Isolation of Candidatus Bartonella rousetti and Other Bat-associated Bartonellae from Bats and Their Flies in Zambia


 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Results
2.1. Isolation of Bartonella from Bats
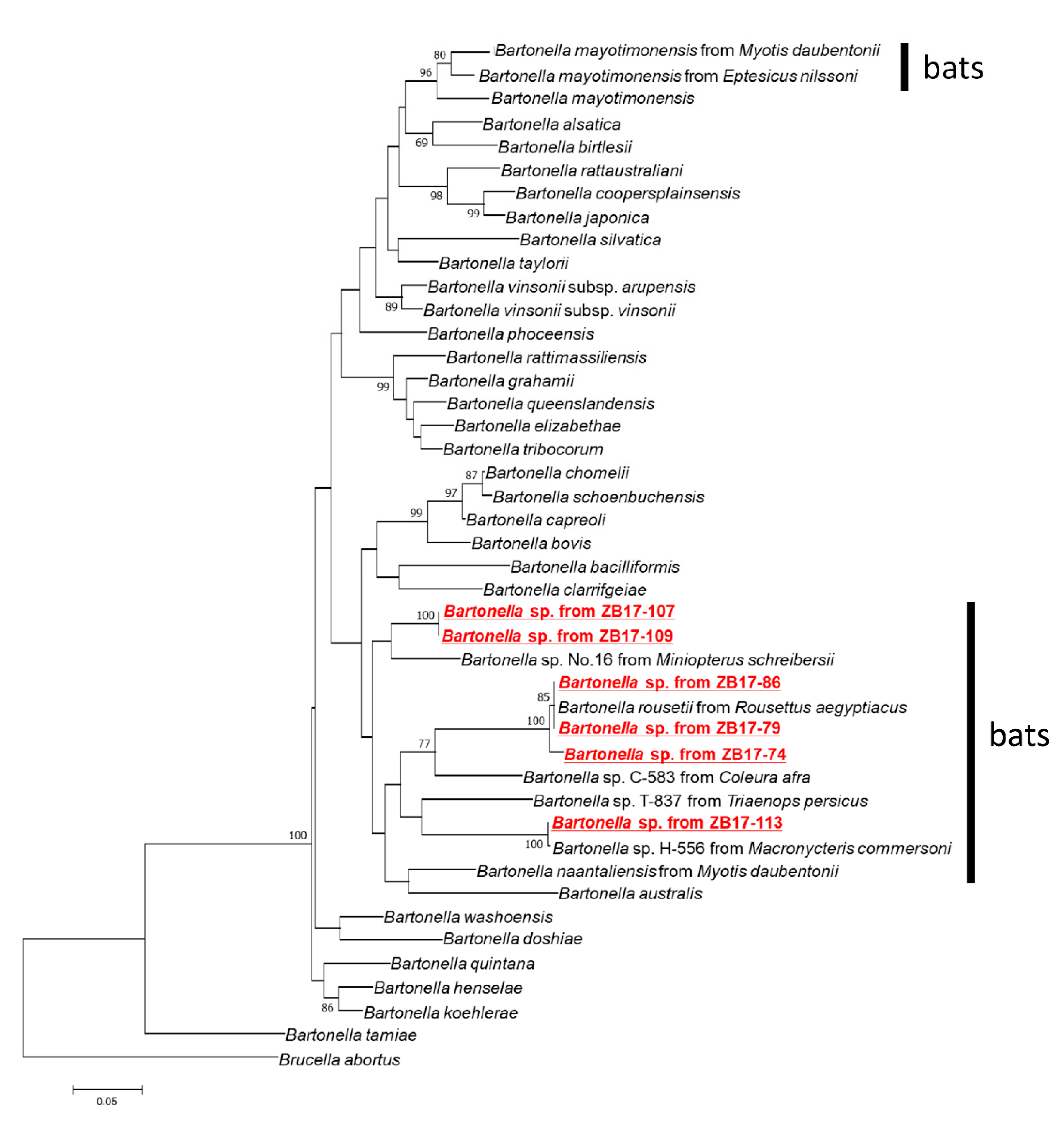
2.2. Molecular Characterization of Bartonella Isolates
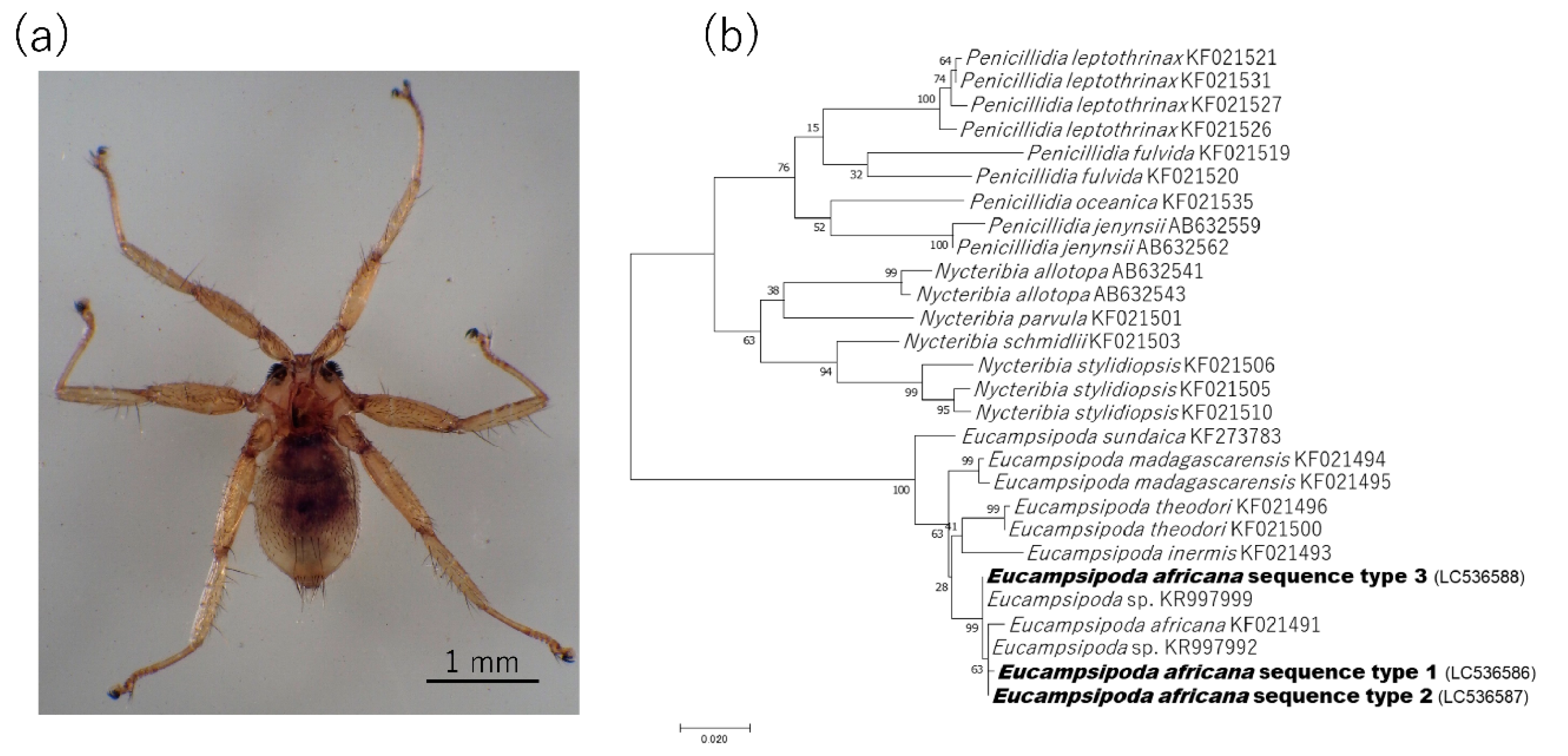
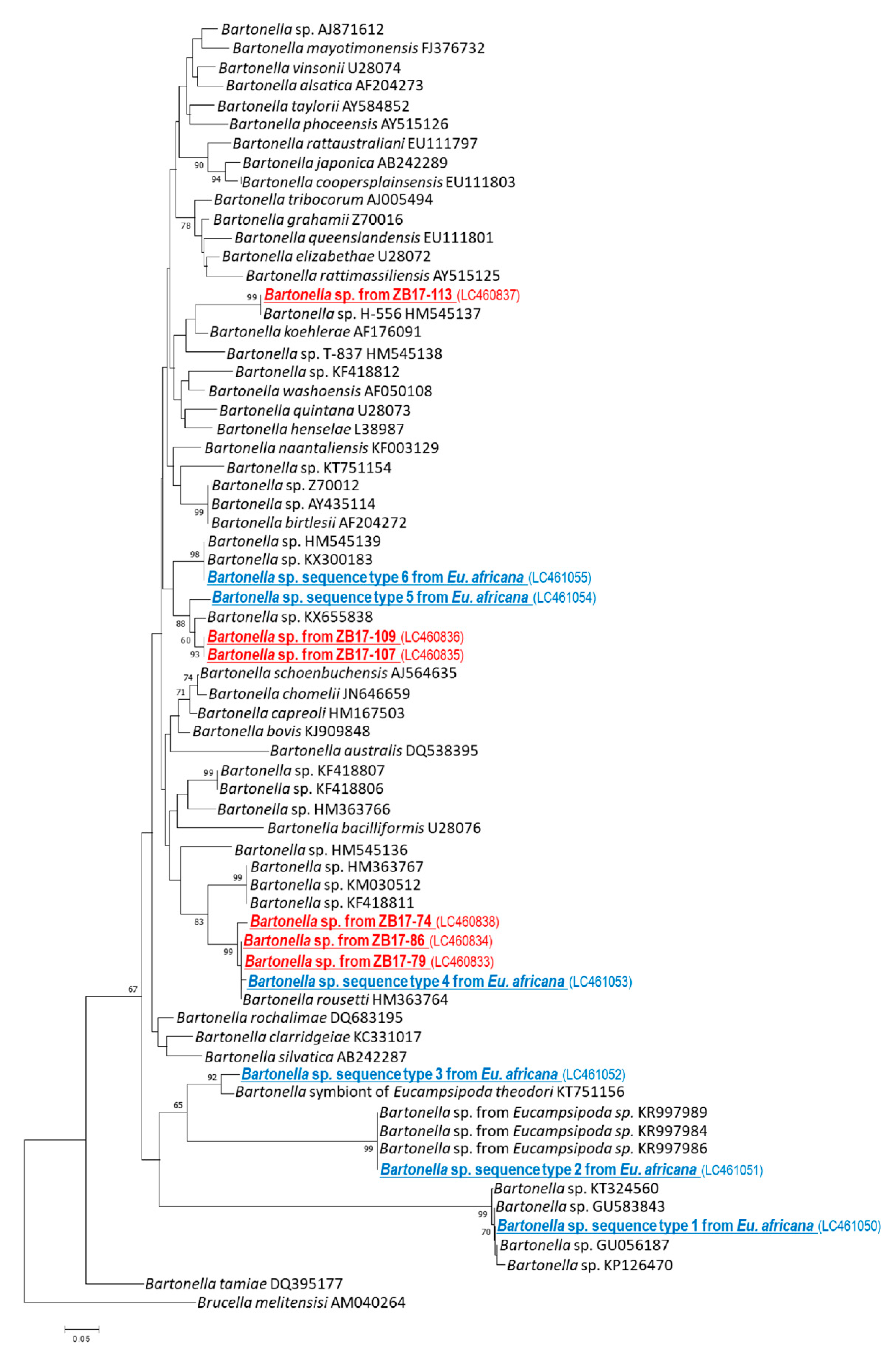
2.3. Detection and Characterization of Bartonella spp. from Bat Flies
3. Discussion
4. Materials and Methods
4.1. Ethics
4.2. Isolation of Bartonella spp.
4.3. Characterization of Bartonella spp.
4.4. Species Identification of Bat Flies and Detection of Bartonella spp.
4.5. Sequencing and Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chomel, B.B.; Boulouis, H.J.; Breitschwerdt, E.B.; Kasten, R.W.; Vayssier-Taussat, M.; Birtles, R.J.; Koehler, J.E.; Dehio, C. Ecological fitness and strategies of adaptation of Bartonella species to their hosts and vectors. Vet. Res. 2009, 40, 29. [Google Scholar] [CrossRef]
- Boulouis, H.J.; Chang, C.C.; Henn, J.B.; Kasten, R.W.; Chomel, B.B. Factors associated with the rapid emergence of zoonotic Bartonella infections. Vet. Res. 2005, 36, 383–410. [Google Scholar] [CrossRef] [PubMed]
- Mühldorfer, K. Bats and bacterial pathogens: A review. Zoonoses Public Health 2013, 60, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Koehler, J.E.; Quinn, F.D.; Berger, T.G.; LeBoit, P.E.; Tappero, J.W. Isolation of Rochalimaea species from cutaneous and osseous lesions of bacillary angiomatosis. N. Engl. J. Med. 1992, 327, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Cardoso, O.; Rodrigues, P.; Cardoso, J.; Machado, J.; Afonso, A.; Bacellar, F.; Marston, E.; Proença, R. Bacillary angiomatosis by Bartonella quintana in an HIV-infected patient. J. Am. Acad. Dermatol. 2000, 42, 299–301. [Google Scholar] [CrossRef]
