Evaluation on Elongation Factor 1 Alpha of Entamoeba histolytica Interaction with the Intermediate Subunit of the Gal/GalNAc Lectin and Actin in Phagocytosis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
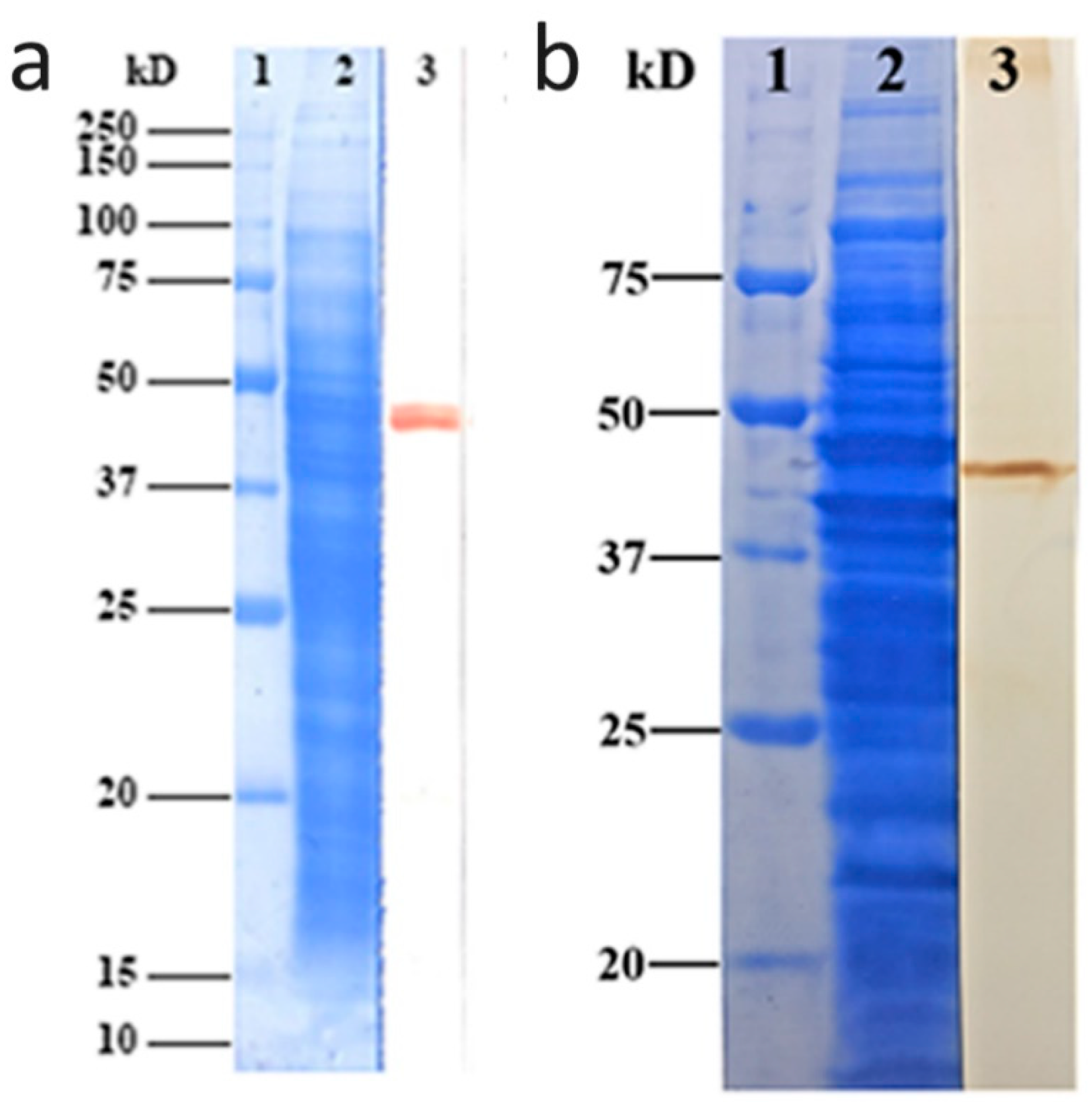
2.1. Specific Binding of EF1a to Igl-C
2.2. Antibody Specificity Confirmed by Western Blotting
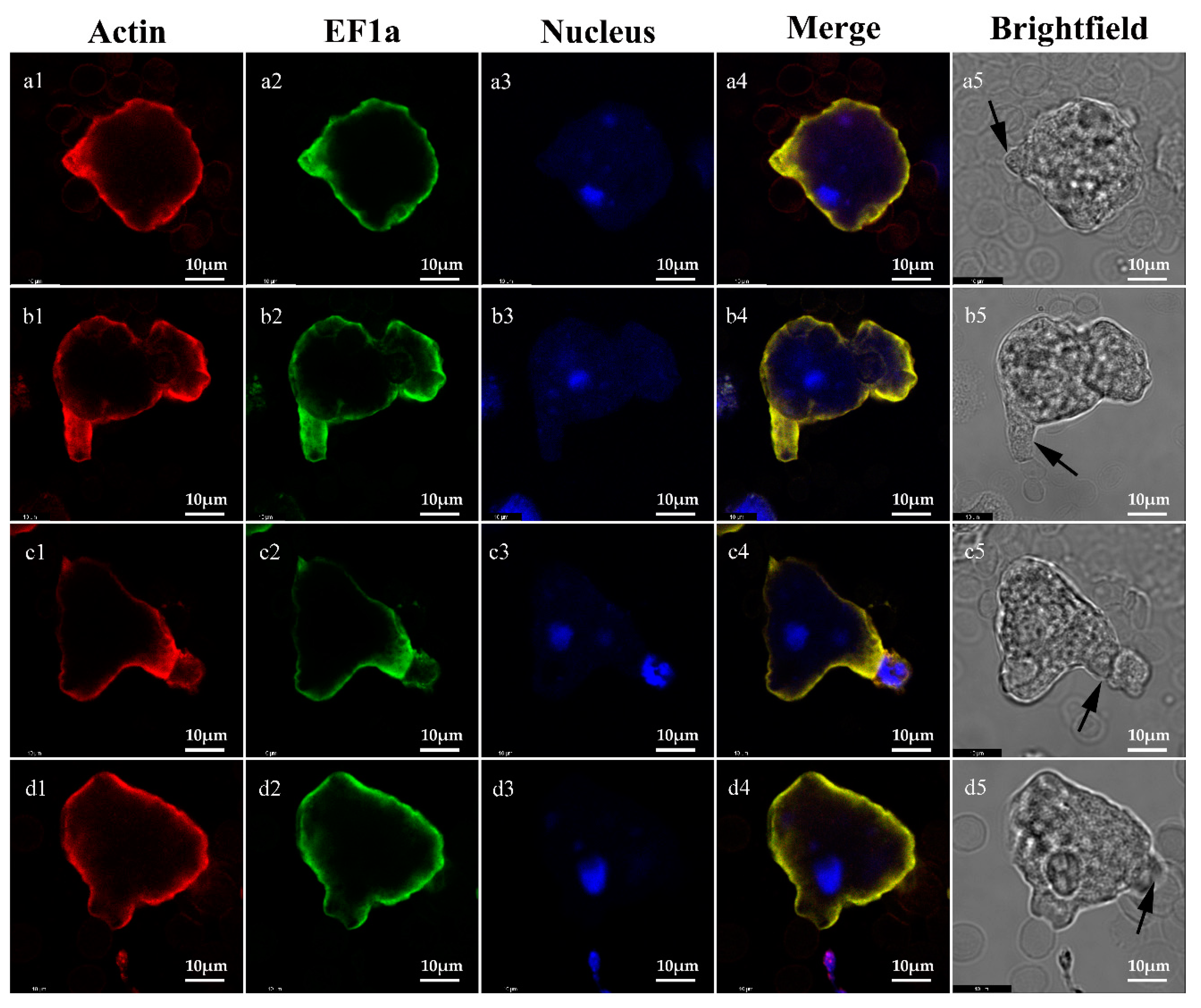
2.3. Localization of Igl and EF1a
2.4. Co-Localization of EF1a and Actin on Trophozoites
2.5. Increased Expression of EF1a and Igl Genes in Trophozoites after the Co-Incubation with CHO Cells
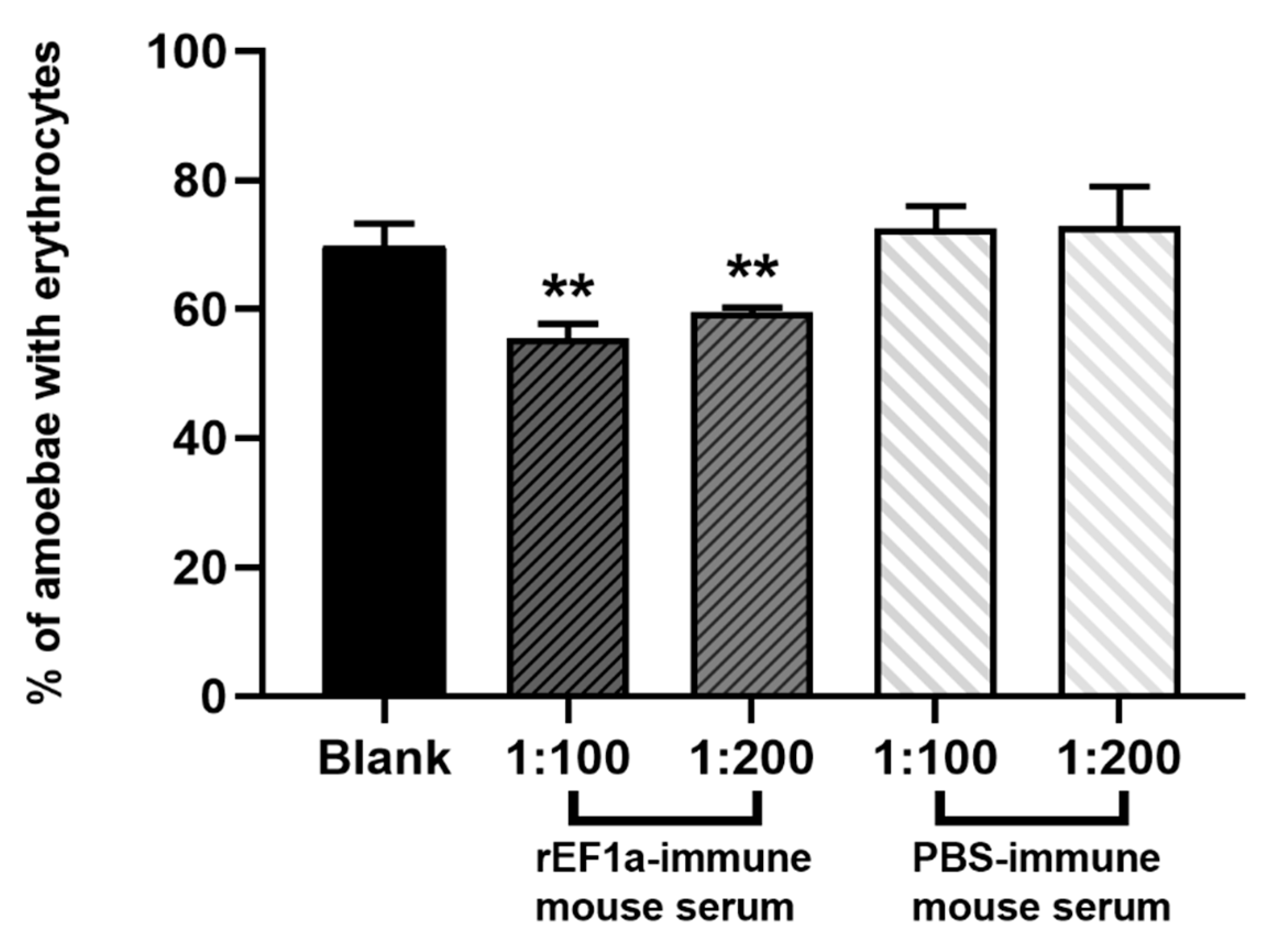
2.6. Erythrophagocytosis Assay
3. Discussion
4. Methods and Materials
4.1. E. histolytica Cell Culture
4.2. Pull-Down Assay and Protein Mass Spectrum
4.3. Cloning of E. histolytica EF1a Gene and Expression of Recombinant Protein
4.4. Biolayer Interferometry Binding Assays
4.5. Monoclonal Antibody Evaluation with the Western Blotting
4.6. Mouse Original Polyclonal Antibodies against rEF1a
4.7. Fluorescence Imaging of Trophozoites
4.8. Co-Incubation of Trophozoites and CHO Cells
4.9. Gene Expression of EF1a and Igl by Real-Time PCR
4.10. Effect of EF1a on the Amebic Pathogenicity with the Erythrophagocytosis Assay
4.11. Ethical Statement
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Stanley, S.L. Amoebiasis. Lancet 2003, 361, 1025–1034. [Google Scholar] [CrossRef]
- Petri, W.A. The bittersweet interface of parasite and host: Lectin-carbohydrate interactions during human invasion by the parasite Entamoeba histolytica. Glycobiology 2000, 10, 1088–1089. [Google Scholar] [CrossRef]
- Cheng, X.J.; Kaneda, Y.; Tachibana, H. A monoclonal antibody against the 150-kDa surface antigen of Entamoeba histolytica inhibits adherence and cytotoxicity to mammalian cells. Med. Ence. Res. 1997, 25, 159–161. [Google Scholar]
