Characterization of Legionella pneumophila Populations by Multilocus Variable Number of Tandem Repeats (MLVA) Genotyping from Drinking Water and Biofilm in Hospitals from Different Regions of the West Bank

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites, Water and Biofilm Sampling
2.2. Cultivation-Dependent Analysis
2.3. Cultivation-Independent Analysis (16S rDNA PCR)
2.4. Sero-Grouping of Legionella Isolates
2.5. Genotyping of L. pneumophila Isolates
2.6. Statistical Analysis
3. Results
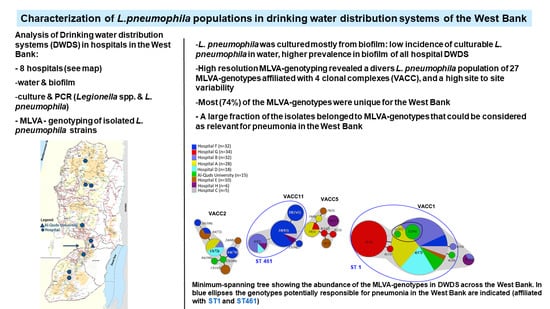
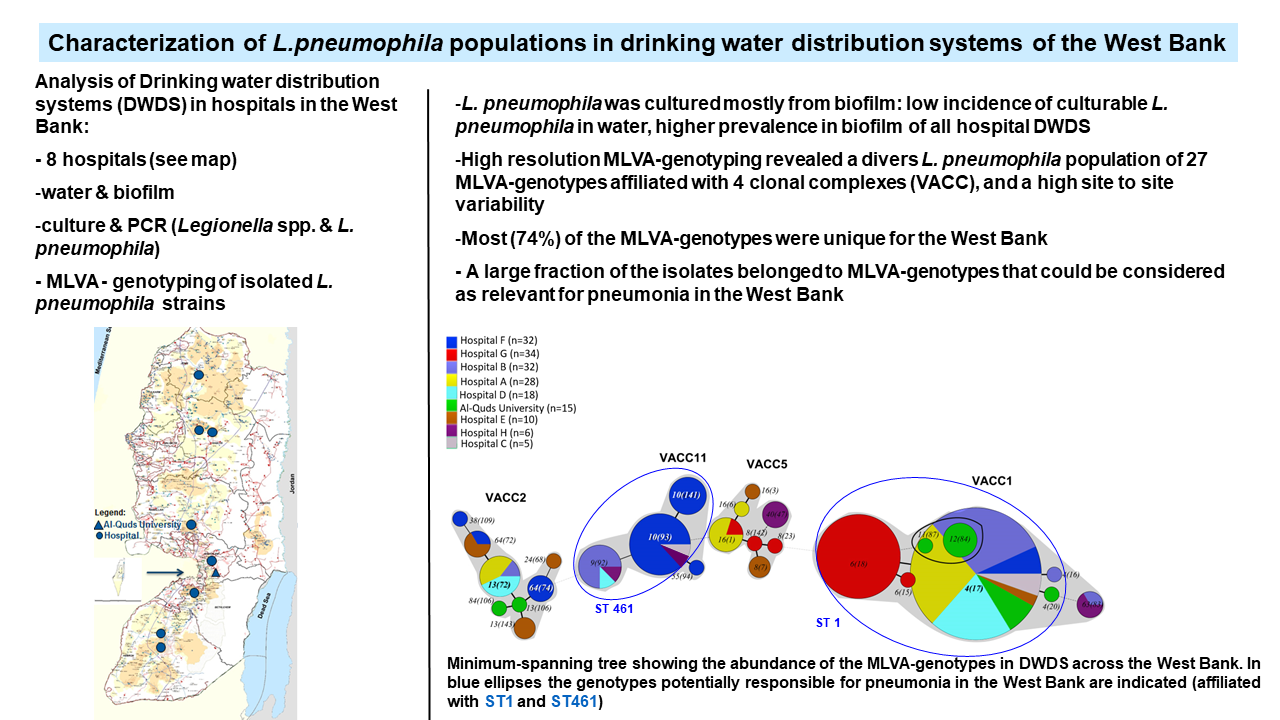
3.1. Biological and Physico-chemical Characteristics of Drinking Water Distribution Systems (DWDS)
3.2. Cultivation Dependent Analysis (CDA) Versus Cultivation Independent Analysis (CIA)
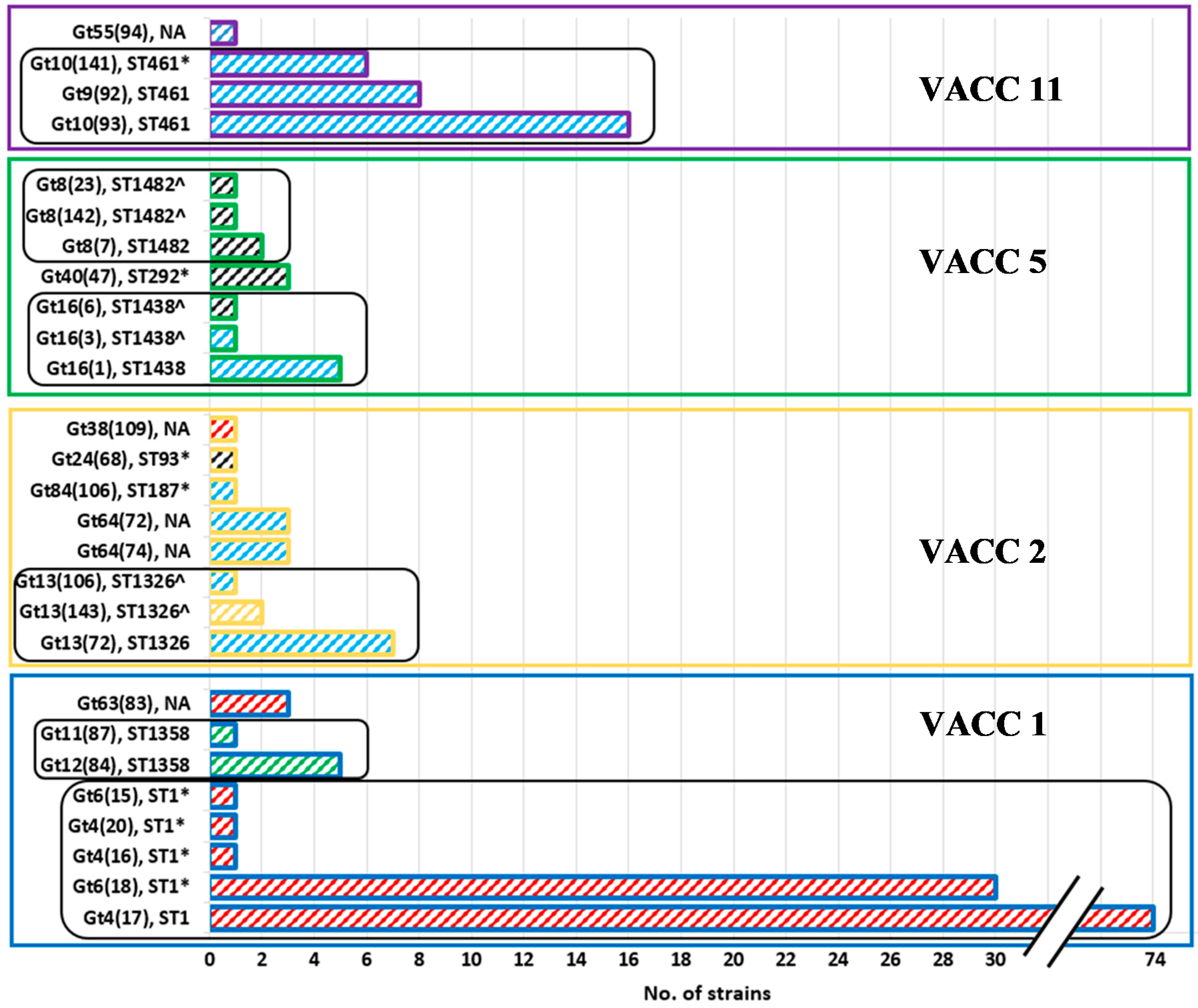
3.3. MLVA-8(12) Genotypes of L. pneumophila Isolates
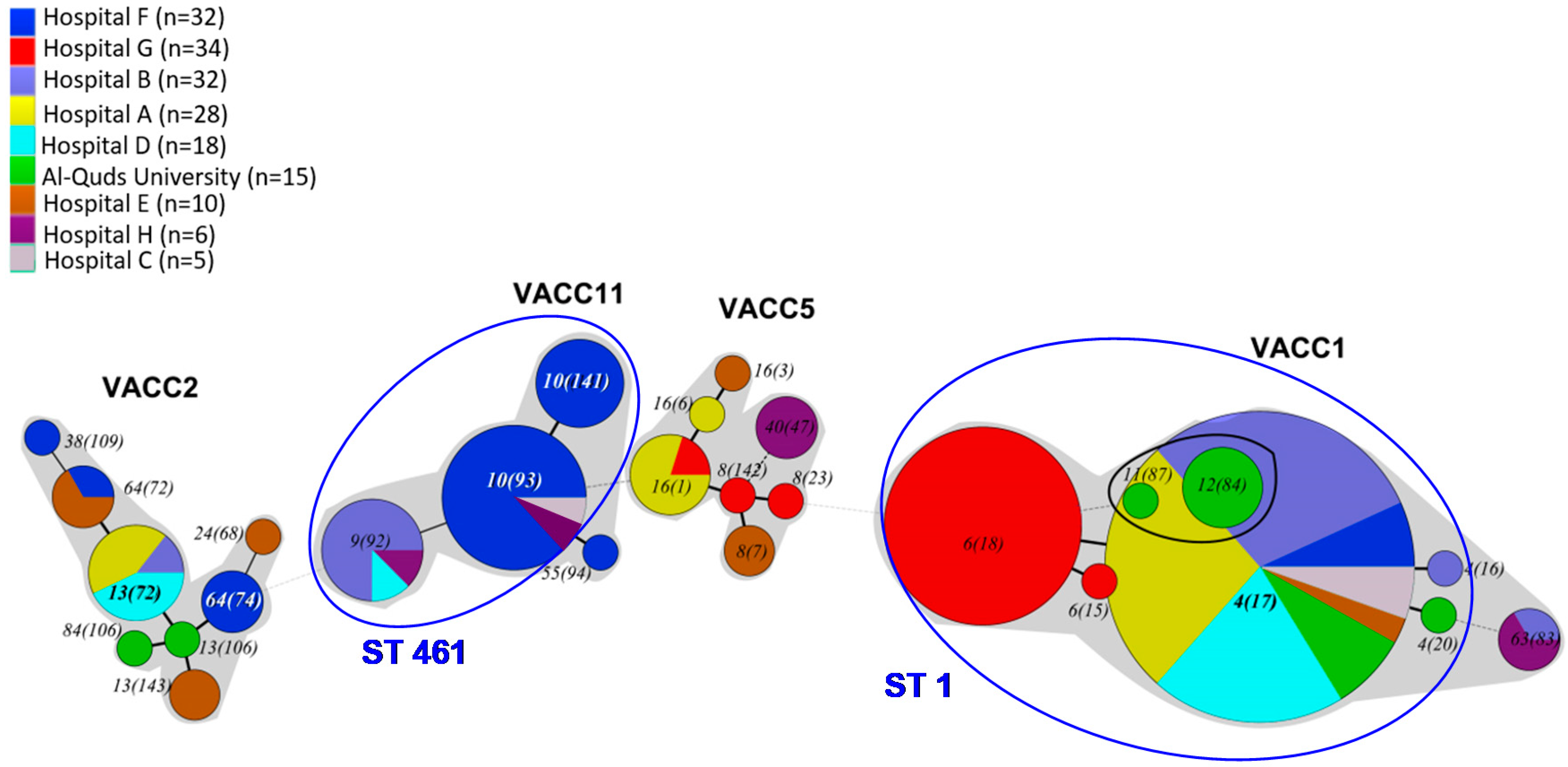
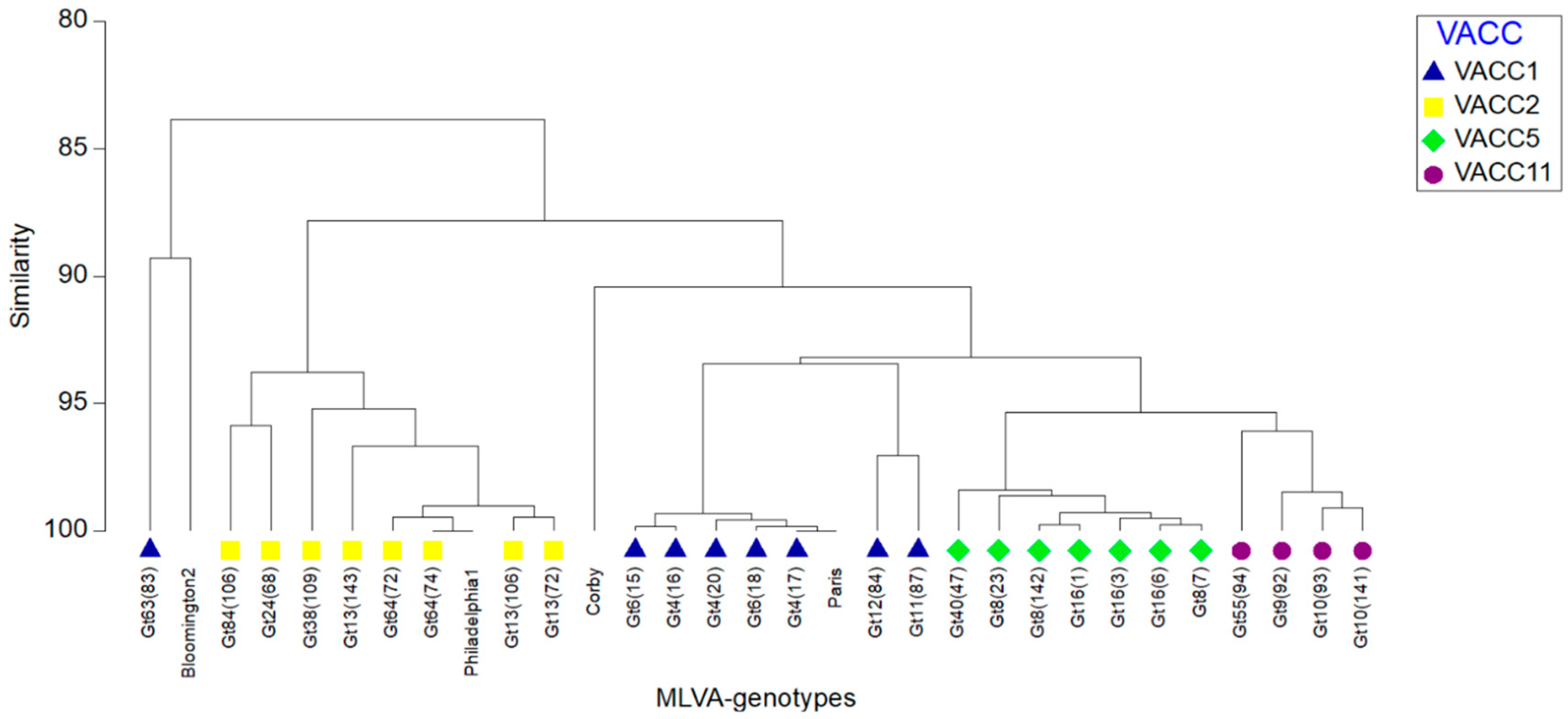
3.4. VNTR Clonal Complexes (VACC) and Relationship among the Genotypes
3.5. Diversity within the Clonal Complexes (VACC)
3.6. Comparison of MLVA-8(12) Genotypes and Clonal Complexes with Sequence Types (ST)
3.7. Serotype Distribution of L. pneumophila Isolates and Relationship with MLVA Genotypes and Clonal Complexes
3.8. Prevalence and Abundance of L. pneumophila MLVA-genotypes and Clonal Complexes (VACCs)
4. Discussion
4.1. Legionella Abundance in Hospital Water and Biofilm of the West Bank
4.2. General Health Relevant Aspects of the Isolated Strains
