Myrrh Oil in Vitro Inhibitory Growth on Bovine and Equine Piroplasm Parasites and Babesia microti of Mice

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. In vitro Growth Inhibition Assay
2.2. Drug Combination Test
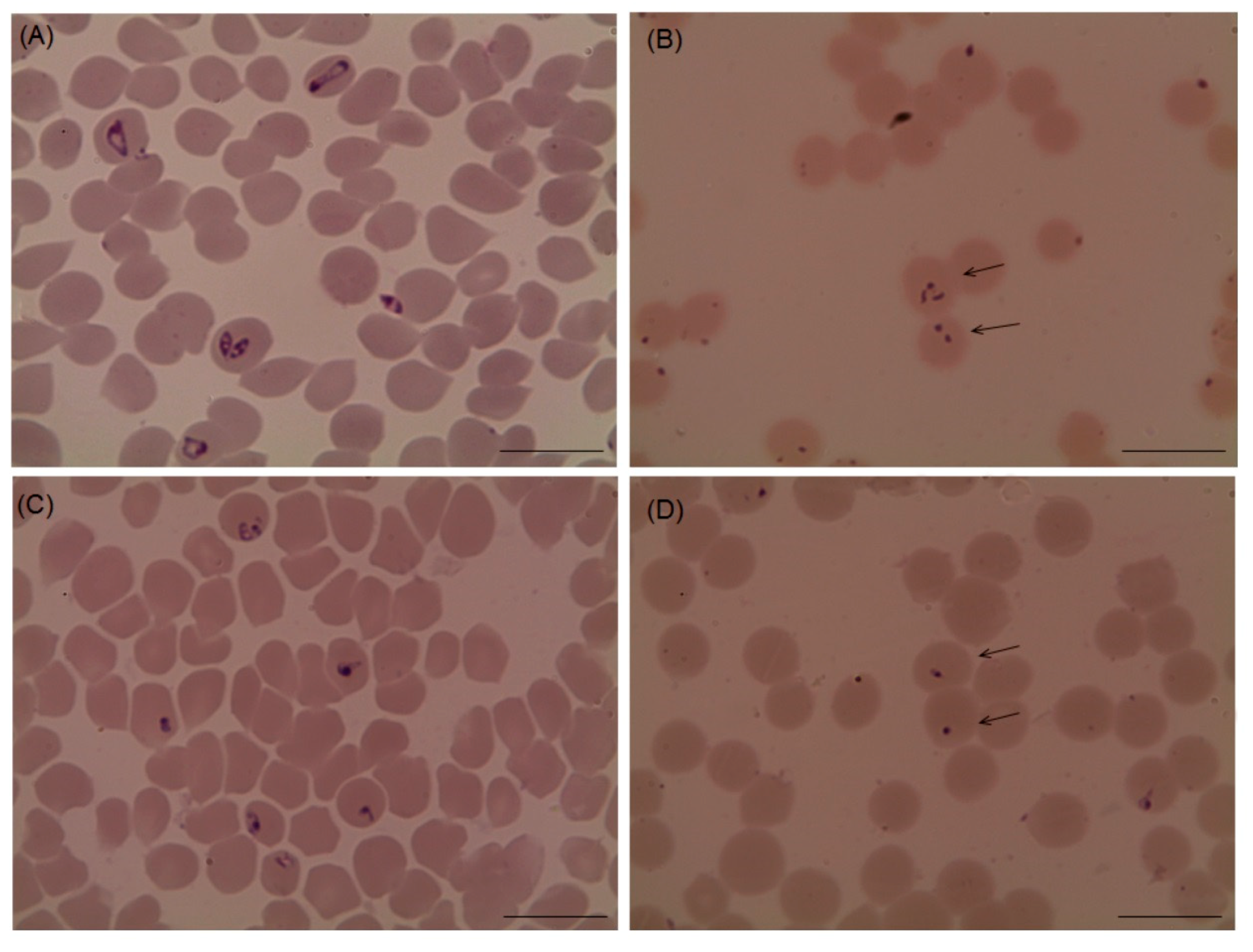
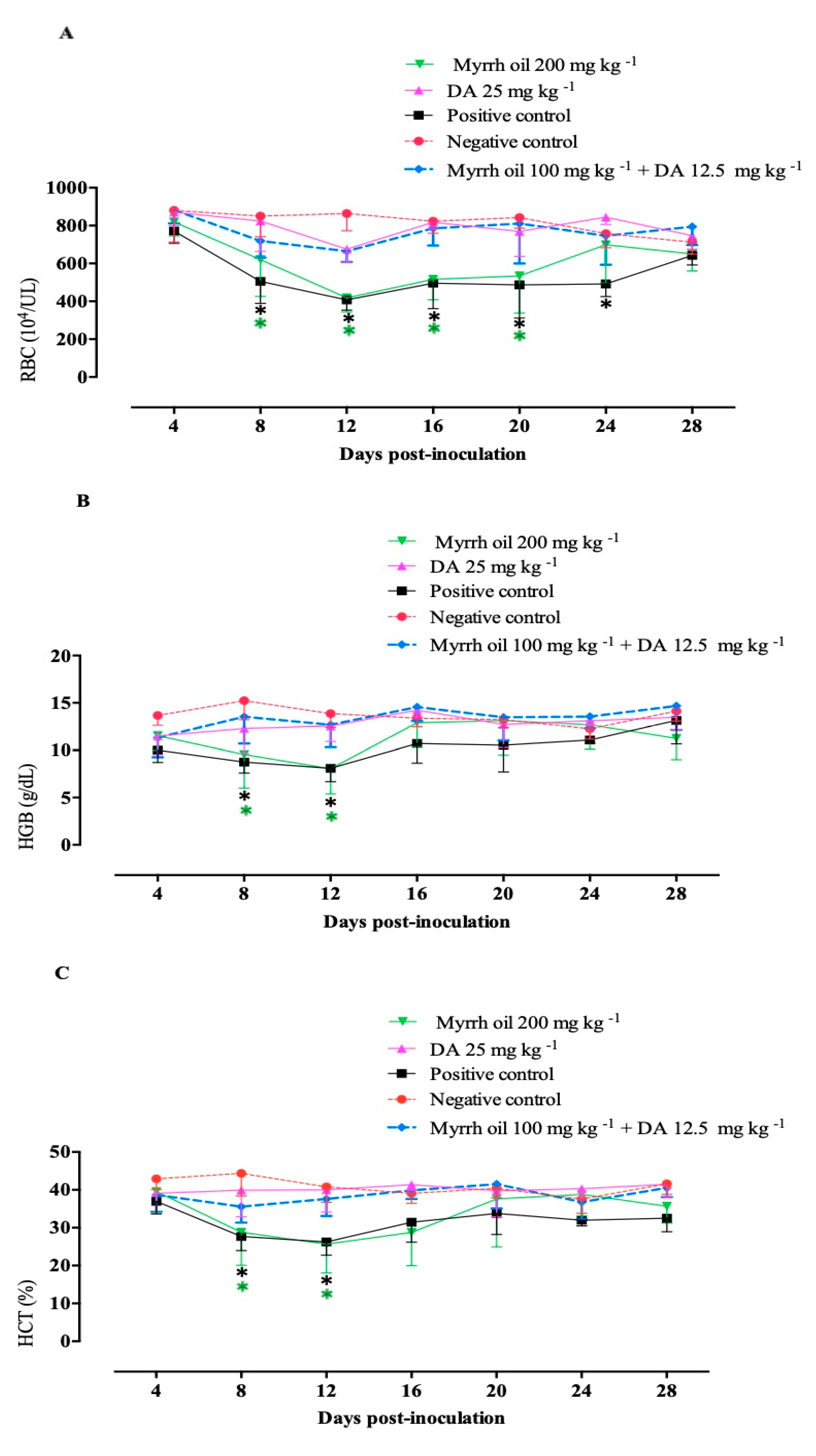
2.3. Chemotherapeutic Efficacy of Myrrh Oil on B. Microti Infection
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. In Vitro Growth Inhibition Assay and Viability Test
4.3. Myrrh Oil in Combination with Other Antibabesial Drugs in Vitro
4.4. Determination of Morphological Changes and the Toxic Effect of Myrrh Oil on Host Erythrocytes in Vitro
4.5. Chemotherapeutic Efficacy of Myrrh Oil on the Growth of B. Microti in Mice
4.6. The Potential of Myrrh Oil in the Recovery from the Anemia Accompanying Babesia
4.7. PCR Detection of B. Microti in Mice
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rizk, M.A.; El-Sayed, S.A.E.-S.; El-Khodery, S.; Yokoyama, N.; Igarashi, I. Discovering the in vitro potent inhibitors against Babesia and Theileria parasites by repurposing the Malaria Box: A review. Vet. Parasitol. 2019, 274, 108895. [Google Scholar] [CrossRef] [PubMed]
- Uilenberg, G. Babesia—A historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, S.A.E.-S.; Rizk, M.A.; Terkawi, M.A.; Yokoyama, N.; Igarashi, I. Molecular identification and antigenic characterization of Babesia divergens erythrocyte binding protein (BdEBP) as a potential vaccine candidate. Parasitol. Int. 2017, 66, 721–726. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, S.A.E.-S.; Rizk, M.A.; Terkawi, M.A.; Mousa, A.; Elsayed, G.; Fouda, M.; Yokoyama, N.; Igarashi, I. Cocktail of Theileria equi antigens for detecting infection in equines. Asian Pac. J. Trop. Biomed. 2015, 5, 977–981. [Google Scholar] [CrossRef]
- Vannier, E.; Gewurz, B.E.; Krause, P.J. Human babesiosis. Infect. Dis. Clin. N. Am. 2008, 22, 469–488. [Google Scholar] [CrossRef]
- Bakari, G.G.; Max, R.A.; Mdegela, R.H.; Phiri, E.C.; Mtambo, M.M. Effect of resinous extract from Commiphora swynnertonii (Burrt) on experimental coccidial infection in chickens. Trop. Anim. Health Prod. 2013, 45, 455–459. [Google Scholar] [CrossRef]