- Zangwill, K.M.; Hamilton, D.H.; Perkins, B.A.; Regnery, R.L.; Plikaytis, B.D.; Hadler, J.L.; Cartter, M.L.; Wenger, J.D. Cat scratch disease in Connecticut. Epidemiology, risk factors, and evaluation of a new diagnostic test. N. Engl. J. Med. 1993, 329, 8–13. [Google Scholar] [CrossRef]
- Kerkhoff, F.T.; Bergmans, A.M.; van Der Zee, A.; Rothova, A. Demonstration of Bartonella grahamii DNA in ocular fluids of a patient with neuroretinitis. J. Clin. Microbiol. 1999, 37, 4034–4038. [Google Scholar] [CrossRef] [PubMed]
- Kosoy, M.; Morway, C.; Sheff, K.W.; Bai, Y.; Colborn, J.; Chalcraft, L.; Dowell, S.F.; Peruski, L.F.; Maloney, S.A.; Baggett, H.; et al. Bartonella tamiae sp. nov., a newly recognized pathogen isolated from three human patients from Thailand. J. Clin. Microbiol. 2008, 46, 772–775. [Google Scholar] [CrossRef] [PubMed]
- Eremeeva, M.E.; Gerns, H.L.; Lydy, S.L.; Goo, J.S.; Ryan, E.T.; Mathew, S.S.; Ferraro, M.J.; Holden, J.M.; Nicholson, W.L.; Dasch, G.A.; et al. Bacteremia, fever, and splenomegaly caused by a newly recognized bartonella species. N. Engl. J. Med. 2007, 356, 2381–2387. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Roblot, F.; Rolain, J.M.; Besnier, J.M.; Loulergue, J.; Bastides, F.; Choutet, P. First isolation of Bartonella alsatica from a valve of a patient with endocarditis. J. Clin. Microbiol. 2006, 44, 278–279. [Google Scholar] [CrossRef] [PubMed]
- McKee, C.D.; Kosoy, M.Y.; Bai, Y.; Osikowicz, L.M.; Franka, R.; Gilbert, A.T.; Boonmar, S.; Rupprecht, C.E.; Peruski, L.F. Diversity and phylogenetic relationships among Bartonella strains from Thai bats. PLoS ONE 2017, 12, e0181696. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.Y.; Tsigrelis, C.; Baddour, L.M.; Lepidi, H.; Rolain, J.M.; Patel, R.; Raoult, D. Candidatus Bartonella mayotimonensis and endocarditis. Emerg. Infect. Dis. 2010, 16, 500–503. [Google Scholar] [CrossRef] [PubMed]