4.3. Genotyping Using MLVA—What Resolution Is Needed for Ecological and Clinical Issues?
4.4. Comparison of L. pneumophila MLVA-8(12) Genotypes from the West Bank with the International Data Base and A Study on A Larger Set of Strains from Germany and Israel
4.5. Conceivable Health Relevance of the L. pneumophila MLVA-8(12) Genotypes in the DWDS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murga, R.; Pruckler, J.M.; Forster, T.S.; Donlan, R.M.; Brown, E.; Fields, B.S. Role of biofilms in the survival of Legionella pneumophila in a model potable-water system. Microbiology 2001, 147, 3121–3126. [Google Scholar] [CrossRef] [PubMed]
- Declerck, P. Biofilms: The environmental playground of Legionella pneumophila. Environ. Microbiol. 2010, 12, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Rowbotham, T.J. Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae. J. Clin. Pathol. 1980, 33, 1179–1183. [Google Scholar] [CrossRef]
- Blatt, S.P.; Parkinson, M.D.; Pace, E.; Hoffman, P.; Dolan, D.; Lauderdale, P.; Zajac, R.A.; Melcher, G.P. Nosocomial legionnaires’ disease: Aspiration as a primary mode of disease acquisition. Am. J. Med. 1993, 95, 16–22. [Google Scholar] [CrossRef]
- Breiman, R.F.; Cozen, W.; Fields, B.S.; Mastro, T.D.; Carr, S.J.; Spika, J.S.; Mascola, L. Role of air sampling in investigation of an outbreak of Legionnaires’ disease associated with exposure to aerosols from an evaporative condenser. J. Infect. Dis. 1990, 161, 1257–1261. [Google Scholar] [CrossRef]