- Cheng, X.J.; Tachibana, H. Protection of hamsters from amebic liver abscess formation by immunization with the 150- and 170-kDa surface antigens of Entamoeba histolytica. Parasitol. Res. 2001, 87, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.J.; Tsukamoto, H.; Kaneda, Y.; Tachibana, H. Identification of the 150-kDa surface antigen of Entamoeba histolytica as a galactose- and N-acetyl-d-galactosamine-inhibitable lectin. Parasitol. Res. 1998, 84, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Riis, B.; Rattan, S.I.S.; Clark, B.F.C.; Merrick, W.C. Eukaryotic protein elongation factors. Trends Biochem. Sci. 1990, 15, 420–424. [Google Scholar] [CrossRef]
- EJIRI, S. Moonlighting functions of polypeptide elongation Factor 1: From actin bundling to zinc finger protein R1-associated nuclear localization. Biosci. Biotechnol. Biochem. 2002, 66, 1–21. [Google Scholar] [CrossRef]
- Sasikumar, A.N.; Perez, W.B.; Kinzy, T.G. The many roles of the eukaryotic elongation factor 1 complex. Wiley Interdiscip. Rev. Rna. 2012, 3, 543–555. [Google Scholar] [CrossRef] [Green Version]
- Mateyak, M.K.; Kinzy, T.G. eEF1a: Thinking outside the ribosome. J. Biol. Chem. 2010, 285, 21209–21213. [Google Scholar] [CrossRef] [Green Version]
- Nandan, D.; Lopez, M.; Ban, F.; Huang, M.; Li, Y.; Reiner, N.E.; Cherkasov, A. Indel-based targeting of essential proteins in human pathogens that have close host orthologue(s): Discovery of selective inhibitors for Leishmania donovani elongation factor-1a. Proteins 2010, 67, 53–64. [Google Scholar] [CrossRef]
- Piedra-Quintero, Z.L.; Apodaca-Medina, A.I.; Beltrán-López, E.; León-Sicairos, C.R.; Chávez-Ontiveros, J.; Rendón-Maldonado, J.G.; Ortiz-Navarrete, V.F.; López-Moreno, H.S. Immunoproteomic identification of p29 antigen as the elongation Factor-1α of Leishmania mexicana. Vector Borne Zoonotic Dis. 2015, 15, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Demarta-Gatsi, C.; Rivkin, A.; Di Bartolo, V.; Peronet, R.; Ding, S.; Commere, P.H.; Guillonneau, F.; Bellalou, J.; Brûlé, S.; Abou Karam, P.; et al. Histamine releasing factor and elongation factor 1 alpha secreted via malaria parasites extracellular vesicles promote immune evasion by inhibiting specific T cell responses. Cell. Microbiol. 2019, 21, e13021. [Google Scholar] [CrossRef] [PubMed]