- Veikkolainen, V.; Vesterinen, E.J.; Lilley, T.M.; Pulliainen, A.T. Bats as reservoir hosts of human bacterial pathogen, Bartonella mayotimonensis. Emerg. Infect. Dis. 2014, 20, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Urushadze, L.; Bai, Y.; Osikowicz, L.; McKee, C.; Sidamonidze, K.; Putkaradze, D.; Imnadze, P.; Kandaurov, A.; Kuzmin, I.; Kosoy, M. Prevalence, diversity, and host associations of Bartonella strains in bats from Georgia (Caucasus). PLoS Negl. Trop. Dis. 2017, 11, e0005428. [Google Scholar] [CrossRef]
- Kosoy, M.; Bai, Y.; Lynch, T.; Kuzmin, I.V.; Niezgoda, M.; Franka, R.; Agwanda, B.; Breiman, R.F.; Rupprecht, C.E. Bartonella spp. in bats, Kenya. Emerg. Infect. Dis. 2010, 16, 1875–1881. [Google Scholar] [CrossRef]
- Kamani, J.; Baneth, G.; Mitchell, M.; Mumcuoglu, K.Y.; Gutiérrez, R.; Harrus, S. Bartonella species in bats (Chiroptera) and bat flies (Nycteribiidae) from Nigeria, West Africa. Vector Borne Zoonotic Dis. 2014, 14, 625–632. [Google Scholar] [CrossRef]
- Bai, Y.; Osinubi, M.O.V.; Osikowicz, L.; McKee, C.; Vora, N.M.; Rizzo, M.R.; Recuenco, S.; Davis, L.; Niezgoda, M.; Ehimiyein, A.M.; et al. Human exposure to novel Bartonella species from contact with fruit bats. Emerg. Infect. Dis. 2018, 24, 2317–2323. [Google Scholar] [CrossRef]
- Han, H.J.; Wen, H.L.; Zhao, L.; Liu, J.W.; Luo, L.M.; Zhou, C.M.; Qin, X.R.; Zhu, Y.L.; Zheng, X.X.; Yu, X.J. Novel Bartonella species in insectivorous bats, Northern China. PLoS ONE 2017, 12, e0167915. [Google Scholar] [CrossRef]
- Bai, Y.; Hayman, D.T.; McKee, C.D.; Kosoy, M.Y. Classification of Bartonella strains associated with straw-colored fruit bats (Eidolon helvum) across Africa using a multi-locus sequence typing platform. PLoS Negl. Trop. Dis. 2015, 9, e0003478. [Google Scholar] [CrossRef]