- Singh, R.; Joshi, V.; Gambhir, S. Anti-inflammatory activity of some traditional medicinal plants. Anc. Sci. Life 1998, 18, 160. [Google Scholar]
- Dolara, P.; Luceri, C.; Ghelardini, C.; Monserrat, C.; Aiolli, S.; Luceri, F.; Lodovici, M.; Menichetti, S.; Romanelli, M.N. Analgesic effects of myrrh. Nature 1996, 379, 29. [Google Scholar] [CrossRef]
- Massoud, A.; Ibrahim, S. Light microscopic study of the effect of new antischistosmal drug (myrrh extract) on the liver of mice. J. Egypt. Soc. Parasitol. 2005, 35, 971–988. [Google Scholar]
- Soliman, O.; El-Arman, M.; Abdul-Samie, E.; El-Nemr, H.; Massoud, A. Evaluation of myrrh (Mirazid) therapy in fascioliasis and intestinal schistosomiasis in children: Immunological and parasitological study. J. Egypt. Soc. Parasitol. 2004, 34, 941–966. [Google Scholar]
- Haridy, F.; Dawoud, H.; Morsy, T. Efficacy of Commiphora molmol (Mirazid) against sheep naturally infected with monieziasis expansa in Al-Santa Center, Gharbia Governorate, Egypt. J. Egypt. Soc. Parasitol. 2004, 34, 775–782. [Google Scholar]
- Al-Mathal, E.; Fouad, M. Myrrh (Commiphora molmol) in treatment of human and sheep dicrocoeliasis dendriticum in Saudi Arabia. J. Egypt. Soc. Parasitol. 2004, 34, 713–720. [Google Scholar] [PubMed]
- Massoud, A.; Kutkat, M.; Abdel, S.S.; El-Khateeb, R.M.; Labib, I.M. Acaricidal efficacy of Myrrh (Commiphora molmol) on the fowl tick Argas persicus (Acari: Argasidae). J. Egypt. Soc. Parasitol. 2005, 35, 667–686. [Google Scholar] [PubMed]
- Massoud, A.; Labib, I.M.; Rady, M. Biochemical changes of Culex pipiens larvae treated with oil and oleo-resin extracts of Myrrh Commiphora molmol. J. Egypt. Soc. Parasitol. 2001, 31, 517–529. [Google Scholar] [PubMed]
- Fathy, F. Effect of mirazid (Commiphora molmol) on experimental giardiasis. J. Egypt. Soc. Parasitol. 2011, 41, 155–177. [Google Scholar]
