Identification of Transmission Routes of Campylobacter and On-Farm Measures to Reduce Campylobacter in Chicken

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Bacteriological Analysis
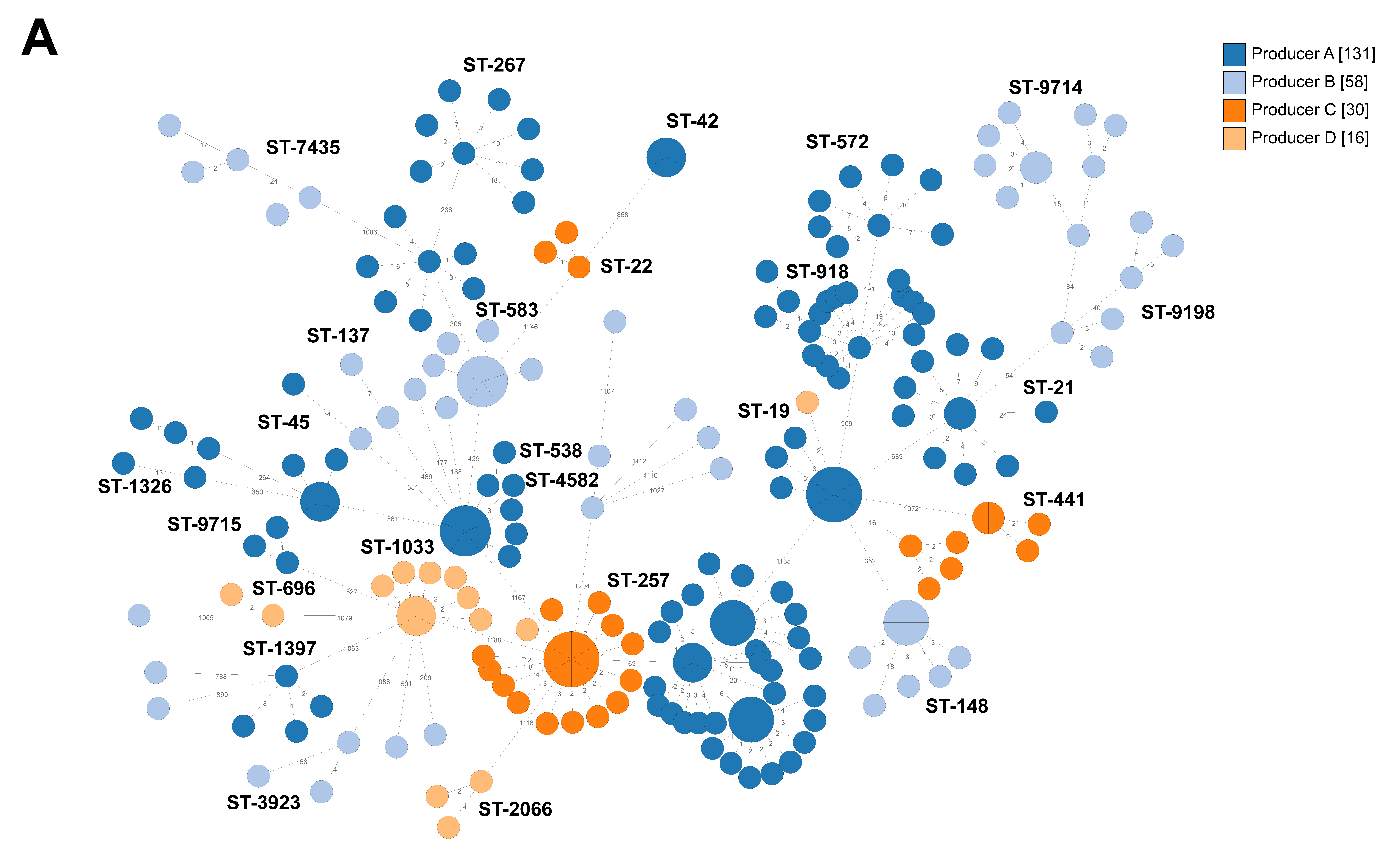
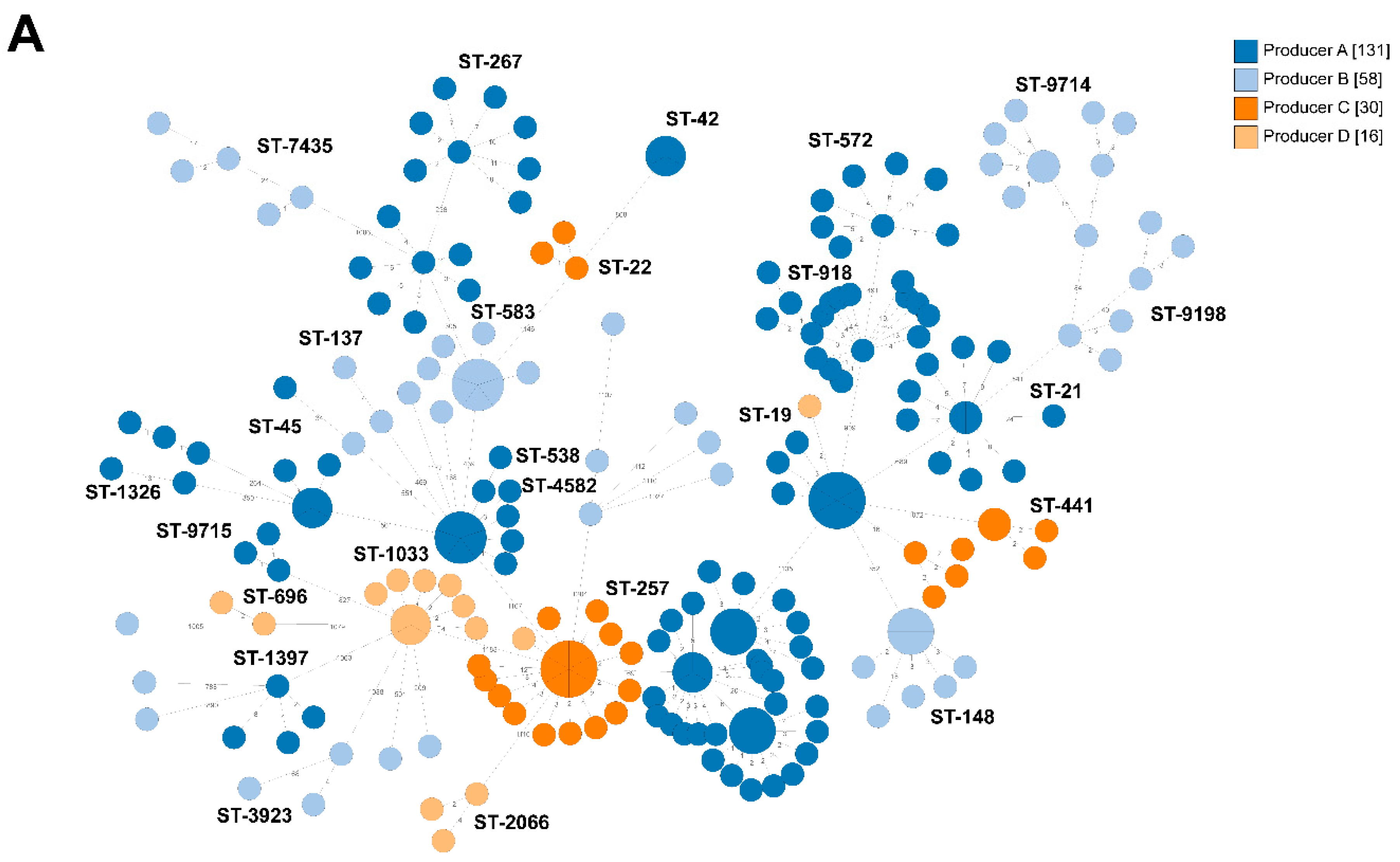
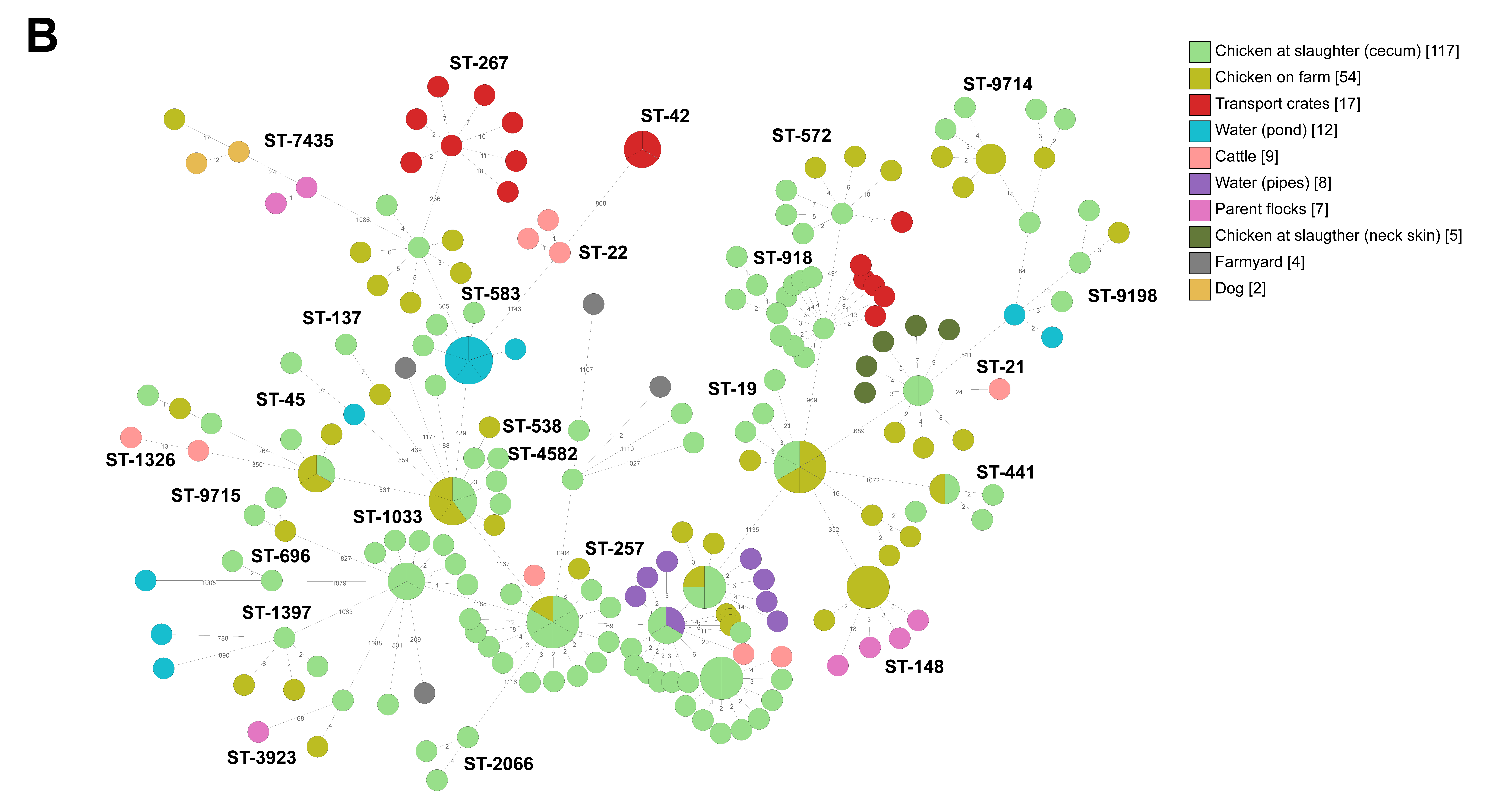
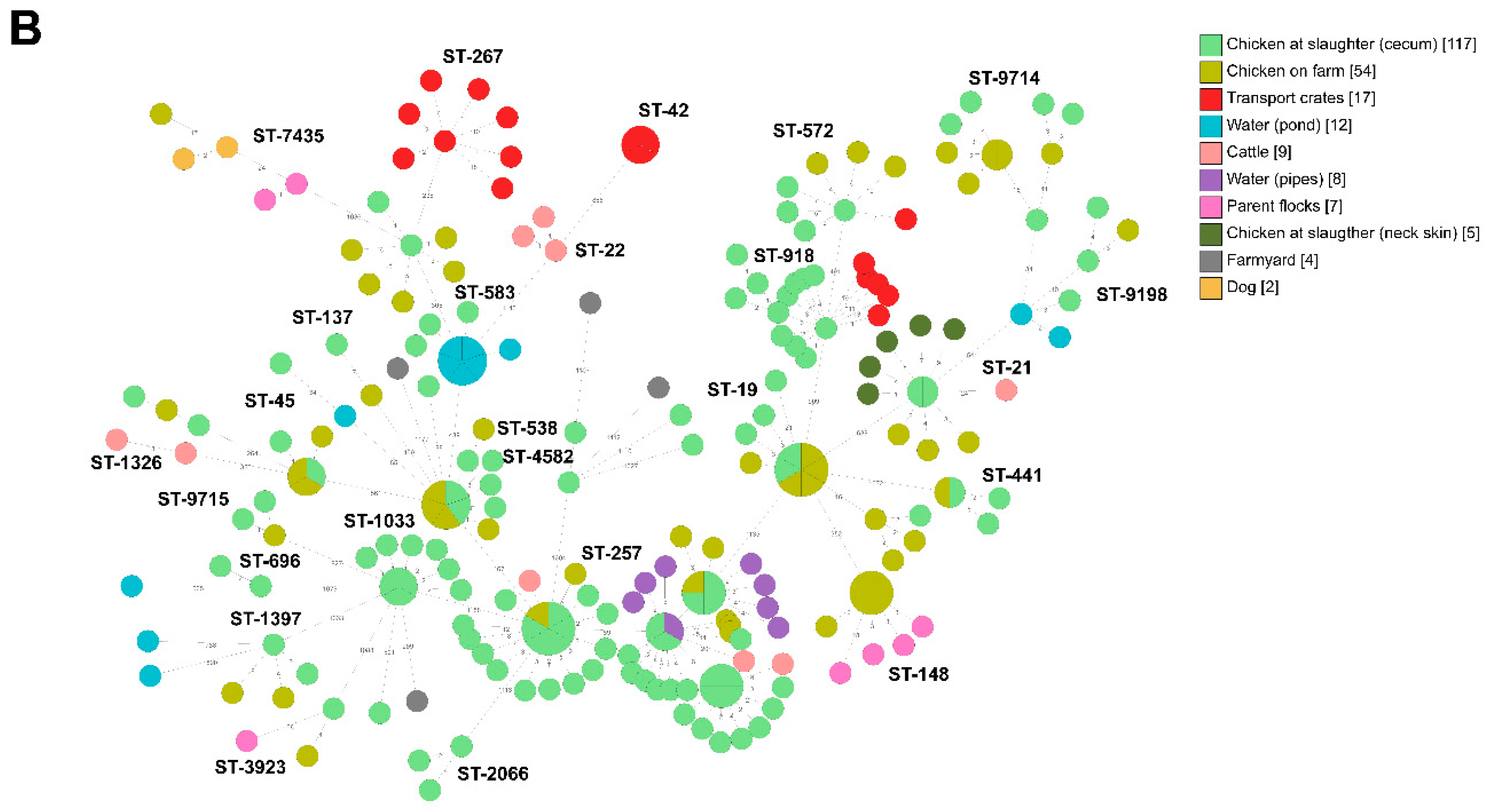
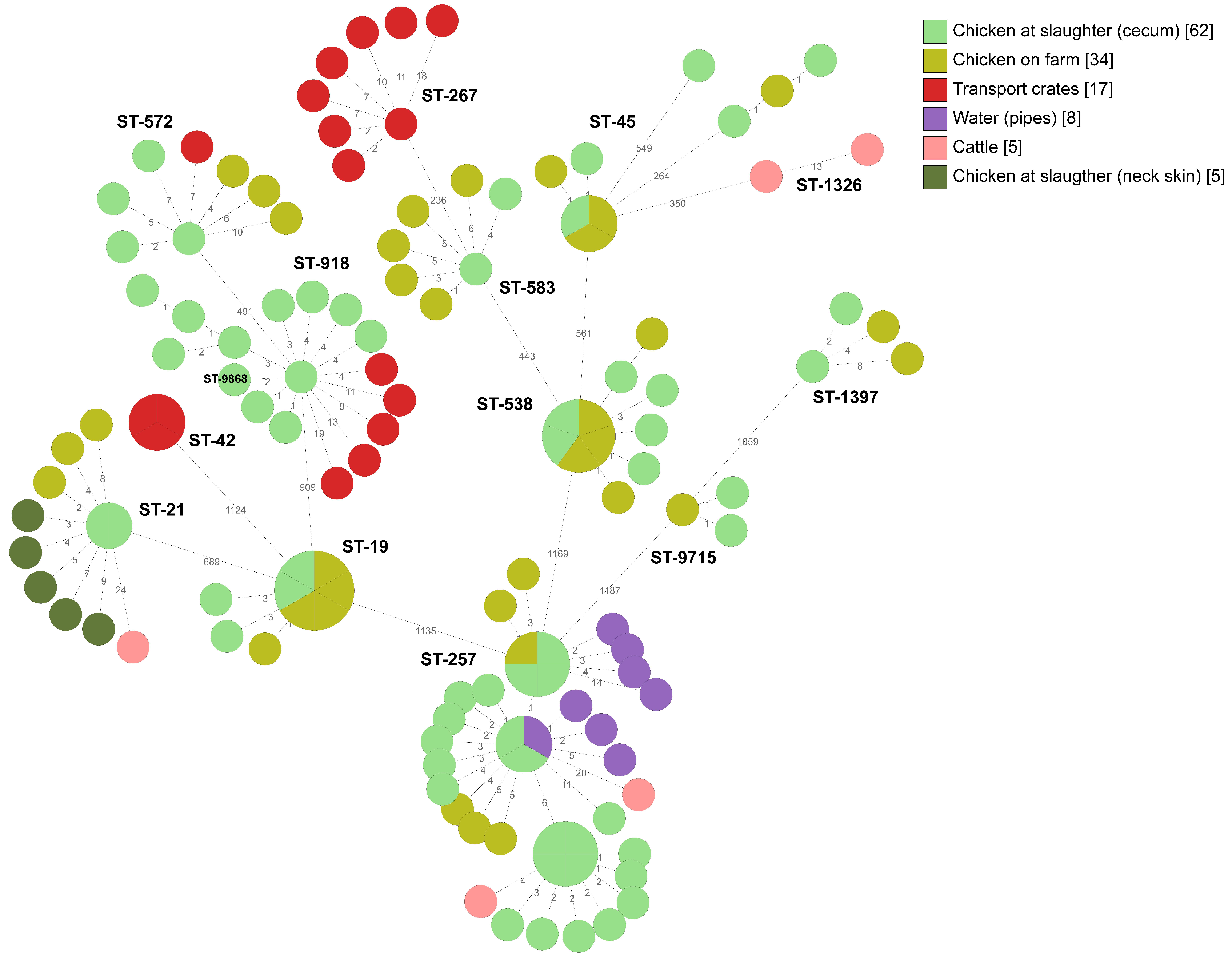
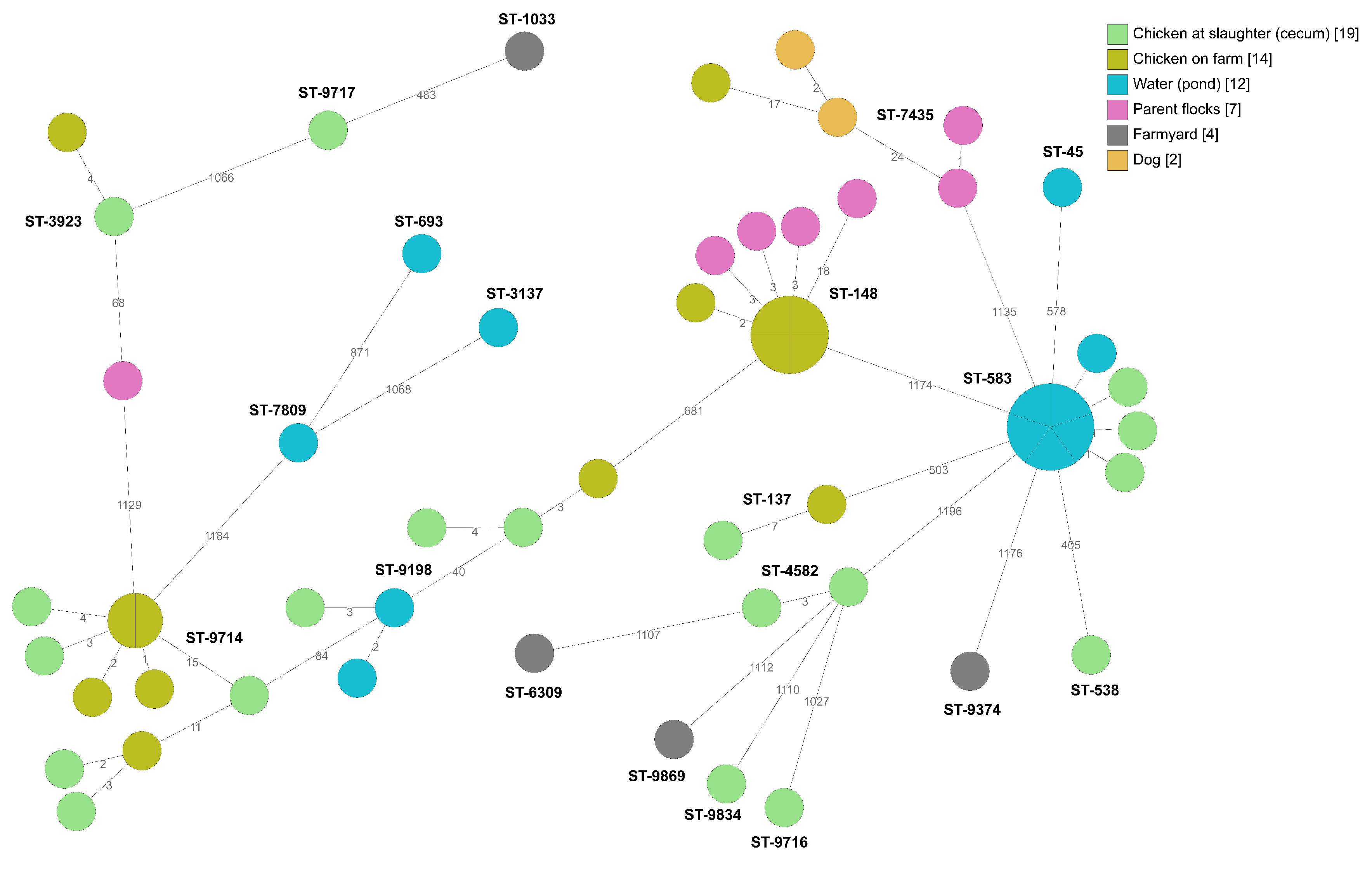
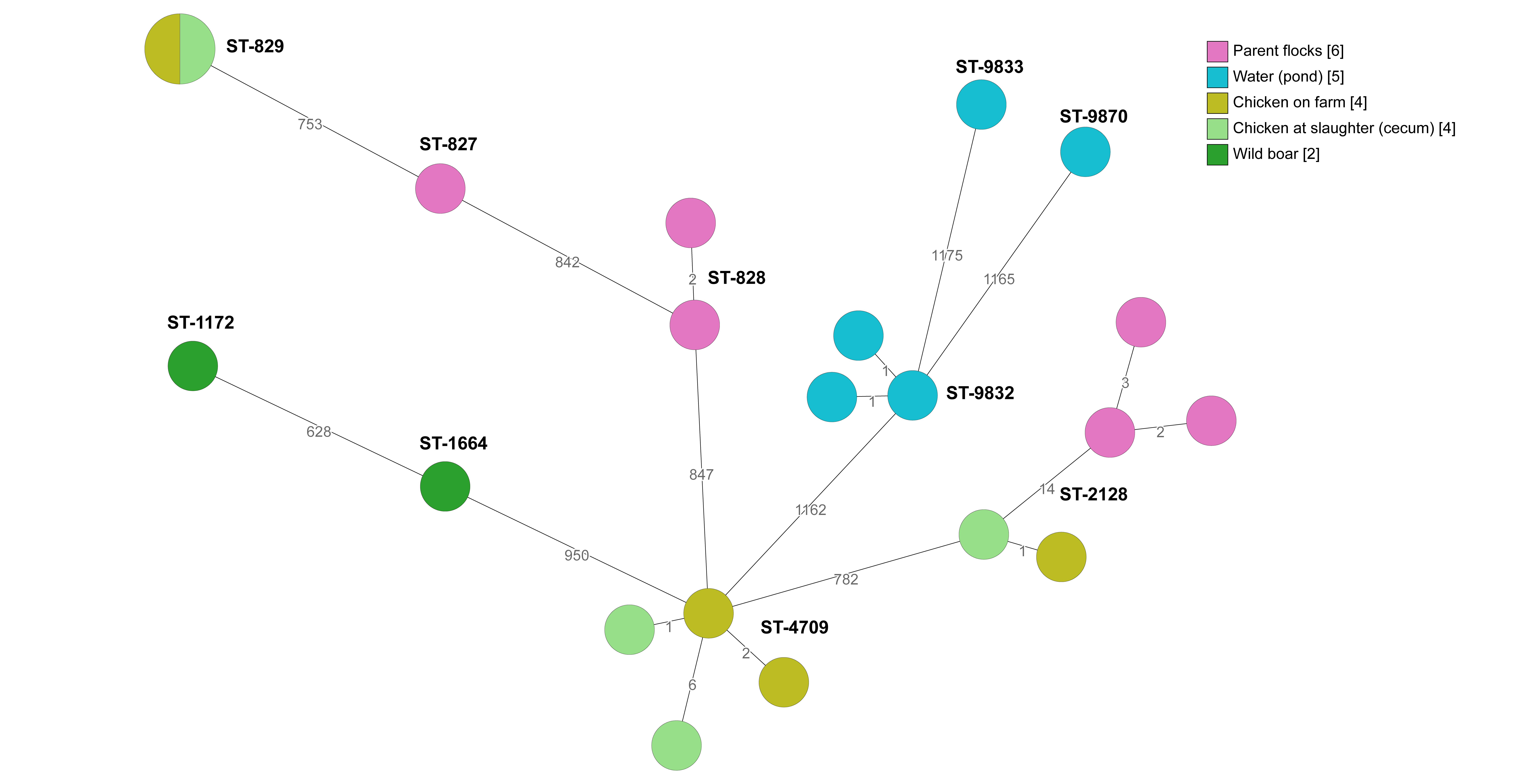
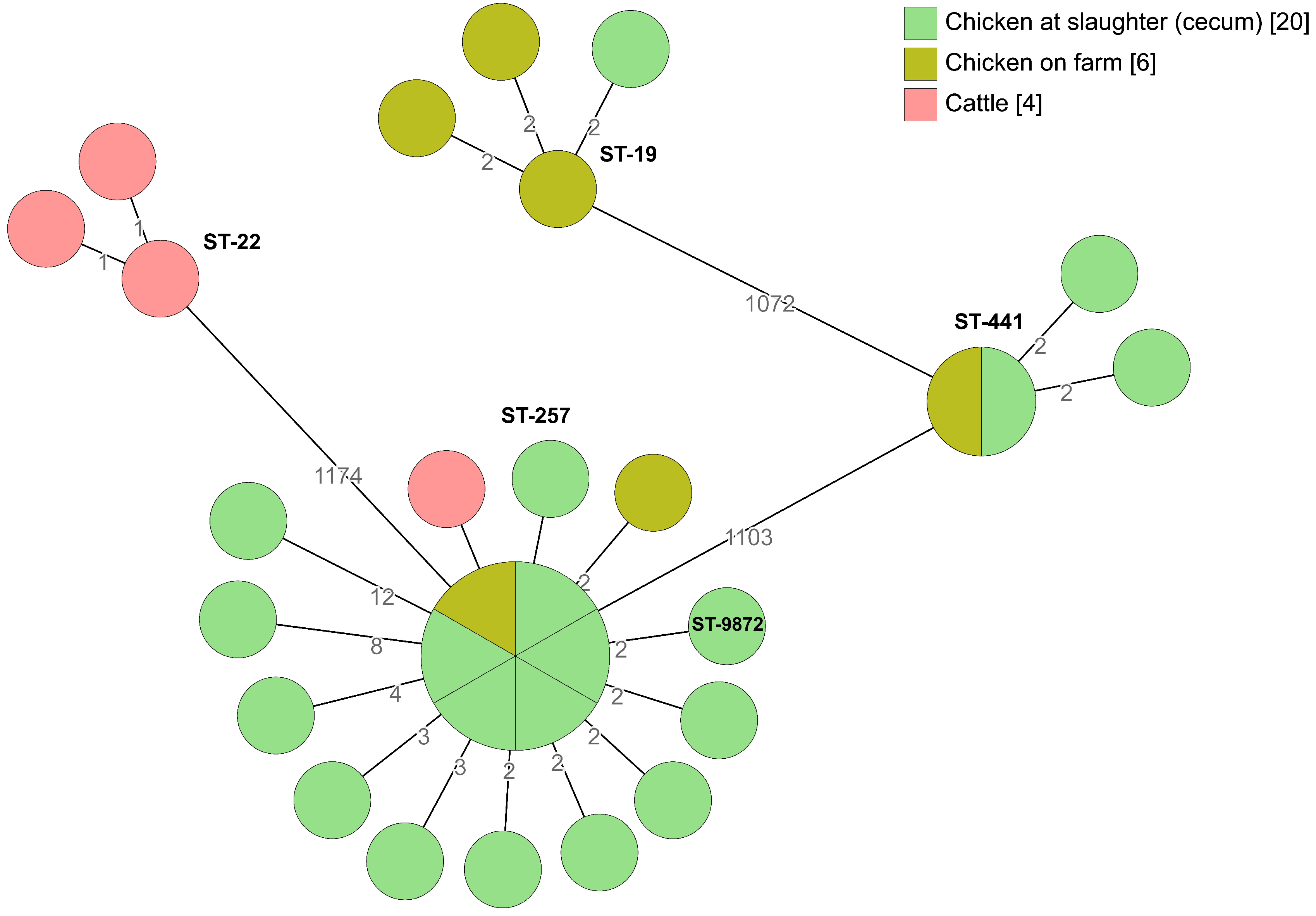
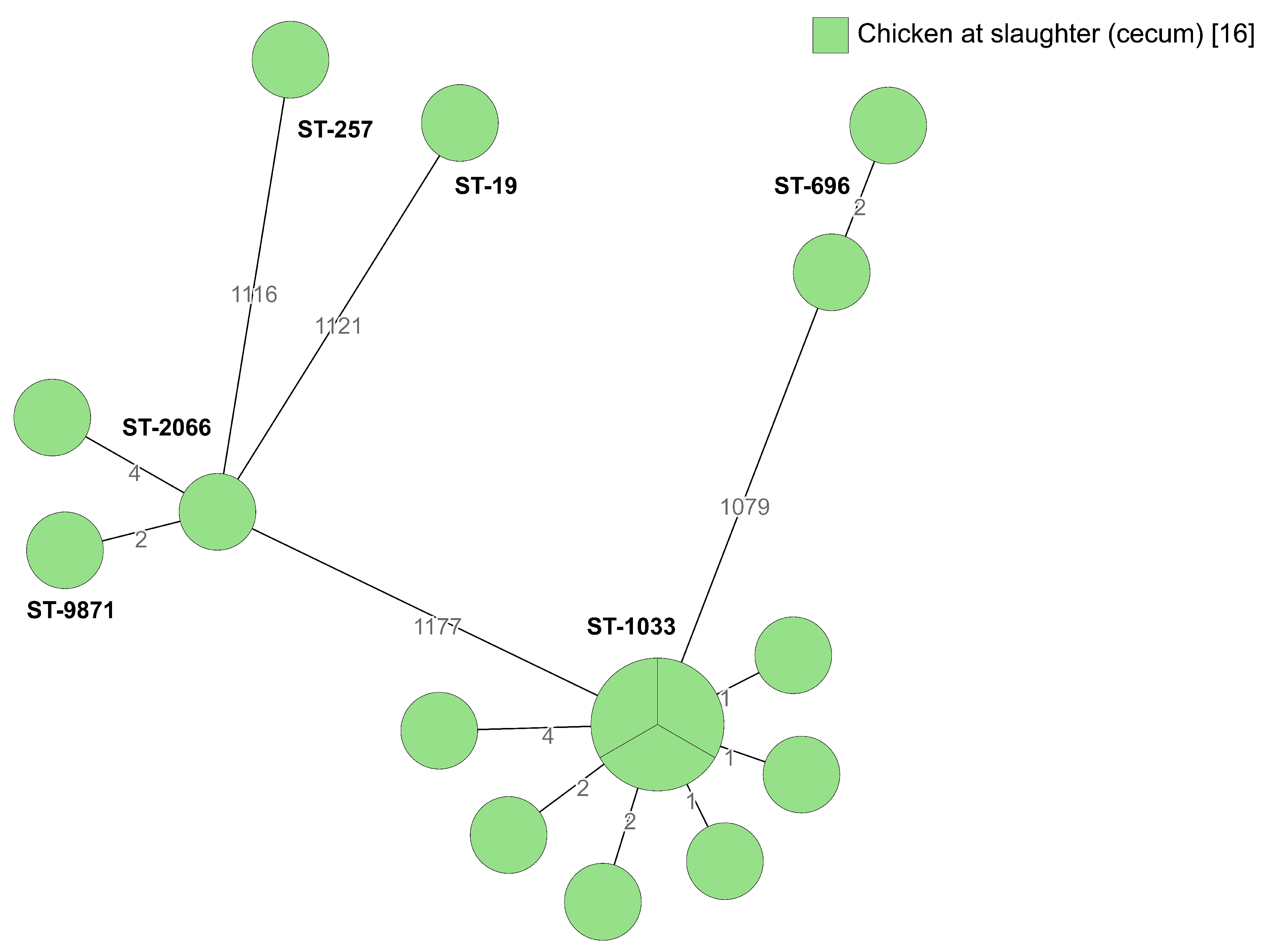
2.2. Whole-Genome Sequencing
2.2.1. Broiler Producer A
2.2.2. Broiler Producer B
2.2.3. Broiler Producer C
2.2.4. Broiler Producer D
3. Discussion
4. Materials and Methods
4.1. Broiler Producers
4.2. Sampling
4.3. Bacteriological Analysis
4.4. Whole-Genome Sequencing
4.5. Statistical Analysis
4.6. On-Farm Measures
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority (EFSA). Scientific report on the European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, e05926. [Google Scholar] [CrossRef]