- Matsumoto, K.; Cook, J.A.; Goethert, H.K.; Telford, S.R. Bartonella sp. Infection of voles trapped from an interior Alaskan site where ticks are absent. J. Wildl. Dis. 2010, 46, 173–178. [Google Scholar] [CrossRef]
- Lin, J.W.; Hsu, Y.M.; Chomel, B.B.; Lin, L.K.; Pei, J.C.; Wu, S.H.; Chang, C.C. Identification of novel Bartonella spp. in bats and evidence of Asian gray shrew as a new potential reservoir of Bartonella. Veter Microbiol. 2012, 156, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, P.; Dsouli, N.; Gomard, Y.; Ramasindrazana, B.; Dick, C.W.; Goodman, S.M. Evolutionary history of Indian Ocean nycteribiid bat flies mirroring the ecology of their hosts. PLoS ONE 2013, 8, e75215. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, M.; Kearney, T.; Seamark, E.C.; Markotter, W. The excreted microbiota of bats: Evidence of niche specialisation based on multiple body habitats. FEMS Microbiol. Lett. 2017, 364, 364. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.A.; Duron, O.; Cordonin, C.; Gomard, Y.; Ramasindrazana, B.; Mavingui, P.; Goodman, S.M.; Tortosa, P. The bacteriome of bat flies (Nycteribiidae) from the Malagasy region: A community shaped by host ecology, bacterial transmission mode, and host-vector specificity. Appl. Environ. Microbiol. 2016, 82, 1778–1788. [Google Scholar] [CrossRef] [PubMed]
- Klangthong, K.; Promsthaporn, S.; Leepitakrat, S.; Schuster, A.L.; McCardle, P.W.; Kosoy, M.; Takhampunya, R. The distribution and diversity of Bartonella species in rodents and their ectoparasites across Thailand. PLoS ONE 2015, 10, e0140856. [Google Scholar] [CrossRef]
- Zouari, S.; Khrouf, F.; M’ghirbi, Y.; Bouattour, A. First molecular detection and characterization of zoonotic Bartonella species in fleas infesting domestic animals in Tunisia. Parasites Vectors 2017, 10, 436. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Nakao, R.; Hang’ombe, B.M.; Sato, K.; Kajihara, M.; Kanchela, S.; Changula, K.; Eto, Y.; Ndebe, J.; Sasaki, M.; et al. Human borreliosis caused by a new world relapsing fever Borrelia-like organism in the old world. Clin. Infect. Dis. 2019, 69, 107–112. [Google Scholar] [CrossRef]
