Tick-Borne Encephalitis Virus: Seasonal and Annual Variation of Epidemiological Parameters Related to Nymph-to-Larva Transmission and Exposure of Small Mammals

 , , , , and
, , , , and 
Abstract
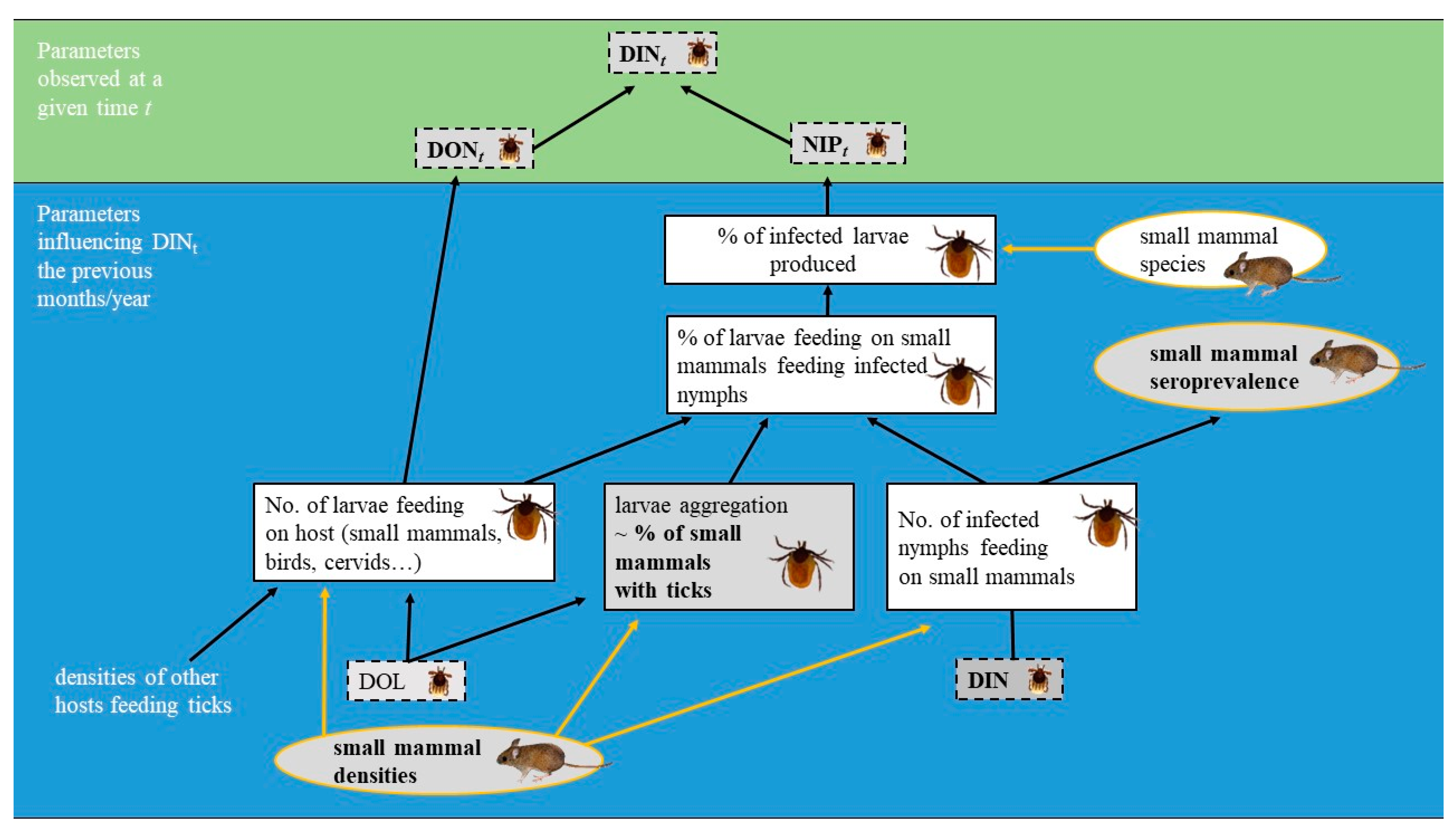
1. Introduction
2. Results
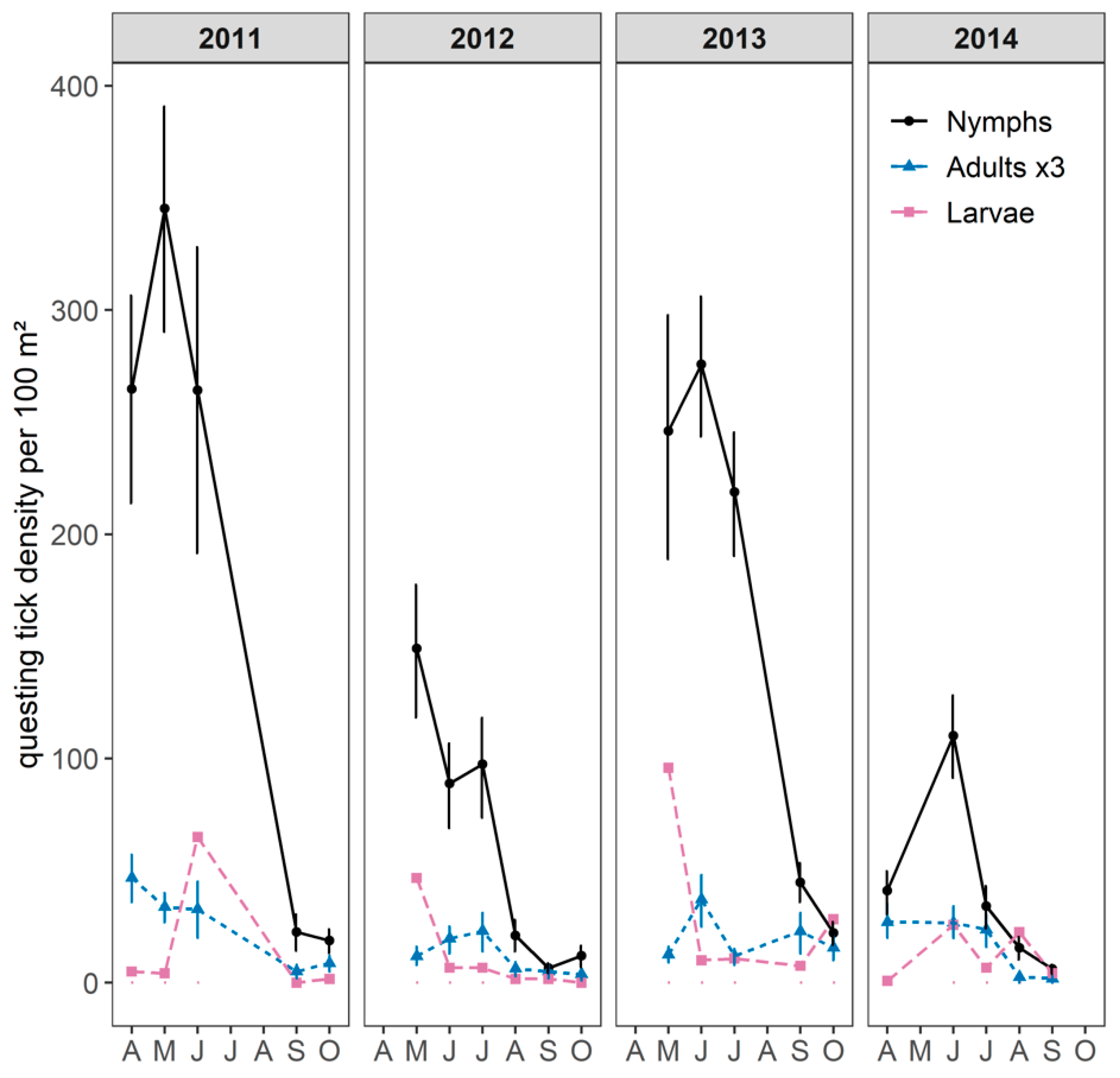
2.1. Questing Tick Densities
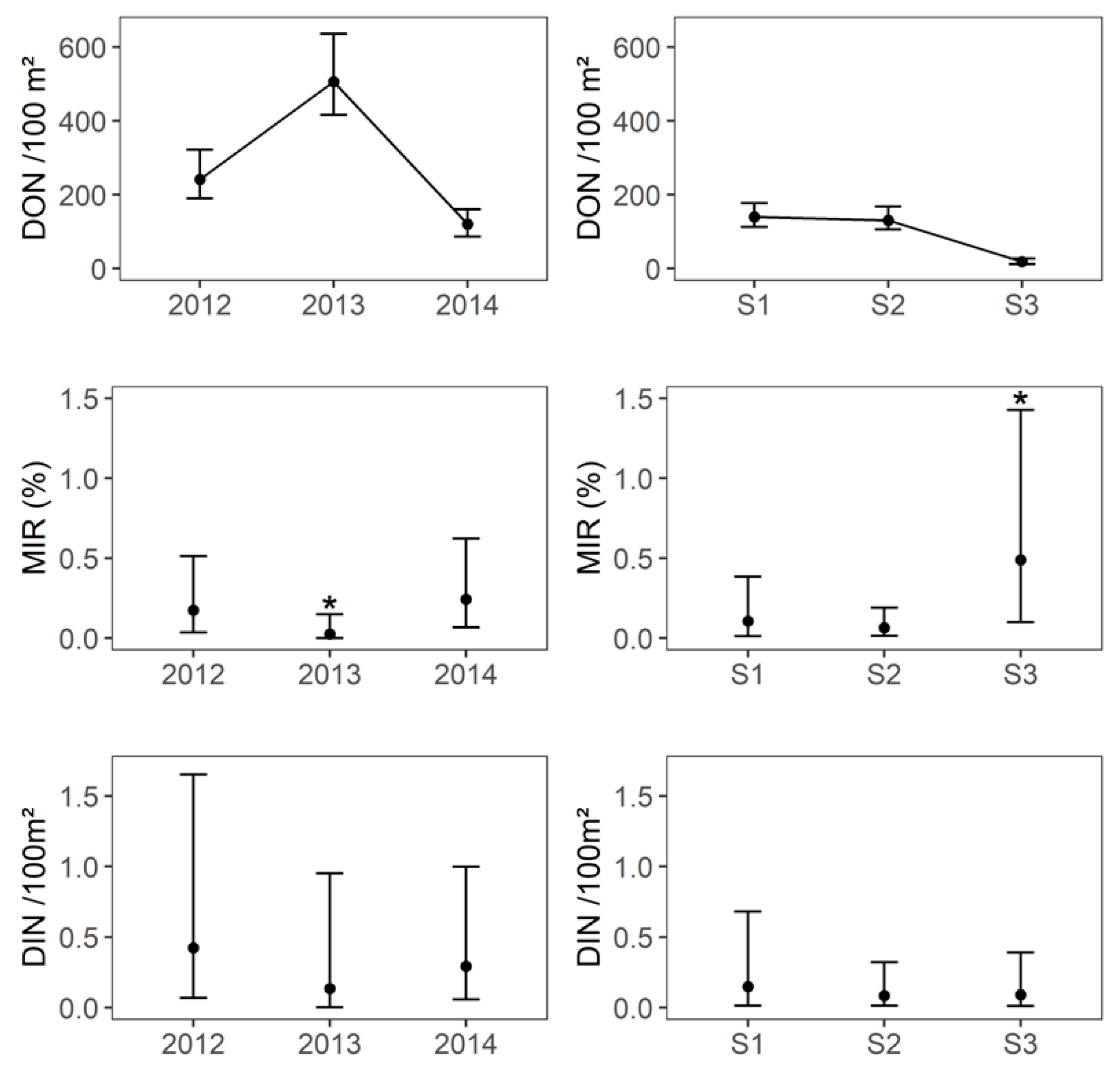
2.2. Tick-Borne Encephalitis Virus Infection Prevalence in Questing Ticks and DIN
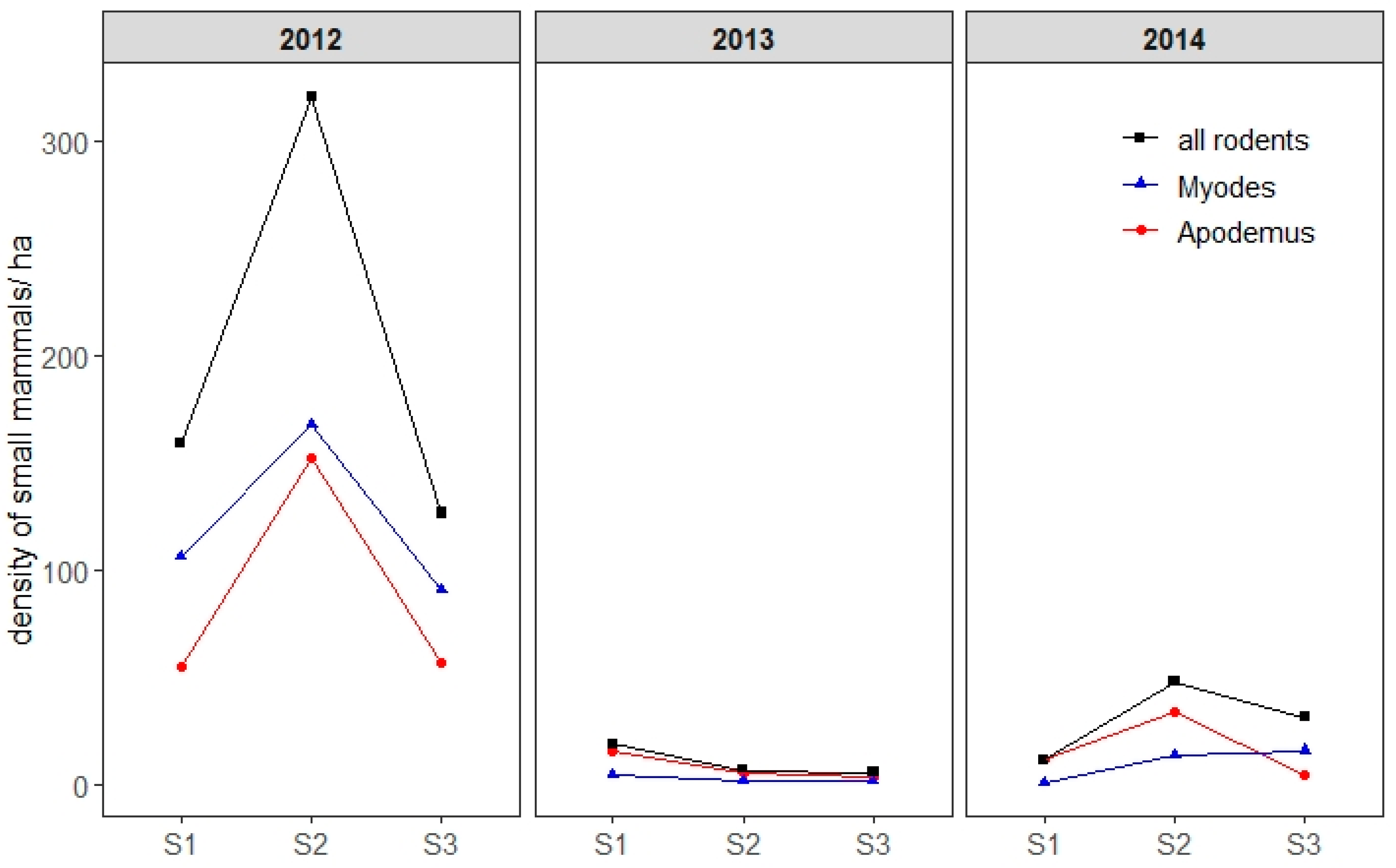
2.3. Small Mammal Abundance
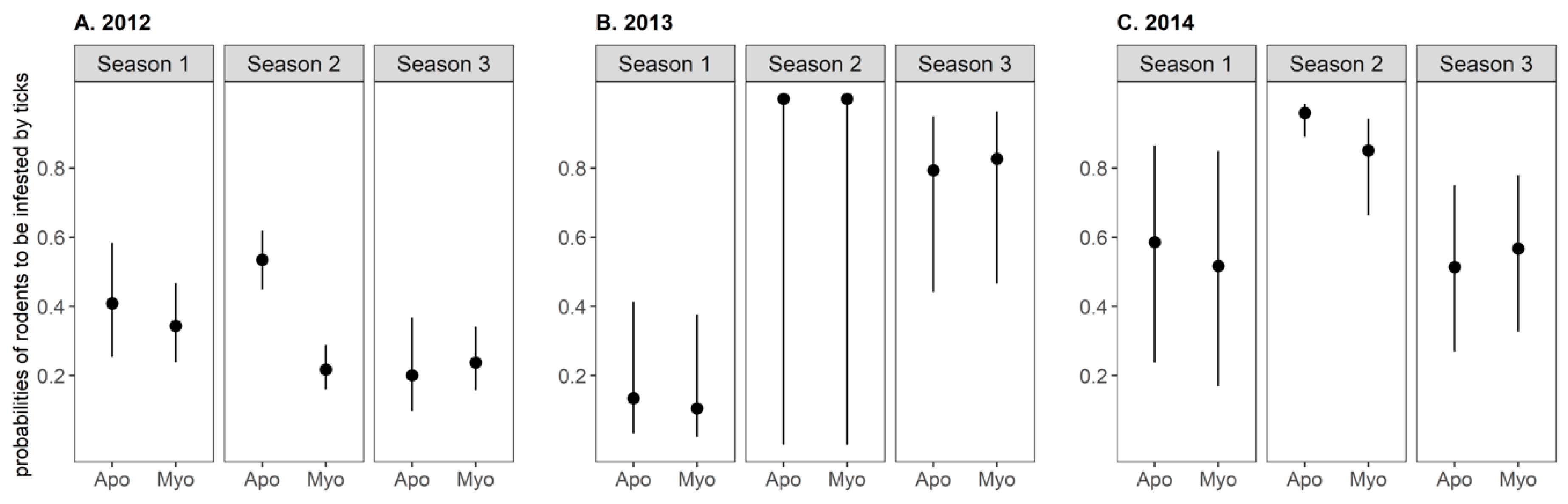
2.4. Tick Infestation on Small Mammals
2.5. Detection of TBEV Antibodies in Small Rodents
2.6. Detection of TBEV in Feeding Ticks
3. Discussion
3.1. Low Circulation of TBEV in the Studied Site
3.2. Seasonal and Inter-Annual Variation of TBEV Prevalence in Ticks and Rodents in 2012–2014
4. Materials and Methods
4.1. Study Area
4.2. Questing Tick Sampling
4.3. Small Mammal Trapping and Sampling
4.4. Ethical Statement
4.5. Laboratory Analysis
4.5.1. TBEV Antibody Detection in Rodents
4.5.2. TBEV Detection in Questing and Fed Ticks
4.6. Descriptive and Statistical Analyses
4.6.1. Definition of Three Seasons
4.6.2. Descriptive and Statistical Analyses of Tick Density and TBEV Infection Prevalence in Ticks
4.6.3. Descriptive and Statistical Analyses of Rodent Density, Tick Infestation, and TBEV Seroprevalence in Rodents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bogovic, P.; Strle, F. Tick-borne encephalitis: A review of epidemiology, clinical characteristics, and management. World J. Clin. Cases 2015, 3, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Süss, J. Tick-borne encephalitis 2010: Epidemiology, risk areas, and virus strains in Europe and Asia—An overview. Ticks Tick-Borne Dis. 2011, 2, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Mannelli, A.; Bertolotti, L.; Gern, L.; Gray, J. Ecology of Borrelia burgdorferi sensu lato in Europe: Transmission dynamics in multi-host systems, influence of molecular processes and effects of climate change. FEMS Microbiol. Rev. 2012, 36, 837–861. [Google Scholar] [CrossRef]
- Gray, J.S.; Kahl, O.; Lane, R.S.; Levin, M.L.; Tsao, J.I. Diapause in ticks of the medically important Ixodes ricinus species complex. Ticks Tick-Borne Dis. 2016, 7, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Perret, J.-L.; Rais, O.; Gern, L. Influence of climate on the proportion of Ixodes ricinus nymphs and adults questing in a tick population. J. Med. Entomol. 2004, 41, 361–365. [Google Scholar] [CrossRef]
- Ernek, E.; Kozuch, O.; Lichard, M.; Nosek, J.; Albrecht, P. Experimental infection of Clethrionomys glareolus and Apodemus flavicollis with tick-borne encephalitis virus. Acta Virol. 1963, 7, 434–436. [Google Scholar]