- Geissler, A.L.; Bustos Carrillo, F.; Swanson, K.; Patrick, M.E.; Fullerton, K.E.; Bennett, C.; Barrett, K.; Mahon, B.E. Increasing Campylobacter Infections, Outbreaks, and Antimicrobial Resistance in the United States, 2004–2012. Clin. Infect. Dis. 2017, 65, 1624–1631. [Google Scholar] [CrossRef] [PubMed]
- Public Health Agency of Sweden. Tabellsamling över Anmälningspliktiga Smittsamma Sjukdomar I Sverige 2018; Public Health Agency of Sweden: Solna, Sweden, 2019. Available online: https://www.folkhalsomyndigheten.se/publicerat-material/publikationsarkiv/t/tabellsamling-over-anmalningspliktiga-smittsamma-sjukdomar-i-sverige-2018-epidemiologisk-arsrapport/ (accessed on 26 March 2020).
- Wagenaar, J.A.; French, N.P.; Havelaar, A.H. Preventing Campylobacter at the source: Why is it so difficult? Clin. Infect. Dis. 2013, 57, 1600–1606. [Google Scholar] [CrossRef] [PubMed]
- Sundström, K. Samhällskostnader för fem livsmedelsburna sjukdomar i Sverige. AgriFood Policy Brief. 2015, 5, 1–7. [Google Scholar]
- Zambrano, L.D.; Levy, K.; Menezes, N.P.; Freeman, M.C. Human diarrhea infections associated with domestic animal husbandry: A systematic review and meta-analysis. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Black, R.E.; Levine, M.M.; Clements, M.L.; Hughes, T.P.; Blaser, M.J. Experimental Campylobacter jejuni infection in humans. J. Infect. Dis. 1988, 157, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.A. Infective dose of Campylobacter jejuni in milk. Br. Med. J. (Clin. Res. Ed.) 1981, 282, 1584. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castano-Rodriguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef]
- Hansson, I.; Pudas, N.; Harbom, B.; Engvall, E.O. Within-flock variations of Campylobacter loads in caeca and on carcasses from broilers. Int. J. Food Microbiol. 2010, 141, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Rosenquist, H.; Sommer, H.M.; Nielsen, N.L.; Christensen, B.B. The effect of slaughter operations on the contamination of chicken carcasses with thermotolerant Campylobacter. Int. J. Food Microbiol. 2006, 108, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Stern, N.J.; Robach, M.C. Enumeration of Campylobacter spp. in broiler feces and in corresponding processed carcasses. J. Food Prot. 2003, 66, 1557–1563. [Google Scholar] [CrossRef]
- Hansson, I.; Gustafsson, P.; Lahti, E.; Olsson Engvall, E. 25 Years of the Swedish Campylobacter monitoring program. In Proceedings of the 18th International Workshop on Campylobacter, Helicobacter and Related Organisms, Rotorua, New Zealand, 1–5 November 2015. [Google Scholar]
- Bouwknegt, M.; van de Giessen, A.W.; Dam-Deisz, W.D.; Havelaar, A.H.; Nagelkerke, N.J.; Henken, A.M. Risk factors for the presence of Campylobacter spp. in Dutch broiler flocks. Prev. Vet. Med. 2004, 62, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Ellis-Iversen, J.; Jorgensen, F.; Bull, S.; Powell, L.; Cook, A.J.; Humphrey, T.J. Risk factors for Campylobacter colonisation during rearing of broiler flocks in Great Britain. Prev. Vet. Med. 2009, 89, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Hansson, I.; Forshell, L.P.; Gustafsson, P.; Boqvist, S.; Lindblad, J.; Engvall, E.O.; Andersson, Y.; Vågsholm, I. Summary of the Swedish Campylobacter program in broilers, 2001 through 2005. J. Food Prot. 2007, 70, 2008–2014. [Google Scholar] [CrossRef] [PubMed]
- Hansson, I.; Sandberg, M.; Habib, I.; Lowman, R.; Engvall, E.O. Knowledge gaps in control of Campylobacter for prevention of campylobacteriosis. Transbound Emerg. Dis. 2018, 65, 30–48. [Google Scholar] [CrossRef]
- Hansson, I.; Engvall, E.O.; Vågsholm, I.; Nyman, A. Risk factors associated with the presence of Campylobacter-positive broiler flocks in Sweden. Prev. Vet. Med. 2010, 96, 114–121. [Google Scholar] [CrossRef]