- Billeter, S.A.; Hayman, D.T.; Peel, A.J.; Baker, K.; Wood, J.L.; Cunningham, A.; Suu-Ire, R.; Dittmar, K.; Kosoy, M.Y. Bartonella species in bat flies (Diptera: Nycteribiidae) from western Africa. Parasitology 2012, 139, 324–329. [Google Scholar] [CrossRef]
- Hubert, J.; Erban, T.; Kopecky, J.; Sopko, B.; Nesvorna, M.; Lichovnikova, M.; Schicht, S.; Strube, C.; Sparagano, O. Comparison of microbiomes between red poultry mite populations (Dermanyssus gallinae): Predominance of Bartonella-like bacteria. Microb. Ecol. 2017, 74, 947–960. [Google Scholar] [CrossRef]
- Kešnerová, L.; Moritz, R.; Engel, P. Bartonella apis sp. nov., a honey bee gut symbiont of the class Alphaproteobacteria. Int. J. Syst. Evol. Microbiol. 2016, 66, 414–421. [Google Scholar] [CrossRef]
- Raymann, K.; Moran, N.A. The role of the gut microbiome in health and disease of adult honey bee workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, T.B.; Ferro, M.; Bacci, M.; De Souza, D.J.; Fontana, R.; Delabie, J.H.C.; Silva, A. Bacterial communities in the midgut of Ponerine ants (Hymenoptera: Formicidae: Ponerinae). Sociobiology 2016, 63, 637–644. [Google Scholar] [CrossRef]
- Segers, F.H.; Kešnerová, L.; Kosoy, M.; Engel, P. Genomic changes associated with the evolutionary transition of an insect gut symbiont into a blood-borne pathogen. ISME J. 2017, 11, 1232–1244. [Google Scholar] [CrossRef] [PubMed]