- Heigl, Z.; Von Zeipel, G. Experimental infection with tick-borne encephalitis virus in Clethrionomys glareolus, Apodemus flavicollis, Apodemus sylvaticus and Mus musculus. 1. Virological studies. Acta Pathol. Microbiol. Scand. 1966, 66, 489–509. [Google Scholar] [CrossRef]
- Kozuch, O.; Chunikhin, S.; Gresikova, M.; Nosek, J.; Kurenkov, V.; Lysý, J. Experimental characteristics of viraemia caused by two strains of tick-borne encephalitis virus in small rodents. Acta Virol. 1981, 25, 219–224. [Google Scholar]
- Chunikhin, S.P.; Kurenkov, V.B. Viraemia in Clethrionomys glareolus -a new ecological marker of tick-borne encephalitis virus. Acta Virol. 1979, 23, 257–260. [Google Scholar]
- Michelitsch, A.; Tews, B.A.; Klaus, C.; Bestehorn-Willmann, M.; Dobler, G.; Beer, M.; Wernike, K. In Vivo Characterization of Tick-Borne Encephalitis Virus in Bank Voles (Myodes glareolus). Viruses 2019, 11, 1069. [Google Scholar] [CrossRef]
- Randolph, S.E. Transmission of tick-borne pathogens between co-feeding ticks: Milan Labuda’s enduring paradigm. Ticks Tick-Borne Dis. 2011, 2, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Hartemink, N.A.; Randolph, S.E.; Davis, S.A.; Heesterbeek, J.A.P. The basic reproduction number for complex disease systems: Defining R0 for tick-borne infections. Am. Nat. 2008, 171, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Labuda, M.; Randolph, S.E. Survival strategy of tick-borne encephalitis virus: Cellular basis and environmental determinants. Zentralblatt Für Bakteriol. 1999, 289, 513–524. [Google Scholar] [CrossRef]
- Randolph, S.E.; Green, R.M.; Peacey, M.F.; Rogers, D.J. Seasonal synchrony: The key to tick-borne encephalitis foci identified by satellite data. Parasitology 2000, 121, 15–23. [Google Scholar] [CrossRef]
- Randolph, S.E. Ticks are not Insects: Consequences of contrasting vector biology for transmission potential. Parasitol. Today 1998, 14, 186–192. [Google Scholar] [CrossRef]
- Harrison, A.; Bennett, N.C. The importance of the aggregation of ticks on small mammal hosts for the establishment and persistence of tick-borne pathogens: An investigation using the R0 model. Parasitology 2012, 139, 1605–1613. [Google Scholar] [CrossRef]
- Woolhouse, M.E.J.; Dye, C.; Etard, J.-F.; Smith, T.; Charlwood, J.D.; Garnett, G.P.; Hagan, P.; Hii, J.L.K.; Ndhlovu, P.D.; Quinnell, R.J.; et al. Heterogeneities in the transmission of infectious agents: Implications for the design of control programs. Proc. Natl. Acad. Sci. USA 1997, 94, 338–342. [Google Scholar] [CrossRef]
- Burri, C.; Bastic, V.; Maeder, G.; Patalas, E.; Gern, L. Microclimate and the zoonotic cycle of tick-borne encephalitis virus in Switzerland. J. Med. Entomol. 2011, 48, 615–627. [Google Scholar] [CrossRef]