- Skarin, H.; Ringqvist, E.; Hellman, U.; Svärd, S.G. Elongation factor 1-alpha is released into the culture medium during growth of Giardia intestinalis trophozoites. Exp. Parasitol. 2011, 127, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Z.; Wang, Y.; Gadahi, J.A.; Xu, L.; Yan, R.; Song, X.; Li, X. Toxoplasma gondii Elongation factor 1-Alpha (TgEF-1α) is a novel vaccine candidate antigen against Toxoplasmosis. Front. Microbiol. 2017, 8, 168. [Google Scholar] [CrossRef]
- Inomata, A.; Murakoshi, F.; Ishiwa, A.; Takano, R.; Takemae, H.; Sugi, T.; Cagayat Recuenco, F.; Horimoto, T.; Kato, K. Heparin interacts with elongation factor 1α of Cryptosporidium parvum and inhibits invasion. Sci. Rep. 2015, 5, 11599. [Google Scholar] [CrossRef] [Green Version]
- Matsubayashi, M.; Teramoto-Kimata, I.; Uni, S.; Lillehoj, H.S.; Matsuda, H.; Furuya, M.; Tani, H.; Sasai, K. Elongation Factor-1α is a novel protein associated with Host Cell Invasion and a Potential Protective Antigen ofCryptosporidium parvum. J. Biol. Chem. 2013, 288, 34111–34120. [Google Scholar] [CrossRef] [Green Version]
- Alves, L.R.; Oliveira, C.; Goldenberg, S. Eukaryotic translation elongation factor-1 alpha is associated with a specific subset of mRNAs in Trypanosoma cruzi. BMC Microbiol. 2015, 15, 104. [Google Scholar] [CrossRef] [Green Version]
- De Meester, F.; Bracha, R.; Huber, M.; Keren, Z.; Rozenblatt, S.; Mirelman, D. Cloning and characterization of an unusual elongation factor-1α cDNA from Entamoeba histolytica. Mol. Biochem. Parasitol. 1991, 44, 23–32. [Google Scholar]
- Hashimoto, T.; Nakamura, Y.; Nakamura, F.; Shirakura, T.; Adachi, J.; Goto, N.; Okamoto, K.; Hasegawa, M. Protein phylogeny gives a robust estimation for early divergences of eukaryotes: Phylogenetic place of a mitochondria-lacking protozoan, Giardia lamblia. Mol. Biol. Evol. 1994, 11, 65–71. [Google Scholar]