- Rizk, M.A.; El-Sayed, S.A.E.-S.; Terkawi, M.A.; Youssef, M.A.; El Said, E.S.E.S.; Elsayed, G.; El-Khodery, S.; El-Ashker, M.; Elsify, A.; Omar, M. Optimization of a fluorescence-based assay for large-scale drug screening against Babesia and Theileria parasites. PLoS ONE 2015, 10, e0125276. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, R.; Kamei, K.; Yamamura, M.; Nishiya, H.; Inouye, S.; Takahashi, M.; Abe, S. In vitro and in vivo anti-plasmodial activity of essential oils, including hinokitiol. Southeast Asian J. Trop. Med. Public Health 2012, 43, 270–279. [Google Scholar]
- Chen, Y.; Zhou, C.; Ge, Z.; Liu, Y.; Liu, Y.; Feng, W.; Li, S.; Chen, G.; Wei, T. Composition and potential anticancer activities of essential oils obtained from myrrh and frankincense. Oncol. Lett. 2013, 6, 1140–1146. [Google Scholar] [CrossRef]
- AbouLaila, M.; Nakamura, K.; Govind, Y.; Yokoyama, N.; Igarashi, I. Evaluation of the in vitro growth-inhibitory effect of epoxomicin on Babesia parasites. Vet. Parasitol. 2010, 167, 19–27. [Google Scholar] [CrossRef]
- Brockelman, C.R.; Tan-ariya, P. Development of an in vitro microtest to assess drug susceptibility of Babesia bovis and Babesia bigemina. J. Parasitol. 1991, 77, 994–997. [Google Scholar] [CrossRef]
- Rodriguez, R.; Trees, A. In vitro responsiveness of Babesia bovis to imidocarb dipropionate and the selection of a drug-adapted line. Vet. Parasitol. 1996, 62, 35–41. [Google Scholar] [CrossRef]
- Bork, S.; Yokoyama, N.; Ikehara, Y.; Kumar, S.; Sugimoto, C.; Igarashi, I. Growth-inhibitory effect of heparin on Babesia parasites. Antimicrob. Agents Chemother. 2004, 48, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, I.; Njonge, F.K.; Kaneko, Y.; Nakamura, Y. Babesia bigemina: In Vitro and in VivoEffects of Curdlan Sulfate on Growth of Parasites. Exp. Parasitol. 1998, 90, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Bork, S.; Yokoyama, N.; Matsuo, T.; Claveria, F.G.; Fujisaki, K.; Igarashi, I. Growth inhibitory effect of triclosan on equine and bovine Babesia parasites. Am. J. Trop. Med. Hyg. 2003, 68, 334–340. [Google Scholar] [CrossRef]
- Aboulaila, M.; Yokoyama, N.; Igarashi, I. Inhibitory effects of (-)-epigallocatechin-3-gallate from green tea on the growth of Babesia parasites. Parasitology 2010, 137, 785–791. [Google Scholar] [CrossRef]
- AbouLaila, M.; Sivakumar, T.; Yokoyama, N.; Igarashi, I. Inhibitory effect of terpene nerolidol on the growth of Babesia parasites. Parasitol. Int. 2010, 59, 278–282. [Google Scholar] [CrossRef]