- Changula, K.; Kajihara, M.; Mori-Kajihara, A.; Eto, Y.; Miyamoto, H.; Yoshida, R.; Shigeno, A.; Hang’ombe, B.; Qiu, Y.; Mwizabi, D.; et al. Seroprevalence of filovirus infection of Rousettus aegyptiacus bats in Zambia. J. Infect. Dis. 2018, 218, S312–S317. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Norman, A.F.; Regnery, R.; Jameson, P.; Greene, C.; Krause, D.C. Differentiation of Bartonella-like isolates at the species level by PCR-restriction fragment length polymorphism in the citrate synthase gene. J. Clin. Microbiol. 1995, 33, 1797–1803. [Google Scholar] [CrossRef]
- Colborn, J.M.; Kosoy, M.Y.; Motin, V.L.; Telepnev, M.V.; Valbuena, G.; Myint, K.S.; Fofanov, Y.; Putonti, C.; Feng, C.; Peruski, L. Improved detection of Bartonella DNA in mammalian hosts and arthropod vectors by real-time PCR using the NADH dehydrogenase gamma subunit (nuoG). J. Clin. Microbiol. 2010, 48, 4630–4633. [Google Scholar] [CrossRef]
- Zeaiter, Z.; Liang, Z.; Raoult, D. Genetic classification and differentiation of Bartonella species based on comparison of partial ftsZ gene sequences. J. Clin. Microbiol. 2002, 40, 3641–3647. [Google Scholar] [CrossRef]
- Diaz, M.H.; Bai, Y.; Malania, L.; Winchell, J.M.; Kosoy, M.Y. Development of a novel genus-specific real-time PCR assay for detection and differentiation of Bartonella species and genotypes. J. Clin. Microbiol. 2012, 50, 1645–1649. [Google Scholar] [CrossRef]
- Renesto, P.; Gouvernet, J.; Drancourt, M.; Roux, V.; Raoult, D. Use of rpoB gene analysis for detection and identification of Bartonella species. J. Clin. Microbiol. 2001, 39, 430–437. [Google Scholar] [CrossRef]
- Theodor, O. An Illustrated Catalogue of the Rothschild Collection of Nycteridiidae (Diptera) in the British Museum (Natural History); Whitefriars Press Ltd: London, UK, 1967. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Bat species | ftsZ | gltA | unoG | rpoB | ssrA | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession Number | BLAST Result | Accession Number | BLAST Result | Accession Number | BLAST Result | Accession Number | BLAST Result | Accession Number | BLAST Result | ||