- Adeleke, A.A.; Fields, B.S.; Benson, R.F.; Daneshvar, M.I.; Pruckler, J.M.; Ratcliff, R.M.; Harrison, T.G.; Weyant, R.S.; Birtles, R.J.; Raoult, D.; et al. Legionella drozanskii sp. nov., Legionella rowbothamii sp. nov. and Legionella fallonii sp. nov.: Three unusual new Legionella species. Int. J. Syst. Evol. Microbiol. 2001, 51, 1151–1160. [Google Scholar] [CrossRef]
- Benson, R.F.; Fields, B.S. Classification of the genus Legionella. Semin. Respir. Infect. 1998, 13, 90–99. [Google Scholar]
- Yu, V.L.; Stout, J.E. Hospital characteristics associated with colonization of water systems by Legionella and risk of nosocomial Legionnaires’ disease: A cohort study of 15 hospitals. Infect. Control. Hosp. Epidemiol. 2000, 21, 434–435. [Google Scholar] [CrossRef]
- Amemura-Maekawa, J.; Kura, F.; Helbig, J.H.; Chang, B.; Kaneko, A.; Watanabe, Y.; Isobe, J.; Nukina, M.; Nakajima, H.; Kawano, K.; et al. Characterization of Legionella pneumophila isolates from patients in Japan according to serogroups, monoclonal antibody subgroups and sequence types. J. Med. Microbiol. 2010, 59, 653–659. [Google Scholar] [CrossRef]
- Messi, P.; Annalisa, B.; Immacolata, A.; Isabella, M.; De Niederhäusern, S.; Moreno, B. Protozoa and human macrophages infection by Legionella pneumophila environmental strains belonging to different serogroups. Arch. Microbiol. 2012, 195, 89–96. [Google Scholar] [CrossRef]