- Perkins, S.E.; Cattadori, I.M.; Tagliapietra, V.; Rizzoli, A.P.; Hudson, P.J. Empirical evidence for key hosts in persistence of a tick-borne disease. Int. J. Parasitol. 2003, 33, 909–917. [Google Scholar] [CrossRef]
- Randolph, S.E.; Miklisová, D.; Lysy, J.; Rogers, D.J.; Labuda, M. Incidence from coincidence: Patterns of tick infestations on rodents facilitate transmission of tick-borne encephalitis virus. Parasitology 1999, 118, 177–186. [Google Scholar] [CrossRef]
- Rosà, R.; Pugliese, A.; Ghosh, M.; Perkins, S.E.; Rizzoli, A. Temporal variation of Ixodes ricinus intensity on the rodent host Apodemus flavicollis in relation to local climate and host dynamics. Vector-Borne Zoonotic Dis. 2007, 7, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Perez, G.; Bastian, S.; Chastagner, A.; Agoulon, A.; Plantard, O.; Vourc’h, G.; Butet, A. Ecological factors influencing small mammal infection by Anaplasma phagocytophilum and Borrelia burgdorferi s.l. in agricultural and forest landscapes: Tick-borne infection in small mammals. Environ. Microbiol. 2017, 19, 4205–4219. [Google Scholar] [CrossRef] [PubMed]
- Kiffner, C.; Vor, T.; Hagedorn, P.; Niedrig, M.; Rühe, F. Factors affecting patterns of tick parasitism on forest rodents in tick-borne encephalitis risk areas, Germany. Parasitol. Res. 2011, 108, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Rosà, R.; Tagliapietra, V.; Manica, M.; Arnoldi, D.; Hauffe, H.C.; Rossi, C.; Rosso, F.; Henttonen, H.; Rizzoli, A. Changes in host densities and co-feeding pattern efficiently predict tick-borne encephalitis hazard in an endemic focus in northern Italy. Int. J. Parasitol. 2019, 49, 779–787. [Google Scholar] [CrossRef]
- Perez-Eid, C.; Hannoun, C.; Rodhain, F. The Alsatian tick-borne encephalitis focus: Presence of the virus among ticks and small mammals. Eur. J. Epidemiol. 1992, 8. [Google Scholar] [CrossRef]
- Zöldi, V.; Papp, T.; Rigó, K.; Farkas, J.; Egyed, L. A 4-year study of a natural tick-borne encephalitis virus focus in Hungary, 2010–2013. EcoHealth 2015, 12, 174–182. [Google Scholar] [CrossRef]
- Crespin, L.; Verhagen, R.; Stenseth, N.C.; Yoccoz, N.G.; Prevot-Julliard, A.-C.; Lebreton, J.-D. Survival in fluctuating bank vole populations: Seasonal and yearly variations. Oikos 2002, 98, 467–479. [Google Scholar] [CrossRef]
- Pucek, Z.; Jędrzejewski, W.; Jędrzejewska, B.; Pucek, M. Rodent population dynamics in a primeval deciduous forest (Białowieża National Park) in relation to weather, seed crop, and predation. Acta Theriol. (Warsz.) 1993, 38, 199–232. [Google Scholar] [CrossRef]
- Stenseth, N.C.; Viljugrein, H.; Jędrzejewski, W.; Mysterud, A.; Pucek, Z. Population dynamics of Clethrionomys glareolus and Apodemus flavicollis: Seasonal components of density dependence and density independence. Acta Theriol. (Warsz.) 2002, 47, 39–67. [Google Scholar] [CrossRef]