| ZB17-74 | Rousettus aegyptiacus | HM363769 | Bartonella rousetti 99% | MH069695 | Bartonella rousetti B32137 100% | KM387321 | Bartonella rousetti 99% | HM363774 | Bartonella rousetti 96% | KM382247 | Bartonella rousetti 99% |
| ZB17-79 | Rousettus aegyptiacus | HM363769 | Bartonella rousetti 100% | HM363764 | Bartonella rousetti 100% | KM387321 | Bartonella rousetti 100% | HM363774 | Bartonella rousetti 100% | KM382247 | Bartonella rousetti 100% |
| ZB17-86 | Rousettus aegyptiacus | HM363769 | Bartonella rousetti 100% | HM363764 | Bartonella rousetti 100% | KM387321 | Bartonella rousetti 100% | HM363774 | Bartonella rousetti 100% | KM382247 | Bartonella rousetti 100% |
| ZB17-107 | Macronycteris vittatus | KX300182 | Bartonella sp. isolate 44552 96% | KX655838 | Uncultured Bartonella sp. SD-3/2015 98% | KX300163 | Bartonella sp. isolate 44552 97% | KX300164 | Bartonella sp. isolate 44552 96% | KM233461 | Bartonella sp. B23983 97% |
| ZB17-109 | Macronycteris vittatus | KX300182 | Bartonella sp. isolate 44552 96% | KX655838 | Uncultured Bartonella sp. SD-3/2015 98% | KX300163 | Bartonella sp. isolate 44552 97% | KX300164 | Bartonella sp. isolate 44552 95% | KM233461 | Bartonella sp. B23983 96% |
| ZB17-113 | Macronycteris vittatus | KM382254 | Bartonella sp. H-556 99% | HM545137 | Bartonella sp. H-556 100% | KM382252 | Bartonella sp. H-556 100% | EU979536 | Bartonella sp. Cr28649 88% | KM382250 | Bartonella sp. H-556 99% |
| Primer Name | Sequence 5’-3’ | Target Organism | Target Gene | Annealing (°C) | Size Expected (bp) | Reference |
|---|---|---|---|---|---|---|
| fD1 | AGAGTTTGATCCTGGCTCAG | Bartonella spp. | 16S ribosomal DNA | 55 | 1400 | 35 |
| Rp2 | ACGGCTACCTTGTTACGACTT | |||||
| BhCS781.p | GGGGACCAGCTCATGGTGG | gltA | 45 | 380 | 36 | |
| BhCS1137.n | AATGCAAAAAGAACAGTAAACA | |||||
| 1400F | CGCATTGGCTTACTTCGTATG | rpoB | 53 | 860 | 40 | |