- Mavridou, A.; Smeti, E.; Mandilara, G.; Pappa, O.; Plakadonaki, S.; Grispou, E.; Polemis, M. Prevalence study of Legionella spp. contamination in Greek hospitals. Int. J. Environ. Heal. Res. 2008, 18, 295–304. [Google Scholar] [CrossRef]
- Fraser, D.W.; Tsai, T.R.; Orenstein, W.; Parkin, W.E.; Beecham, H.J.; Sharrar, R.G.; Harris, J.; Mallison, G.F.; Martin, S.M.; McDade, J.E.; et al. Legionnaires’ disease. N. Engl. J. Med. 1977, 297, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Buse, H.Y.; Morris, B.J.; Gomez-Alvarez, V.; Szabo, J.G.; Hall, J.S. Legionella diversity and spatiotemporal variation in the occurrence of opportunistic pathogens within a large building water system. Pathogens 2020, 9, 567. [Google Scholar] [CrossRef]
- Schijven, J.F.; Husman, A.D.R. Effect of climate changes on waterborne disease in The Netherlands. Water Sci. Technol. 2005, 51, 79–87. [Google Scholar] [CrossRef]
- Fiore, A.E.; Nuorti, J.P.; Levine, O.S.; Marx, A.; Weltman, A.C.; Yeager, S.; Benson, R.F.; Pruckler, J.; Edelstein, P.H.; Greer, P.; et al. Epidemic Legionnaires’ disease two decades later: Old sources, new diagnostic methods. Clin. Infect. Dis. 1998, 26, 426–433. [Google Scholar] [CrossRef]
- Graham, F.F.; Hales, S.; White, P.S.; Baker, M.G. Review Global seroprevalence of legionellosis-A systematic review and meta-analysis. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- International Standards Organization. Water Quality-Detection and Enumeration of Legionella; ISO 11731; International Standards Organization: Geneva, Switzerland, 1998; Available online: https://www.iso.org/standard/19653.html (accessed on 15 July 2016).