- Jensen, T.S. Seed production and outbreaks of non-cyclic rodent populations in deciduous forests. Oecologia 1982, 54, 184–192. [Google Scholar] [CrossRef]
- Brugger, K.; Walter, M.; Chitimia-Dobler, L.; Dobler, G.; Rubel, F. Forecasting next season’s Ixodes ricinus nymphal density: The example of southern Germany 2018. Exp. Appl. Acarol. 2018, 75, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Ostfeld, R.S.; Levi, T.; Keesing, F.; Oggenfuss, K.; Canham, C.D. Tick-borne disease risk in a forest food web. Ecology 2018, 99, 1562–1573. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, A.I.; van Duijvendijk, G.L.A.; Swart, A.; Heylen, D.; Jaarsma, R.I.; Jacobs, F.H.H.; Fonville, M.; Sprong, H.; Takken, W. Effect of rodent density on tick and tick-borne pathogen populations: Consequences for infectious disease risk. Parasit. Vectors 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Dizij, A.; Kurtenbach, K. Clethrionomys glareolus, but not Apodemus flavicollis, acquires resistance to Ixodes ricinus, the main European vector of Borrelia burgdorferi. Parasite Immunol. 1995, 17, 177–183. [Google Scholar] [CrossRef]
- Kurtenbach, K.; Kampen, H.; Dizij, A.; Arndt, S.; Seitz, H.M.; Schaible, U.E.; Simon, M.M. Infestation of rodents with larval Ixodes ricinus (Acari; Ixodidae) is an important factor in the transmission cycle of Borrelia burgdorferi s.l. in German woodlands. J. Med. Entomol. 1995, 32, 807–817. [Google Scholar] [CrossRef]
- Pérez, D.; Kneubühler, Y.; Rais, O.; Gern, L. Seasonality of Ixodes ricinus ticks on vegetation and on rodents and Borrelia burgdorferi sensu lato genospecies diversity in two Lyme borreliosis–endemic areas in Switzerland. Vector-Borne Zoonotic Dis. 2012, 12, 633–644. [Google Scholar] [CrossRef]
- Hansmann, Y.; Pierre Gut, J.; Remy, V.; Martinot, M.; Allard Witz, M.; Christmann, D. Tick-borne encephalitis in eastern France. Scand. J. Infect. Dis. 2006, 38, 520–526. [Google Scholar] [CrossRef]
- Velay, A.; Argemi, X.; Wendling, M.-J.; Martinot, M.; Hansmann, Y.; Fafi-Kremer, S. L’encéphalite à tique en France: qu’en savons-nous aujourd’hui ? Rev. Francoph. Lab. 2019, 2019, 34–43. [Google Scholar] [CrossRef]
- Gaumann, R.; Muhlemann, K.; Strasser, M.; Beuret, C.M. High-throughput procedure for tick surveys of tick-borne encephalitis virus and its application in a national surveillance study in Switzerland. Appl. Environ. Microbiol. 2010, 76, 4241–4249. [Google Scholar] [CrossRef]
- Rieille, N.; Bressanelli, S.; Freire, C.C.M.; Arcioni, S.; Gern, L.; Péter, O.; Voordouw, M.J. Prevalence and phylogenetic analysis of tick-borne encephalitis virus (TBEV) in field-collected ticks (Ixodes ricinus) in southern Switzerland. Parasit. Vectors 2014, 7, 443. [Google Scholar] [CrossRef]