- Mughini Gras, L.; Smid, J.H.; Wagenaar, J.A.; de Boer, A.G.; Havelaar, A.H.; Friesema, I.H.; French, N.P.; Busani, L.; van Pelt, W. Risk factors for campylobacteriosis of chicken, ruminant, and environmental origin: A combined case-control and source attribution analysis. PLoS ONE 2012, 7, e42599. [Google Scholar] [CrossRef]
- Boysen, L.; Rosenquist, H.; Larsson, J.T.; Nielsen, E.M.; Sorensen, G.; Nordentoft, S.; Hald, T. Source attribution of human campylobacteriosis in Denmark. Epidemiol. Infect. 2014, 142, 1599–1608. [Google Scholar] [CrossRef]
- Thépault, A.; Poezevara, T.; Quesne, S.; Rose, V.; Chemaly, M.; Rivoal, K. Prevalence of Thermophilic Campylobacter in Cattle Production at Slaughterhouse Level in France and Link Between C. jejuni Bovine Strains and Campylobacteriosis. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Sondergaard, M.S.; Josefsen, M.H.; Lofstrom, C.; Christensen, L.S.; Wieczorek, K.; Osek, J.; Hoorfar, J. Low-cost monitoring of Campylobacter in poultry houses by air sampling and quantitative PCR. J. Food Prot. 2014, 77, 325–330. [Google Scholar] [CrossRef]
