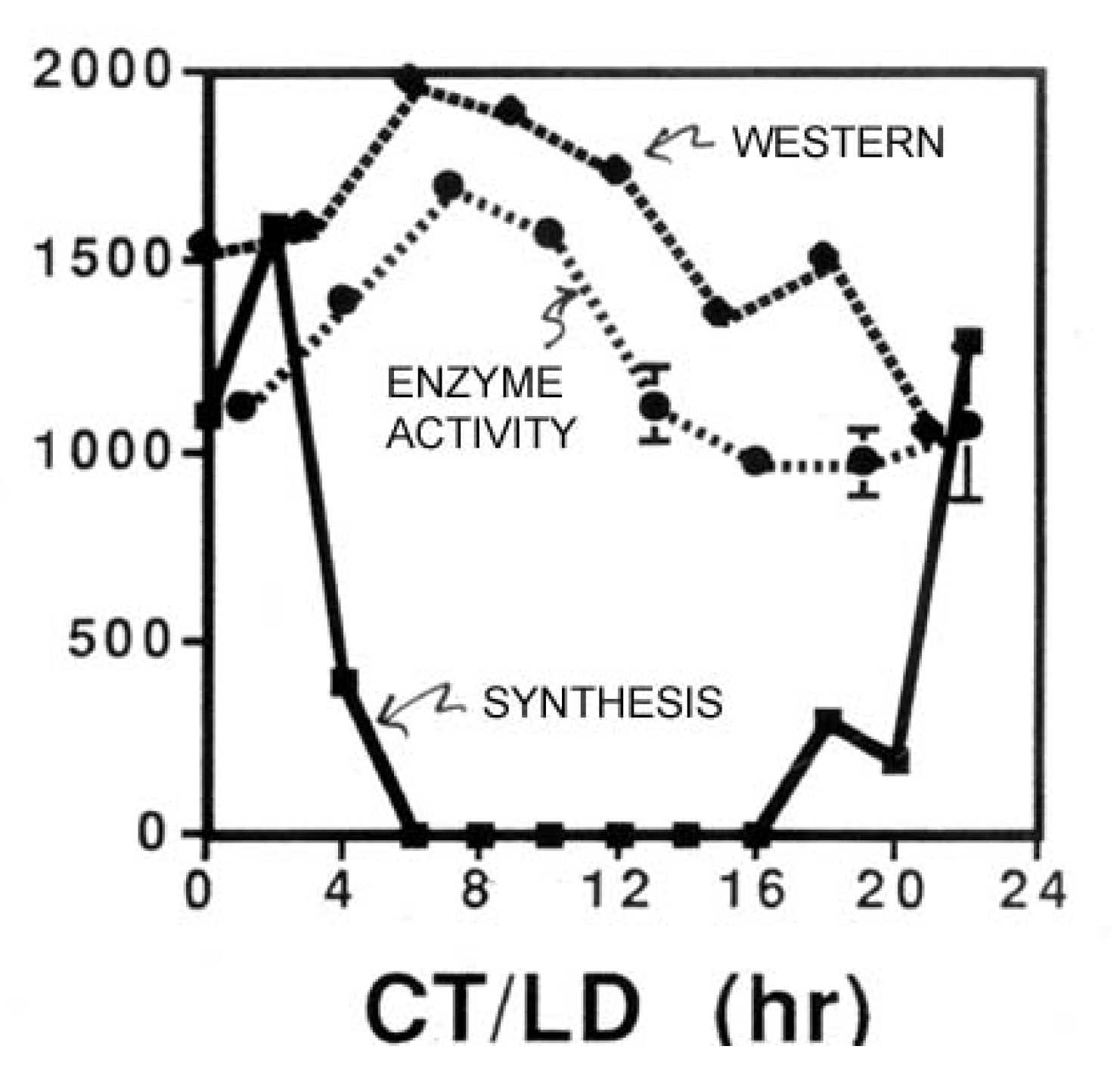