- Carpi, G.; Bertolotti, L.; Rosati, S.; Rizzoli, A. Prevalence and genetic variability of tick-borne encephalitis virus in host-seeking Ixodes ricinus in northern Italy. J. Gen. Virol. 2009, 90, 2877–2883. [Google Scholar] [CrossRef] [PubMed]
- Radda, A.; Hofmann, H.; Kunz, C. Experimentelle Infektion uniger heimischer Säugerarten mit dem Frühsommer-Meningo- Enzephalitis Virus. Zbl Bakt Mikr Hyg Abt Orig A 1968, 208, 100–104. [Google Scholar]
- Nikitina, N.; Pchelkina, A.; Kovalevskaya, Y.I. Study of persistence of antibody to tick-borne encephalitis virus in naturally occurring small rodents. Med. Parazit. 1967, 36, 474–478. [Google Scholar]
- Achazi, K.; Růžek, D.; Donoso-Mantke, O.; Schlegel, M.; Ali, H.S.; Wenk, M.; Schmidt-Chanasit, J.; Ohlmeyer, L.; Rühe, F.; Vor, T.; et al. Rodents as sentinels for the prevalence of tick-borne encephalitis virus. Vector-Borne Zoonotic Dis. 2011, 11, 641–647. [Google Scholar] [CrossRef]
- Labuda, M.; Kozuch, O.; Zuffová, E.; Elecková, E.; Hails, R.S.; Nuttall, P.A. Tick-borne encephalitis virus transmission between ticks cofeeding on specific immune natural rodent hosts. Virology 1997, 235, 138–143. [Google Scholar] [CrossRef]
- Grindstaff, J.L.; Brodie, E.D.; Ketterson, E.D. Immune function across generations: Integrating mechanism and evolutionary process in maternal antibody transmission. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 2309–2319. [Google Scholar] [CrossRef]
- Kallio, E.R.; Poikonen, A.; Vaheri, A.; Vapalahti, O.; Henttonen, H.; Koskela, E.; Mappes, T. Maternal antibodies postpone hantavirus infection and enhance individual breeding success. Proc. R. Soc. B Biol. Sci. 2006, 273, 2771–2776. [Google Scholar] [CrossRef]
- Gomez-Chamorro, A.; Heinrich, V.; Sarr, A.; Roethlisberger, O.; Genné, D.; Bregnard, C.; Jacquet, M.; Voordouw, M.J. Maternal antibodies provide bank voles with strain-specific protection against infection by the Lyme disease pathogen. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Cagnacci, F.; Bolzoni, L.; Rosà, R.; Carpi, G.; Hauffe, H.C.; Valent, M.; Tagliapietra, V.; Kazimirova, M.; Koci, J.; Stanko, M.; et al. Effects of deer density on tick infestation of rodents and the hazard of tick-borne encephalitis. I: Empirical assessment. Int. J. Parasitol. 2012, 42, 365–372. [Google Scholar] [CrossRef]
- Burri, C.; Korva, M.; Bastic, V.; Knap, N.; Avšič-Županc, T.; Gern, L. Serological evidence of tick-borne encephalitis virus infection in rodents captured at four sites in Switzerland. J. Med. Entomol. 2012, 49, 436–439. [Google Scholar] [CrossRef]