- Gross, S.R.; Kinzy, T.G. Translation elongation factor 1A is essential for regulation of the actin cytoskeleton and cell morphology. Nat. Struct. Mol. Biol. 2005, 12, 772–778. [Google Scholar] [CrossRef]
- Okada, M.; Huston, C.D.; Mann, B.J.; Petri, W.A., Jr.; Kita, K.; Nozaki, T. Proteomic analysis of phagocytosis in the enteric protozoan parasite Entamoeba histolytica. Eukaryot. Cell 2005, 4, 827–831. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Demma, M.; Warren, V.; Dharmawardhane, S.; Condeelis, J. Identification of an actin-binding protein from Dictyostelium as elongation factor 1a. Nature 1990, 347, 494–496. [Google Scholar] [CrossRef] [PubMed]
- Demma, M.; Warren, V.; Hock, R.; Dharmawardhane, S.; Condeelis, J. Isolation of an abundant 50,000-dalton actin filament bundling protein from Dictyostelium amoebae. J. Biol. Chem. 1990, 265, 2286–2291. [Google Scholar] [PubMed]
- Edmonds, B.T. ABP50: An actin-binding elongation factor 1α from Dictyostelium discoideum. J. Cell. Biochem. 1993, 52, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Boss, W.F. Regulation of phosphatidylinositol 4-kinase by the protein activator PIK-A49: Activation requires phosphorylation of PIK-A49. J. Biol. Chem. 1994, 269, 3852–3857. [Google Scholar] [PubMed]
- Raucher, D.; Stauffer, T.; Chen, W.; Shen, K.; Guo, S.; York, J.D.; Sheetz, M.P.; Meyer, T. Phosphatidylinositol 4,5-bisphosphate functions as a second messenger that regulates cytoskeleton–plasma membrane adhesion. Cell 2000, 100, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Nandan, D.; Yi, T.; Lopez, M.; Lai, C.; Reiner, N.E. Leishmania EF-1alpha activates the Src homology 2 domain containing tyrosine phosphatase SHP-1 leading to macrophage deactivation. J. Biol. Chem. 2002, 277, 50190–50197. [Google Scholar] [CrossRef] [Green Version]