- Steinert, M.; Hentschel, U.; Hacker, J. Legionella pneumophila: An aquatic microbe goes astray. FEMS Microbiol. Rev. 2002, 26, 149–162. [Google Scholar] [CrossRef]
- Steinert, M.; Emödy, L.; Amann, R.; Hacker, J. Resuscitation of viable but nonculturable Legionella pneumophila Philadelphia JR32 by Acanthamoeba castellanii. Appl. Environ. Microbiol. 1997, 63, 2047–2053. [Google Scholar] [CrossRef] [PubMed]
- Nederbragt, A.J.; Balasingham, A.; Sirevåg, R.; Utkilen, H.; Jakobsen, K.S.; Anderson-Glenna, M.J. Multiple-locus variable-number tandem repeat analysis of Legionella pneumophila using multi-colored capillary electrophoresis. J. Microbiol. Methods 2008, 73, 111–117. [Google Scholar] [CrossRef]
- Villari, P.; Motti, E.; Farullo, C.; Torre, I. Comparison of conventional culture and PCR methods for the detection of Legionella pneumophila in water. Lett. Appl. Microbiol. 1998, 27, 106–110. [Google Scholar] [CrossRef]
- Reimer, A.R.; Au, S.; Schindle, S.; Bernard, K.A. Legionella pneumophila monoclonal antibody subgroups and DNA sequence types isolated in Canada between 1981 and 2009: Laboratory Component of National Surveillance. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 29, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Tijet, N.; Tang, P.; Romilowych, M.; Duncan, C.; Ng, V.; Fisman, D.N.; Jamieson, F.B.; Low, D.E.; Guyard, C. New endemic Legionella pneumophila serogroup I clones, Ontario, Canada. Emerg. Infect. Dis. 2010, 16, 447–454. [Google Scholar] [CrossRef]