- Knap, N.; Korva, M.; Dolinšek, V.; Sekirnik, M.; Trilar, T.; Avšič-Županc, T. Patterns of tick-borne encephalitis virus infection in rodents in Slovenia. Vector-Borne Zoonotic Dis. 2012, 12, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Tonteri, E.; Jääskeläinen, A.E.; Tikkakoski, T.; Voutilainen, L.; Niemimaa, J.; Henttonen, H.; Vaheri, A.; Vapalahti, O. Tick-borne encephalitis virus in wild rodents in winter, Finland, 2008–2009. Emerg. Infect. Dis. 2011, 17, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Bakhvalova, V.N.; Potapova, O.F.; Panov, V.V.; Morozova, O.V. Vertical transmission of tick-borne encephalitis virus between generations of adapted reservoir small rodents. Virus Res. 2009, 140, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Kozuch, O.; Gresíková, M.; Nosek, J.; Lichard, M.; Sekeyová, M. The role of small rodents and hedgehogs in a natural focus of tick-borne encephalitis. Bull. World Health Organ. 1967, 36 (Suppl 1), 61–66. [Google Scholar]
- Couret, J.; Dyer, M.C.; Mather, T.N.; Han, S.; Tsao, J.I.; Lebrun, R.A.; Ginsberg, H.S. Acquisition of Borrelia burgdorferi infection by larval Ixodes scapularis (Acari: Ixodidae) associated with engorgement measures. J. Med. Entomol. 2017, 54, 1055–1060. [Google Scholar] [CrossRef]
- Randolph, S.E.; Gern, L.; Nuttall, P.A. Co-feeding ticks: Epidemiological significance for tick-borne pathogen transmission. Parasitol. Today 1996, 12, 472–479. [Google Scholar] [CrossRef]
- Perez-Eid, C. Les relations tiques—petits mammifères dans le foyer Alsacien d’encéphalite à tiques. Acarologia 1990, 31, 131–141. [Google Scholar]
- Reye, A.L.; Hubschen, J.M.; Sausy, A.; Muller, C.P. Prevalence and seasonality of tick-borne pathogens in questing Ixodes ricinus ticks from Luxembourg. Appl. Environ. Microbiol. 2010, 76, 2923–2931. [Google Scholar] [CrossRef]
- May, K.; Jordan, D.; Fingerle, V.; Strube, C. Borrelia burgdorferi sensu lato and co-infections with Anaplasma phagocytophilum and Rickettsia spp. in Ixodes ricinus in Hamburg, Germany: Borrelia infections in ticks in northern Germany. Med. Vet. Entomol. 2015, 29, 425–429. [Google Scholar] [CrossRef]
- Labuda, M.; Nuttall, P.A.; Kozuch, O.; Elecková, E.; Williams, T.; Zuffová, E.; Sabó, A. Non-viraemic transmission of tick-borne encephalitis virus: A mechanism for arbovirus survival in nature. Experientia 1993, 49, 802–805. [Google Scholar] [CrossRef]
- Mishaeva, N.P.; Erofeeva, N.I. The effect of diapause of the tick Ixodes ricinus (Ixodidae) upon the multiplication of the tick encephalitis virus in them. Parazitologiya 1979, 13, 218–222. [Google Scholar]
- Labuda, M.; Austyn, J.M.; Zuffova, E.; Kozuch, O.; Fuchsberger, N.; Lysy, J.; Nuttall, P.A. Importance of localized skin infection in tick-borne encephalitis virus transmission. Virology 1996, 219, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Zeipel, G.V.; Heigl, Z. Experimental infection with tick-borne encephalitis virus in Clethrionomys glareolus, Apodemus flavicollis, Apodemus sylvaticus and Mus musculus. 2. Serological studies. Acta Pathol. Microbiol. Scand. 1966, 68, 211–229. [Google Scholar] [CrossRef]