- AbouLaila, M.; Munkhjargal, T.; Sivakumar, T.; Ueno, A.; Nakano, Y.; Yokoyama, M.; Yoshinari, T.; Nagano, D.; Katayama, K.; El-Bahy, N. Apicoplast-targeting antibacterials inhibit the growth of Babesia parasites. Antimicrob. Agents Chemother. 2012, 56, 3196–3206. [Google Scholar] [CrossRef]
- Salama, A.A.; AbouLaila, M.; Moussa, A.A.; Nayel, M.A.; El-Sify, A.; Terkawi, M.A.; Hassan, H.Y.; Yokoyama, N.; Igarashi, I. Evaluation of in vitro and in vivo inhibitory effects of fusidic acid on Babesia and Theileria parasites. Vet. Parasitol. 2013, 191, 1–10. [Google Scholar] [CrossRef]
- AbouLaila, M.; Batadoj, D.; Salama, A.; Munkhjargal, T.; Ichikawa-Seki, M.; Terkawi, M.A.; Yokoyama, N.; Igarashi, I. Evaluation of the inhibitory effects of miltefosine on the growth of Babesia and Theileria parasites. Vet. Parasitol. 2014, 204, 104–110. [Google Scholar] [CrossRef]
- Salama, A.A.; AbouLaila, M.; Terkawi, M.A.; Mousa, A.; El-Sify, A.; Allaam, M.; Zaghawa, A.; Yokoyama, N.; Igarashi, I. Inhibitory effect of allicin on the growth of Babesia and Theileria equi parasites. Parasitol. Res. 2014, 113, 275–283. [Google Scholar] [CrossRef]
- Rizk, M.A.; El-Sayed, S.A.E.-S.; AbouLaila, M.; Yokoyama, N.; Igarashi, I. Evaluation of the inhibitory effect of N-acetyl-L-cysteine on Babesia and Theileria parasites. Exp. Parasitol. 2017, 179, 43–48. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, S.A.E.-S.; Rizk, M.A.; Yokoyama, N.; Igarashi, I. Evaluation of the in vitro and in vivo inhibitory effect of thymoquinone on piroplasm parasites. Parasites Vectors 2019, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.A.; AbouLaila, M.; El-Sayed, S.A.E.-S.; Guswanto, A.; Yokoyama, N.; Igarashi, I. Inhibitory effects of fluoroquinolone antibiotics on Babesia divergens and Babesia microti, blood parasites of veterinary and zoonotic importance. Infect. Drug Resist. 2018, 11, 1605. [Google Scholar] [CrossRef] [PubMed]
- Aboulaila, M.R.; Rizk, M.A.; El-Sayed, S.; Yoko-Yama, N.; Igarashi, I. In vitro antiparasitic effects of six beverages on the growth of Babesia and Theileria parasites. Ann. Complement. Altern. Med. 2018, 3, 3–8. [Google Scholar]