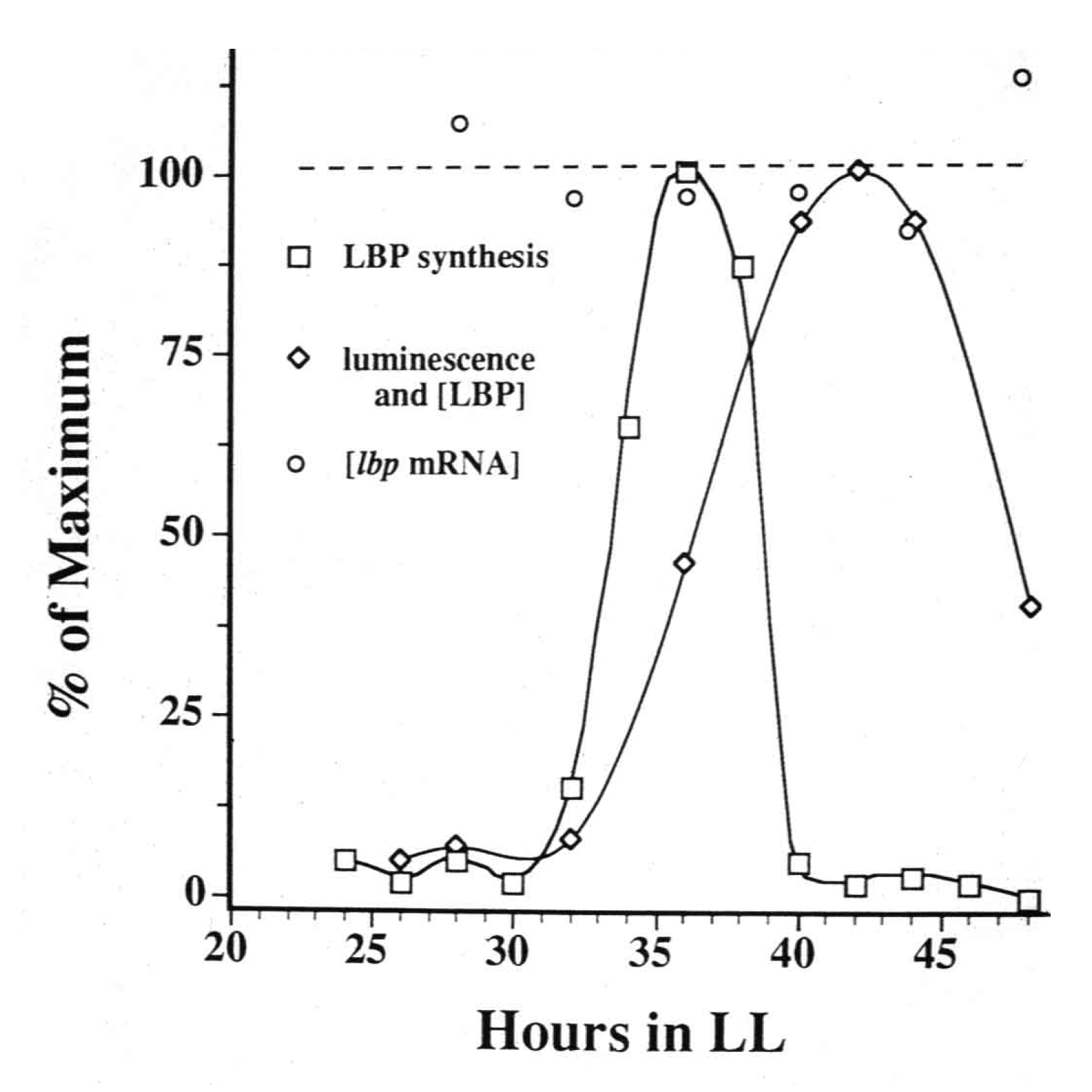{kind=link}
{kind=link}
| Protein MW | Acrophase | ID |
|---|---|---|
| 75 kDa | Early night | LBP |
| 135 kDa | Early night | Luciferase |
| 21 kD | Late night | Unidentified dinoflagellate protein |
| 55 kDa | Late night | Rubisco |
| 32 kD | Early day | PCP |
| 33 kDa | Early day | Oxygen evolving enzyme 1 |
| 45 kDa | Early day | GAPDH |
4. Why Protein Synthesis and Destruction?
5. Conclusions
Acknowledgements
Conflicts of Interest
References
- Valiadi, M.; Iglesias-Rodriguez, D. Understanding bioluminescence in dinoflagellates—How far have we come? Microrganisms 2013, 1, 3–25. [Google Scholar] [CrossRef]
- Hastings, J.W. Progress and Perspectives on Bioluminescence: From Luminous Organisms to Molecular Mechanisms. In Analytical Chemiluminescence and Bioluminescence: Past, Present and Future; Roda, A., Ed.; Royal Society of Chemistry: Cambridge, UK, 2011; pp. 91–112. [Google Scholar]
- Hastings, J.W. Bioluminescence. In Cell Physiology, 4th ed.; Sperelakis, N., Ed.; Academic Press: New York, NY, USA, 2011; pp. 925–947. [Google Scholar]
- Wilson, T.; Hastings, J.W. Bioluminescence: Living Lights, Lights for Living; Harvard University Press: Cambridge, MA, USA, 2013; p. 185. [Google Scholar]
- Hastings, J.W. The Gonyaulax clock at 50: Translational control of circadian expression. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 141–144. [Google Scholar] [CrossRef]
- Markovic, P.; Roenneberg, T.; Morse, D. Phased protein synthesis at several circadian times does not change protein levels in Gonyaulax polyedra. J. Biol. Rhythms 1996, 11, 57–67. [Google Scholar] [CrossRef]
- Nassoury, N.; Hastings, J. Woodland and Morse, David Biochemistry and Circadian Regulation of Output from the Gonyaulax Clock: Are There Many Clocks or Simply Many Hands? In The Circadian Clock in Eukaryotic Microbes; Kippert, F., Ed.; Landes Bioscience: Austin, TX, USA, 2005. [Google Scholar]
- Roenneberg, T.; Morse, D. Two circadian oscillators in one cell. Nature 1993, 362, 362–364. [Google Scholar] [CrossRef]
- Dunlap, J.; Hastings, J.W. The biological clock in Gonyaulax controls luciferase activity by regulating turnover. J. Biol. Chem. 1981, 256, 10509–10518. [Google Scholar]
- Johnson, C.H.; Roeber, J.; Hastings, J.W. Circadian changes in enzyme concentration account for rhythm of enzyme activity in Gonyaulax. Science 1984, 223, 1428–1430. [Google Scholar]
- Morse, D.; Milos, P.M.; Roux, E.; Hastings, J.W. Circadian regulation of the synthesis of substrate binding protein in the Gonyaulax bioluminescent system involves translational control. Proc. Natl. Acad. Sci. USA 1989, 86, 172–176. [Google Scholar]