- Pérez-Eid, C. Les tiques: Identification, biologie, importance médicale et vétérinaire.; Monographies de microbiologie; TEC & DOC Lavoisier: Paris, France, 2007. [Google Scholar]
- Quéré, J.-P.; le Louarn, H. Les rongeurs de France: Faunistique et biologie; Éditions Quæ: Versailles, France, 2011; ISBN 978-2-7592-1032-9. [Google Scholar]
- Beck, C.; Desprès, P.; Paulous, S.; Vanhomwegen, J.; Lowenski, S.; Nowotny, N.; Durand, B.; Garnier, A.; Blaise-Boisseau, S.; Guitton, E.; et al. A high-performance multiplex immunoassay for serodiagnosis of flavivirus-associated neurological diseases in horses. BioMed Res. Int. 2015, 2015, 1–13. [Google Scholar] [CrossRef]
- Schwaiger, M.; Cassinotti, P. Development of a quantitative real-time RT-PCR assay with internal control for the laboratory detection of tick borne encephalitis virus (TBEV) RNA. J. Clin. Virol. 2003, 27, 136–145. [Google Scholar] [CrossRef]
- Puchhammer-Stöckl, E.; Kunz, C.; Mandl, C.W.; Heinz, F.X. Identification of tick-borne encephalitis virus ribonucleic acid in tick suspensions and in clinical specimens by a reverse transcription-nested polymerase chain reaction assay. Clin. Diagn. Virol. 1995, 4, 321–326. [Google Scholar] [CrossRef]
- Bestehorn, M.; Weigold, S.; Kern, W.V.; Chitimia-Dobler, L.; Mackenstedt, U.; Dobler, G.; Borde, J.P. Phylogenetics of tick-borne encephalitis virus in endemic foci in the upper Rhine region in France and Germany. PLoS ONE 2018, 13, e0204790. [Google Scholar] [CrossRef]
- Cannon, R.M. Sense and sensitivity—Designing surveys based on an imperfect test. Prev. Vet. Med. 2001, 49, 141–163. [Google Scholar] [CrossRef]
- Schnabel, Z.E. The estimation of total fish populations of a lake. Am. Math. Mon. 1938, 45, 348–352. [Google Scholar]
- James, G.; Written, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning; Springer Science Business Media: New York, NY, USA, 2017. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: http://www.R-project.org/ (accessed on 23 June 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Season | No. Infested by Ticks/No. Inspected (%) | No. TBEV-Positive/No. Tested (%) | ||||
|---|---|---|---|---|---|---|---|
| Tot | Myodes | Apodemus | Tot | Myodes | Apodemus | ||
| 2012 | Season 1 | 37/104 (35.6) | 24/72 (33.3) | 13/32 (40.6) | 4/95 (4.2) | 2/65 (3.1) | 2/30 (6.7) |
| Season 2 | 158/359 (44.0) | 54/182 (29.7) | 104/177 (58.8) | 11/349 (3.2) | 5/178 (2.8) | 6/171 (3.5) | |
| Season 3 | 28/121 (23.1) | 22/93 (23.7) | 6/28 (21.4) | 5/97 (5.2) | 4/75 (5.3) | 1/22 (4.5) | |
| 2013 | Season 1 | 2/16 (12.5) | 0/4 (0) | 2/12 (16.7) | 0/16 (0) | 0/4 (0) | 0/12 (0) |
| Season 2 | 11/11 (100) | 3/3 (100) | 8/8 (100) | 1/10 (10.0) | 0/3 (0) | 1/7 (14.3) | |
| Season 3 | 8/10 (80.0) | 2/3 (66.7) | 6/7 (85.7) | 0/10 (0) | 0/3 (0) | 0/7 (0) | |