- Carelli, J.D.; Sethofer, S.G.; Smith, G.A.; Miller, H.R.; Simard, J.L.; Merrick, W.C.; Jain, R.K.; Ross, N.T.; Taunton, J. Ternatin and improved synthetic variants kill cancer cells by targeting the elongation factor-1A ternary complex. eLife 2015, 4, e10222. [Google Scholar] [CrossRef]
- Sánchez-Murcia, P.A.; Cortés-Cabrera, Á.; Gago, F. Structural rationale for the cross-resistance of tumor cells bearing the A399V variant of elongation factor eEF1a1 to the structurally unrelated Didemnin B, Ternatin, Nannocystin A and Ansatrienin B. J. Ofuter Aided Mol. Des. 2017, 31, 915–928. [Google Scholar] [CrossRef]
- Zhong, D.; Zhang, J.; Yang, S.; Soh, U.J.K.; Buschdorf, J.P.; Zhou, Y.T.; Yang, D.; Low, B.C. The SAM domain of the RhoGAP DLC1 binds EF1a1 to regulate cell migration. J. Cell Sci. 2009, 122, 414–424. [Google Scholar] [CrossRef] [Green Version]
- Nandan, D.; Cherkasov, A.; Sabouti, R.; Yi, T.; Reiner, N.E. Molecular cloning, biochemical and structural analysis of elongation factor-1α from Leishmania donovani: Comparison with the mammalian homologue. Biochem. Biophys. Res. Commun. 2003, 302, 646–652. [Google Scholar] [CrossRef]
- Kobayashi, S.; Imai, E.; Haghighi, A.; Khalifa, S.A.; Tachibana, H.; Takeuchi, T. Axenic cultivation of Entamoeba dispar in newly designed yeast extract-iron-gluconic acid-DIHYRDOXYACETONE-serum medium. J. Parasitol. 2005, 91, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, H.; Cheng, X.J.; Masuda, G.; Horiki, N.; Takeuchi, T. Evaluation of recombinant fragments of Entamoeba histolytica Gal/GalNAc lectin intermediate subunit for serodiagnosis of amebiasis. J. Clin. Microbiol. 2004, 42, 1069–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Tachibana, H.; Kobayashi, X.; Cheng, X.J. Hiwatashi Differentiation of Entamoeba histolytica from E. dispar facilitated by monoclonal antibodies against a 150-kDa surface antigen. Parasitol. Res. 1997, 83, 435–439. [Google Scholar] [CrossRef]