| 2300R | GTAGACTGATTAGAACGCTG | |||||
| nuoG F | GGCGTGATTGTTCTCGTTA | nuoG | 55 | 360 | 37 | |
| nuoG R | CACGACCACGGCTATCAAT | |||||
| Bfp1 | ATTAATCTGCAYCGGCCAGA | ftsZ | 55 | 900 | 38 | |
| Bfp2 | ACVGADACACGAATAACACC | |||||
| ssrA-F | GCTATGGTAATAAATGGACAATGAAATAA | ssrA | 60 | 300 | 39 | |
| ssrA-R | GCTTCTGTTGCCAGGTG | |||||
| LCO1490 | GGTCAACAAATCATAAAGATATTGG | Bat fly | COI | 57 | 710 | 42 |
| HCO2198 | TAAACTTCAGGGTGACCAAAAAATCA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Y.; Kajihara, M.; Nakao, R.; Mulenga, E.; Harima, H.; Hang’ombe, B.M.; Eto, Y.; Changula, K.; Mwizabi, D.; Sawa, H.; et al. Isolation of Candidatus Bartonella rousetti and Other Bat-associated Bartonellae from Bats and Their Flies in Zambia. Pathogens 2020, 9, 469. https://doi.org/10.3390/pathogens9060469
Qiu Y, Kajihara M, Nakao R, Mulenga E, Harima H, Hang’ombe BM, Eto Y, Changula K, Mwizabi D, Sawa H, et al. Isolation of Candidatus Bartonella rousetti and Other Bat-associated Bartonellae from Bats and Their Flies in Zambia. Pathogens. 2020; 9(6):469. https://doi.org/10.3390/pathogens9060469
Chicago/Turabian StyleQiu, Yongjin, Masahiro Kajihara, Ryo Nakao, Evans Mulenga, Hayato Harima, Bernard Mudenda Hang’ombe, Yoshiki Eto, Katendi Changula, Daniel Mwizabi, Hirofumi Sawa, and et al. 2020. "Isolation of Candidatus Bartonella rousetti and Other Bat-associated Bartonellae from Bats and Their Flies in Zambia" Pathogens 9, no. 6: 469. https://doi.org/10.3390/pathogens9060469
APA StyleQiu, Y., Kajihara, M., Nakao, R., Mulenga, E., Harima, H., Hang’ombe, B. M., Eto, Y., Changula, K., Mwizabi, D., Sawa, H., Higashi, H., Mweene, A., Takada, A., Simuunza, M., & Sugimoto, C. (2020). Isolation of Candidatus Bartonella rousetti and Other Bat-associated Bartonellae from Bats and Their Flies in Zambia. Pathogens, 9(6), 469. https://doi.org/10.3390/pathogens9060469