- Ganchimeg, D.; Batbold, B.; Murata, T.; Davaapurev, B.-O.; Munkhjargal, T.; Tuvshintulga, B.; Suganuma, K.; Igarashi, I.; Buyankhishig, B.; Sasaki, K. Flavonoids isolated from the flowers of Pulsatilla flavescens and their anti-piroplasm activity. J. Nat. Med. 2019, 73, 633–640. [Google Scholar] [CrossRef]
- Guswanto, A.; Nugraha, A.B.; Tuvshintulga, B.; Tayebwa, D.S.; Rizk, M.A.; Batiha, G.E.-S.; Gantuya, S.; Sivakumar, T.; Yokoyama, N.; Igarashi, I. 17-DMAG inhibits the multiplication of several Babesia species and Theileria equi on in vitro cultures, and Babesia microti in mice. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 104–111. [Google Scholar] [CrossRef]
- Nugraha, A.B.; Tuvshintulga, B.; Guswanto, A.; Tayebwa, D.S.; Rizk, M.A.; Gantuya, S.; Batiha, G.E.-S.; Beshbishy, A.M.; Sivakumar, T.; Yokoyama, N. Screening the Medicines for Malaria Venture Pathogen Box against piroplasm parasites. Int. J. Parasitol. Drugs Drug Resist. 2019, 10, 84–90. [Google Scholar] [CrossRef]
- Rizk, M.A.; El-Sayed, S.A.E.-S.; AbouLaila, M.; Eltaysh, R.; Yokoyama, N.; Igarashi, I. Performance and consistency of a fluorescence-based high-throughput screening assay for use in Babesia drug screening in mice. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Brieskorn, C.H.; Noble, P. Drei neue furanogermacrene aus myrrhe. Tetrahedron Lett. 1980, 21, 1511–1514. [Google Scholar] [CrossRef]
- Maradufu, A.; Warthen Jr, J.D. Furanosesquiterpenoids from Commiphora myrrh oil. Plant Sci. 1988, 57, 181–184. [Google Scholar] [CrossRef]
- Rizk, M.A.; Ji, S.; Liu, M.; El-Sayed, S.A.E.-S.; Li, Y.; Byamukama, B.; Ringo, A.E.; Xuan, X.; Igarashi, I. Closing the empty anti-Babesia gibsoni drug pipeline in vitro using fluorescence-based high throughput screening assay. Parasitol. Int. 2020, 75, 102054. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.A.; El-Sayed, S.A.E.-S.; AbouLaila, M.; Tuvshintulga, B.; Yokoyama, N.; Igarashi, I. Large-scale drug screening against Babesia divergens parasite using a fluorescence-based high-throughput screening assay. Vet. Parasitol. 2016, 227, 93–97. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | IC50 (µg/mL) a | |