| 2014 | Season 1 | 4/7 (57.1) | 1/1 (100) | 3/6 (50) | 0/7 (0) | 0/1 (0) | 0/6 (0) |
| Season 2 | 65/69 (94.2) | 17/20 (85.0) | 48/49 (98.0) | 0/60 (0) | 0/16 (0) | 0/44 (0) | |
| Season 3 | 11/20 (55.0) | 8/12 (66.7) | 3/8 (37.5) | 1/20 (5.0) | 1/12 (8.3) | 0/8 (0) | |
| 2012 | 2013 | |
|---|---|---|
| No. of rodent-sessions in which ticks were collected | 67 | 12 |
| Infested with nymphs/total infested rodent-sessions (%) | 3/67 (4.5%) | 6/12 (50.0%) |
| Infested with nymphs and larvae/total infested rodent-sessions (%) | 2/67 (3.0%) | 5/12 (41.7%) |
| Tick infestation per rodent-session infested by ticks | ||
| Mean of larvae (+/− SD) per rodent-session infested by ticks | 2.2 (2.2) | 5.3 (4.8) |
| Median of larvae per rodent-session infested by ticks | 2 | 4.5 |
| Mean of nymphs (+/− SD) per rodent-session infested by ticks | 0.1 (0.3) | 0.6 (0.7) |
| Median of nymphs per rodent-session infested by ticks | 0 | 0.5 |
| Larvae feeding on rodent-sessions infested by nymphs | ||
| Number of larvae (%) feeding on rodent-sessions infested by nymphs | 4/147 (2.7%) | 37/64 (57.8%) |
| Mean number of larvae per rodent-session infested by nymphs | 2 | 7.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bournez, L.; Umhang, G.; Moinet, M.; Richomme, C.; Demerson, J.-M.; Caillot, C.; Devillers, E.; Boucher, J.-M.; Hansmann, Y.; Boué, F.; et al. Tick-Borne Encephalitis Virus: Seasonal and Annual Variation of Epidemiological Parameters Related to Nymph-to-Larva Transmission and Exposure of Small Mammals. Pathogens 2020, 9, 518. https://doi.org/10.3390/pathogens9070518
Bournez L, Umhang G, Moinet M, Richomme C, Demerson J-M, Caillot C, Devillers E, Boucher J-M, Hansmann Y, Boué F, et al. Tick-Borne Encephalitis Virus: Seasonal and Annual Variation of Epidemiological Parameters Related to Nymph-to-Larva Transmission and Exposure of Small Mammals. Pathogens. 2020; 9(7):518. https://doi.org/10.3390/pathogens9070518
Chicago/Turabian StyleBournez, Laure, Gerald Umhang, Marie Moinet, Céline Richomme, Jean-Michel Demerson, Christophe Caillot, Elodie Devillers, Jean-Marc Boucher, Yves Hansmann, Franck Boué, and et al. 2020. "Tick-Borne Encephalitis Virus: Seasonal and Annual Variation of Epidemiological Parameters Related to Nymph-to-Larva Transmission and Exposure of Small Mammals" Pathogens 9, no. 7: 518. https://doi.org/10.3390/pathogens9070518
APA StyleBournez, L., Umhang, G., Moinet, M., Richomme, C., Demerson, J.-M., Caillot, C., Devillers, E., Boucher, J.-M., Hansmann, Y., Boué, F., & Moutailler, S. (2020). Tick-Borne Encephalitis Virus: Seasonal and Annual Variation of Epidemiological Parameters Related to Nymph-to-Larva Transmission and Exposure of Small Mammals. Pathogens, 9(7), 518. https://doi.org/10.3390/pathogens9070518