- Mercante, J.W.; Winchell, J.M. Current and emerging Legionella diagnostics for laboratory and outbreak investigations. Clin. Microbiol. Rev. 2015, 28, 95–133. [Google Scholar] [CrossRef] [PubMed]
- Chenal-Francisque, V.; Diancourt, L.; Cantinelli, T.; Passet, V.; Tran-Hykes, C.; Bracq-Dieye, H.; Leclercq, A.; Pourcel, C.; Lecuit, M.; Brisse, S. Optimized multilocus variable-number tandem-repeat analysis assay and its complementarity with pulsed-field gel electrophoresis and multilocus sequence typing for Listeria monocytogenes clone identification and surveillance. J. Clin. Microbiol. 2013, 51, 1868–1880. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.; Wang, Q.; Yan, C.; Jeoffreys, N.; Wang, L.; Li, S.; Gilbert, G.L.; Sun, H. Molecular characterizations of PCR-positive Mycoplasma pneumoniae specimens collected from Australia and China. J. Clin. Microbiol. 2014, 52, 1478–1482. [Google Scholar] [CrossRef]
- Lindstedt, B.-A.; Torpdahl, M.; Vergnaud, G.; Le Hello, S.; Weill, F.-X.; Tietze, E.; Malorny, B.; Prendergast, D.M.; Ghallchoir, E.N.; Lista, R.F.; et al. Use of multilocus variable-number tandem repeat analysis (MLVA) in eight European countries, 2012. Eurosurveillance 2013, 18, 20385. [Google Scholar] [CrossRef]