- Bahrndorff, S.; Rangstrup-Christensen, L.; Nordentoft, S.; Hald, B. Foodborne disease prevention and broiler chickens with reduced Campylobacter infection. Emerg. Infect. Dis. 2013, 19, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Allen, V.M.; Burton, C.H.; Wilkinson, D.J.; Whyte, R.T.; Harris, J.A.; Howell, M.; Tinker, D.B. Evaluation of the performance of different cleaning treatments in reducing microbial contamination of poultry transport crates. Br. Poult. Sci. 2008, 49, 233–240. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Atterbury, R.J.; Gigante, A.M.; Tinker, D.; Howell, M.; Allen, V.M. An improved cleaning system to reduce microbial contamination of poultry transport crates in the United Kingdom. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Callicott, K.A.; Friethriksdottir, V.; Reiersen, J.; Lowman, R.; Bisaillon, J.R.; Gunnarsson, E.; Berndtson, E.; Hiett, K.L.; Needleman, D.S.; Stern, N.J. Lack of evidence for vertical transmission of Campylobacter spp. in chickens. Appl. Environ. Microbiol. 2006, 72, 5794–5798. [Google Scholar] [CrossRef] [PubMed]
- Public Health Agency of Sweden. Campylobacter (Sverige nov 2018-jan 2019). Available online: https://www.folkhalsomyndigheten.se/smittskydd-beredskap/utbrott/utbrottsarkiv/campylobacter-sverige-2018-/ (accessed on 26 March 2020).