- Tachibana, H.; Kobayashi, S.; Kato, Y.; Nagakura, K.; Kaneda, Y.; Takeuchi, T. Identification of a pathogenic isolate-specific 30,000-Mr antigen of Entamoeba histolytica by using a monoclonal antibody. Infect. Immun. 1990, 58, 955–960. [Google Scholar] [CrossRef] [Green Version]
- Min, X.; Feng, M.; Guan, Y.; Man, S.; Fu, Y.; Cheng, X.; Tachibana, H. evaluation of the C-terminal fragment of Entamoeba histolytica Gal/GalNAc lectin intermediate subunit as a vaccine candidate against amebic liver abscess. PLoS Negl. Trop. Dis. 2016, 10, e0004419. [Google Scholar] [CrossRef] [Green Version]
- Ravdin, J.I.; Croft, B.Y.; Guerrant, R.L. Cytopathogenic mechanisms of Entamoeba histolytica. J. Exp. Med. 1980, 152, 377–390. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Guan, Y.; Feng, M.; Fu, Y.; Tachibana, H.; Cheng, X. Evaluation on Elongation Factor 1 Alpha of Entamoeba histolytica Interaction with the Intermediate Subunit of the Gal/GalNAc Lectin and Actin in Phagocytosis. Pathogens 2020, 9, 702. https://doi.org/10.3390/pathogens9090702
Zhou H, Guan Y, Feng M, Fu Y, Tachibana H, Cheng X. Evaluation on Elongation Factor 1 Alpha of Entamoeba histolytica Interaction with the Intermediate Subunit of the Gal/GalNAc Lectin and Actin in Phagocytosis. Pathogens. 2020; 9(9):702. https://doi.org/10.3390/pathogens9090702
Chicago/Turabian StyleZhou, Hang, Yue Guan, Meng Feng, Yongfeng Fu, Hiroshi Tachibana, and Xunjia Cheng. 2020. "Evaluation on Elongation Factor 1 Alpha of Entamoeba histolytica Interaction with the Intermediate Subunit of the Gal/GalNAc Lectin and Actin in Phagocytosis" Pathogens 9, no. 9: 702. https://doi.org/10.3390/pathogens9090702
APA StyleZhou, H., Guan, Y., Feng, M., Fu, Y., Tachibana, H., & Cheng, X. (2020). Evaluation on Elongation Factor 1 Alpha of Entamoeba histolytica Interaction with the Intermediate Subunit of the Gal/GalNAc Lectin and Actin in Phagocytosis. Pathogens, 9(9), 702. https://doi.org/10.3390/pathogens9090702