|---|---|---|
| Myrrh Oil | Diminazene Aceturate | |
| Babesiabovis | 1.04 ± 0.13 | 0.18 ± 0.03 |
| Babesiabigemina | 0.96 ± 0.03 | 0.09 ± 0.005 |
| Babesiadivergens | 7.70 ± 0.41 | 0.08 ± 0.01 |
| Theileria equi | 0.33 ± 0.10 | 0.35 ± 0.01 |
| Babesiacaballi | 0.18 ± 0.03 | 0.01 ± 0.001 |
| Plasmodium falciparumb | 100 | ND |
| HepG2 cells c | 39.2 | ND |
| Parasite | Drug Concentrations (μg/mL) a | |||||
|---|---|---|---|---|---|---|
| 0.1 | 0.5 | 1.0 | 2.0 | 5.0 | 10 | |
| B. bovis | + | + | + | − | − | − |
| B. bigemina | + | + | + | − | − | − |
| B. divergens | + | + | + | + | + | − |
| T. equi | + | + | + | + | + | − |
| B. caballi | + | + | + | + | + | − |
| Group | Fluorescence Values (mean ± SD) a | |
|---|---|---|
| B. divergens | B. bovis | |
| Control | 516.46 ± 42.51 | 220.44 ± 17.28 |
| DA IC50 | 240.01±21.86 | 121.96 ± 8.30 |
| M1 (0.75:0.50)b | 424.33 ± 23.72 * | 30.59 ± 12.69 ** |
| M2 (0.75:0.12) b | 446.65 ± 24.54 * | 67.23 ± 13.26 ** |
| M3 (0.50:0.50) b | 478.39 ± 35.17 | 67.07 ± 7.77 ** |
| M4 (0.50:0.12) b | 483.60 ± 38.57 | 95.56 ± 1.48 ** |
| M5 (0.25:0.50) b | 481.66 ± 42.51 | 106.02 ± 25.12 * |
| M6 (0.25:0.12) b | 499.17 ± 41.87 | 114.05 ± 5.74 * |
| M7 (0.12:0.50) b | 509.22 ± 33.15 | 120.75 ± 7.51 * |
| M8 (0.12:0.12) b | 513.33 ± 27.66 | 124.88 ± 10.39 * |
| M9 (0.06:0.50) b | 516.25 ± 21.08 | 131.02 ± 9.47 * |
| M10 (0.06:0.12) b | 516.10 ± 31.42 | 140.11 ± 11.08 * |
| Group | Fluorescence Values (mean ± SD) a | |
|---|---|---|
| B. divergens | B. bovis | |
| Control | 530.21 ± 14.83 | 230.44 ± 35.56 |
| PYR IC50 | 283.50 ± 20.11 | 127.31 ± 14.21 |
| M1 (0.75:0.50)b | 484.18 ± 6.04 * | 31.77 ± 4.95 ** |
| M2 (0.75:0.12) b | 496.21 ± 9.04 * | 37.39 ± 9.39 ** |
| M3 (0.50:0.50) b | 496.40 ± 10.79 * | 36.18 ± 8.26 ** |
| M4 (0.50:0.12) b | 511.88 ± 4.91 | 38.09 ± 7.68 ** |
| M5 (0.25:0.50) b | 512.41 ± 5.39 | 45.83 ± 11.18 ** |
| M6 (0.25:0.12) b | 514.30 ± 10.50 | 74.04 ± 16.82 ** |
| M7 (0.12:0.50) b | 516.90 ± 6.02 | 85.515 ± 14.53 ** |
| M8 (0.12:0.12) b | 520.18 ± 6.32 | 122.44 ± 24.81 * |