- Jacobs-Reitsma, W.F. Campylobacter bacteria in breeder flocks. Avian. Dis. 1995, 39, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Cox, N.A.; Richardson, L.J.; Maurer, J.J.; Berrang, M.E.; Fedorka-Cray, P.J.; Buhr, R.J.; Byrd, J.A.; Lee, M.D.; Hofacre, C.L.; O’Kane, P.M.; et al. Evidence for horizontal and vertical transmission in Campylobacter passage from hen to her progeny. J. Food Protect. 2012, 75, 1896–1902. [Google Scholar] [CrossRef]
- Näther, G.; Alter, T.; Martin, A.; Ellerbroek, L. Analysis of risk factors for Campylobacter species infection in broiler flocks. Poult. Sci. 2009, 88, 1299–1305. [Google Scholar] [CrossRef]
- The Institute of Environmental Science and Research Ltd. Notifiable Diseases in New Zealand: Annual Report Porirua, New Zealand. 2015. Available online: https://surv.esr.cri.nz/PDF_surveillance/AnnualRpt/AnnualSurv/2015/2015AnnualReportFinal.pdf (accessed on 26 March 2020).
- Nilsson, A.; Johansson, C.; Skarp, A.; Kaden, R.; Bertilsson, S.; Rautelin, H. Survival of Campylobacter jejuni and Campylobacter coli water isolates in lake and well water. APMIS 2018, 126, 762–770. [Google Scholar] [CrossRef]
- Trigui, H.; Thibodeau, A.; Fravalo, P.; Letellier, A.; Faucher, S.P. Survival in water of Campylobacter jejuni strains isolated from the slaughterhouse. Springerplus 2015, 4, 799. [Google Scholar] [CrossRef][Green Version]
- Public Health Agency of Sweden. Epidemiologisk Typning av Campylobacterisolat Insamlade Vecka 11 2017 (Epidemiological Typing of Campylobacter Isolates Collected Week 11 during 2017); Public Health Agency of Sweden: Solna, Sweden, 2017; pp. 1–10. Available online: https://www.folkhalsomyndigheten.se/contentassets/bc4b2a860df24cec84568829414f6cd6/typningsresultat-campylobacter-vecka-11-2017.pdf (accessed on 26 March 2020)(In Swedish, with English summary).
- Public Health Agency of Sweden. Epidemiologisk Typning av Campylobacterisolat Insamlade Vecka 34 2017 (Epidemiological Typing of Campylobacter Isolates Collected Week 34 during 2017); Public Health Agency of Sweden: Solna, Sweden, 2017; pp. 1–13. Available online: https://www.folkhalsomyndigheten.se/contentassets/bc4b2a860df24cec84568829414f6cd6/campylobacter-typningsrapport-v-34.pdf (accessed on 26 March 2020)(In Swedish, with English summary).
- Dryselius, R.; Jernberg, C.; Swedish Food Agency & Public Health Agency of Sweden. Campylobacter från Butik och Klinik (Campylobacter from Store and Clinic); Swedish Food Agency: Uppsala, Sweden, 2019; pp. 1–24. Available online: https://www.livsmedelsverket.se/globalassets/publikationsdatabas/rapporter/2019/s-2019-nr-01-campylobacter-fran-butik-och-klinik-livsmedelsverkets-rapportserie-s.pdf?AspxAutoDetectCookieSupport=1 (accessed on 26 March 2020)(In Swedish, with English summary).
- Joensen, K.G.; Kiil, K.; Gantzhorn, M.R.; Nauerby, B.; Engberg, J.; Holt, H.M.; Nielsen, H.L.; Petersen, A.M.; Kuhn, K.G.; Sando, G.; et al. Whole-Genome Sequencing to Detect Numerous Campylobacter jejuni Outbreaks and Match Patient Isolates to Sources, Denmark, 2015–2017. Emerg. Infect. Dis. 2020, 26, 523–532. [Google Scholar] [CrossRef]
- Public Health Agency of Sweden. Epidemiologisk Typning Av Campylobacterisolat Insamlade Vecka 11 2018 ((Epidemiological Typing of Campylobacter Isolates Collected Week 11 during 2018); Public Health Agency of Sweden: Solna, Sweden, 2018; pp. 1–10. Available online: https://www.folkhalsomyndigheten.se/contentassets/bc4b2a860df24cec84568829414f6cd6/epidemiologisk-typning-campylobacterisolat-insamlade-vecka-11-2018-rev-16-juli-2018.pdf (accessed on 26 March 2020)(In Swedish, with English summary).
- Battersby, T.; Walsh, D.; Whyte, P.; Bolton, D. Evaluating and improving terminal hygiene practices on broiler farms to prevent Campylobacter cross-contamination between flocks. Food Microbiol. 2017, 64, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Frosth, S.; Svensson, L.; Fernström, L.L.; Skarin, H.; Hansson, I. Detection of Campylobacter spp. in water by dead-end ultrafiltration and application at farm level. J. Appl. Microbiol. 2019, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- The Swedish Poultry Meat Association. All-time low prevalence of Campylobacter. Available online: https://svenskfagel.se/all-time-low-prevalence-of-campylobacter/ (accessed on 26 March 2020).
- Public Health Agency of Sweden. Campylobacterinfektion. Available online: https://www.folkhalsomyndigheten.se/folkhalsorapportering-statistik/statistik-a-o/sjukdomsstatistik/campylobacterinfektion/?t=county (accessed on 26 March 2020).