- Fagan, T.; Morse, D.; Hastings, J.W. Circadian synthesis of a nuclear-encoded chloroplast glyceraldehyde-3-phosphate dehydrogenase in the dinoflagellate Gonyaulax polyedra is translationally controlled. Biochemistry 1999, 38, 7689–7695. [Google Scholar] [CrossRef]
- Milos, P.; Morse, D.; Hastings, J.W. Circadian control over synthesis of many Gonyaulax proteins is at the translational level. Naturwissenschaften 1990, 77, 87–89. [Google Scholar] [CrossRef]
- McMurry, L. Studies on Properties and Biochemistry of Circadian Rhythms in the bioluminescent dinoflagellate, Gonyaulax polyedra. PhD thesis, Department of Biology, Harvard University, Cambridge, MA, USA, 1970. [Google Scholar]
- Knaust, R.; Urbig, T.; Li, L.; Taylor, W.; Hastings, J.W. The circadian rhythm of bioluminescence in Pyrocystis is not due to differences in the amount of luciferase: A comparative study of three bioluminescent marine dinoflagellates. J. Phycol. 1998, 34, 167–172. [Google Scholar]
- Colepicolo, P.; Roenneberg, T; Morse, D.; Taylor, W.; Hastings, J.W. Circadian regulation of bioluminescence in the dinoflagellate Pyrocystis lunula. J. Phycol. 1993, 29, 173–179. [Google Scholar]
- Hardeland, R. Circadian rhythms of bioluminescence in two species of Pyrocystis (Dinophyta). Measurements in cell populations and in single cells. J. Interdiscipl. Cycle Res. 1982, 13, 49–54. [Google Scholar] [CrossRef]
- Hardeland, R.; Nord, P. Visualization of free-running circadian rhythms in the dinoflagellate Pyrocystis noctiluca. Mar. Behav. Physiol. 1984, 11, 199–207. [Google Scholar] [CrossRef]
- Bhovichitra, M.; Swift, E. Light and dark uptake of nitrate and ammonium by large oceanic dinoflagellates: Pyrocystis noctiluca, Pyrocystis fusiformis, and Dissodinium lunula. Limnol. Oceanogr. 1977, 22, 73–83. [Google Scholar] [CrossRef]
- Raven, J.; Richardson, K. Dinophyte flagella—A cost-benefit-analysis. N. Phytol. 1984, 98, 259–276. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hastings, J.W. Circadian Rhythms in Dinoflagellates: What Is the Purpose of Synthesis and Destruction of Proteins? Microorganisms 2013, 1, 26-32. https://doi.org/10.3390/microorganisms1010026
Hastings JW. Circadian Rhythms in Dinoflagellates: What Is the Purpose of Synthesis and Destruction of Proteins? Microorganisms. 2013; 1(1):26-32. https://doi.org/10.3390/microorganisms1010026
Chicago/Turabian StyleHastings, J. Woodland. 2013. "Circadian Rhythms in Dinoflagellates: What Is the Purpose of Synthesis and Destruction of Proteins?" Microorganisms 1, no. 1: 26-32. https://doi.org/10.3390/microorganisms1010026
APA StyleHastings, J. W. (2013). Circadian Rhythms in Dinoflagellates: What Is the Purpose of Synthesis and Destruction of Proteins? Microorganisms, 1(1), 26-32. https://doi.org/10.3390/microorganisms1010026