| M9 (0.06:0.50) b | 529.13 ± 4.90 | 130.81 ± 28.24 * |
| M10 (0.06:0.12) b | 534.72 ± 8.31 | 141.06 ± 19.66 * |
| Group | Fluorescence Values (mean ± SD) a | |
|---|---|---|
| B. divergens | B. bovis | |
| Control | 447.33 ± 37.50 | 222.94 ± 39.10 |
| Luteolin IC50 | 231.40 ± 16.02 | 119.35 ± 10.86 |
| M1 (0.75:0.50)b | 413.52 ± 23.38 | 2.57 ± 1.27 ** |
| M2 (0.75:0.12) b | 420.86 ± 28.36 | 8.95 ± 1.91 ** |
| M3 (0.50:0.50) b | 429.83 ± 27.96 | 22.76 ± 7.33 ** |
| M4 (0.50:0.12) b | 425.425 ± 27.55 | 31.58 ± 8.19 ** |
| M5 (0.25:0.50) b | 433.56 ± 29.78 | 100.89 ± 2.68 ** |
| M6 (0.25:0.12) b | 435.37 ± 28.84 | 122.34 ± 28.41 * |
| M7 (0.12:0.50) b | 435.43 ± 31.26 | 140.94 ± 19.29 * |
| M8 (0.12:0.12) b | 434.18 ± 25.39 | 178.99 ± 8.20 |
| M9 (0.06:0.50) b | 436.66 ± 27.88 | 185.54 ± 10.68 |
| M10 (0.06:0.12) b | 448.03 ± 15.92 | 191.33 ± 18.37 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
AbouLaila, M.; El-Sayed, S.A.E.-S.; Omar, M.A.; Al-Aboody, M.S.; Abdel Aziz, A.R.; Abdel-Daim, M.M.; Rizk, M.A.; Igarashi, I. Myrrh Oil in Vitro Inhibitory Growth on Bovine and Equine Piroplasm Parasites and Babesia microti of Mice. Pathogens 2020, 9, 173. https://doi.org/10.3390/pathogens9030173
AbouLaila M, El-Sayed SAE-S, Omar MA, Al-Aboody MS, Abdel Aziz AR, Abdel-Daim MM, Rizk MA, Igarashi I. Myrrh Oil in Vitro Inhibitory Growth on Bovine and Equine Piroplasm Parasites and Babesia microti of Mice. Pathogens. 2020; 9(3):173. https://doi.org/10.3390/pathogens9030173
Chicago/Turabian StyleAbouLaila, Mahmoud, Shimaa Abd El-Salam El-Sayed, Mosaab A. Omar, Mohammad Saleh Al-Aboody, Amer R. Abdel Aziz, Mohamed M. Abdel-Daim, Mohamed Abdo Rizk, and Ikuo Igarashi. 2020. "Myrrh Oil in Vitro Inhibitory Growth on Bovine and Equine Piroplasm Parasites and Babesia microti of Mice" Pathogens 9, no. 3: 173. https://doi.org/10.3390/pathogens9030173
APA StyleAbouLaila, M., El-Sayed, S. A. E.-S., Omar, M. A., Al-Aboody, M. S., Abdel Aziz, A. R., Abdel-Daim, M. M., Rizk, M. A., & Igarashi, I. (2020). Myrrh Oil in Vitro Inhibitory Growth on Bovine and Equine Piroplasm Parasites and Babesia microti of Mice. Pathogens, 9(3), 173. https://doi.org/10.3390/pathogens9030173