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Dingle, K.E.; Colles, F.M.; Wareing, D.R.; Ure, R.; Fox, A.J.; Bolton, F.E.; Bootsma, H.J.; Willems, R.J.; Urwin, R.; Maiden, M.C. Multilocus sequence typing system for Campylobacter jejuni. J. Clin. Microbiol. 2001, 39, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Silva, M.; Machado, M.P.; Silva, D.N.; Rossi, M.; Moran-Gilad, J.; Santos, S.; Ramirez, M.; Carrico, J.A. chewBBACA: A complete suite for gene-by-gene schema creation and strain identification. Microb. Genom. 2018, 4, e000166. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carrico, J.A.; Achtman, M. GrapeTree: Visualization of core genomic relationships among 100,000 bacterial pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Type | No. of Samples | C. jejuni | C. coli | C. lari | C. lanienae |
|---|---|---|---|---|---|
| Internal environment | |||||
| Anteroom | 19 | ||||
| Chicken on farm | 387 | 54 | 3 | ||
| Insects (flies) | 4 | ||||
| Ventilation vent | 2 | ||||
| Water | 22 | 2 | |||
| External environment | |||||
| Cattle | 16 | 9 | 3 | ||
| Chicken at slaughter (neck skin) | 5 | 5 | |||
| Dog | 2 | 2 | |||
| Fallow deer | 1 | ||||
| Farmyard | 23 | 4 | |||
| Mouse | 5 | ||||
| Parent flocks | 35 | 7 | 6 | 3 | |
| Snail | 2 | ||||
| Transport crates | 56 | 17 | |||
| Water | 18 | 8 | 1 | ||
| Wild birds | 18 | 1 | |||
| Wild boar | 11 | 4 | 4 | ||
| Total no. of samples | 626 | 108 | 14 | 7 | 4 |
| Clonal Complex | MLST | No. of Isolates | Broiler Producer | Source Origin |
|---|---|---|---|---|
| 21 | 19 | 14 | A, C, D | Chicken on farm, chicken at slaughter |
| 21 | 21 | 11 | A | Cattle, chicken on farm, chicken at slaughter (caecum and neck skin samples) |
| 22 | 22 | 3 | C | Cattle |
| 42 | 42 | 3 | A | Transport crates |
| 45 | 45 | 10 | A, B | Chicken on farm, chicken at slaughter, water (pond) |
| 45 | 137 | 2 | B | Chicken on farm, chicken at slaughter |
| 21 | 148 | 9 | B | Chicken on farm, parent flocks |
| 257 | 257 | 58 | A, C, D | Cattle, chicken on farm, chicken at slaughter, water (pipes) |
| 283 | 267 | 8 | A | Transport crates |
| NA | 441 | 4 | C | Chicken on farm, chicken at slaughter |
| 45 | 538 | 12 | A, B | Chicken on farm, chicken at slaughter |
| 206 | 572 | 8 | A | Chicken on farm, chicken at slaughter, transport crates |
| 45 | 583 | 16 | A, B | Chicken on farm, chicken at slaughter, water (pond) |
| NA | 693 | 1 | B | Water (pond) |
| 1332 | 696 | 2 | D | Chicken at slaughter |
| 828 | 827 | 1 | B | Parent flocks |
| 828 | 828 | 2 | B | Parent flocks |
| 828 | 829 | 2 | B | Chicken on farm, chicken at slaughter |
| 48 | 918 | 16 | A | Chicken at slaughter, transport crates |
| 1034 | 1033 | 10 | B, D | Farmyard, chicken at slaughter |
| NA | 1172 | 1 | B | Wild boar |
| 45 | 1326 | 2 | A | Cattle |
| NA | 1397 | 4 | A | Chicken on farm, chicken at slaughter |
| 828 | 1664 | 1 | B | Wild boar |
| 52 | 2066 | 2 | D | Chicken at slaughter |
| 828 | 2178 | 5 | B | Chicken on farm, chicken at slaughter, parent flocks |
| NA | 3137 | 1 | B | Water (pond) |
| NA | 3923 | 3 | B | Chicken on farm, chicken at slaughter, parent flocks |
| 952 | 4582 | 2 | B | Chicken at slaughter |
| 828 | 4709 | 4 | B | Chicken on farm, chicken at slaughter |
| NA | 6309 | 1 | B | Farmyard |
| NA | 7435 | 5 | B | Chicken on farm, dog, parent flocks |
| 1034 | 7809 | 1 | B | Water (pond) |
| 21 | 9198 | 6 | B | Chicken on farm, chicken at slaughter, water (pond) |
| NA | 9374 | 1 | B | Farmyard |
| 21 | 9714 * | 10 | B | Chicken on farm, chicken at slaughter |
| 1034 | 9715 * | 3 | A | Chicken on farm, chicken at slaughter |
| 952 | 9716 * | 1 | B | Chicken at slaughter |
| 1034 | 9717 * | 1 | B | Chicken at slaughter |
| NA | 9832 * | 3 | B | Water (pond) |
| NA | 9833 * | 1 | B | Water (pond) |
| NA | 9834 * | 1 | B | Chicken at slaughter |
| 48 | 9868 * | 1 | A | Chicken at slaughter |
| NA | 9869 * | 1 | B | Farmyard |
| NA | 9870 * | 1 | B | Water (pond) |
| 52 | 9871 * | 1 | D | Chicken at slaughter |
| 257 | 9872 * | 1 | C | Chicken at slaughter |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frosth, S.; Karlsson-Lindsjö, O.; Niazi, A.; Fernström, L.-L.; Hansson, I. Identification of Transmission Routes of Campylobacter and On-Farm Measures to Reduce Campylobacter in Chicken. Pathogens 2020, 9, 363. https://doi.org/10.3390/pathogens9050363
Frosth S, Karlsson-Lindsjö O, Niazi A, Fernström L-L, Hansson I. Identification of Transmission Routes of Campylobacter and On-Farm Measures to Reduce Campylobacter in Chicken. Pathogens. 2020; 9(5):363. https://doi.org/10.3390/pathogens9050363
Chicago/Turabian StyleFrosth, Sara, Oskar Karlsson-Lindsjö, Adnan Niazi, Lise-Lotte Fernström, and Ingrid Hansson. 2020. "Identification of Transmission Routes of Campylobacter and On-Farm Measures to Reduce Campylobacter in Chicken" Pathogens 9, no. 5: 363. https://doi.org/10.3390/pathogens9050363
APA StyleFrosth, S., Karlsson-Lindsjö, O., Niazi, A., Fernström, L.-L., & Hansson, I. (2020). Identification of Transmission Routes of Campylobacter and On-Farm Measures to Reduce Campylobacter in Chicken. Pathogens, 9(5), 363. https://doi.org/10.3390/pathogens9050363