- Pourcel, C.; Visca, P.; Afshar, B.; D’Arezzo, S.; Vergnaud, G.; Fry, N. Identification of variable-number tandem-repeat (VNTR) sequences in Legionella pneumophila and development of an optimized multiple-locus VNTR analysis typing scheme. J. Clin. Microbiol. 2007, 45, 1190–1199. [Google Scholar] [CrossRef]
- Sobral, D.; Le Cann, P.; Gerard, A.; Jarraud, S.; Lebeau, B.; Loisy-Hamon, F.; Vergnaud, G.; Pourcel, C. High-throughput typing method to identify a non-outbreak-involved Legionella pneumophila strain colonizing the entire water supply system in the town of Rennes, France. Appl. Environ. Microbiol. 2011, 77, 6899–6907. [Google Scholar] [CrossRef]
- Zayed, A.R. Ecology, virulence and genomics of Legionella pneumophila Isolates from the West Bank, Palestine. Ph.D. Thesis, Technical University of Braunschweig, Braunschweig, Germany, 2018; p. 107ff. Available online: https://publikationsserver.tu-braunschweig.de/receive/dbbs_mods_00066061 (accessed on 13 October 2020).
- Centers for Disease Control and Prevention CDC. Procedures for the Recovery of Legionella from the Environment Centers for Disease Control and Prevention. 2005; pp. 1–13. Available online: https://www.cdc.gov/legionella/labs/procedures-manual.html (accessed on 21 October 2020).
- Kahlisch, L.; Henne, K.; Groebe, L.; Draheim, J.; Höfle, M.G.; Brettar, I. Molecular analysis of the bacterial drinking water community with respect to live/dead status. Water Sci. Technol. 2010, 61, 9–14. [Google Scholar] [CrossRef]
- Senderovich, Y.; Gershtein, Y.; Halewa, E.; Halpern, M. Vibrio cholerae and Aeromonas: Do they share a mutual host? ISME J. 2008, 2, 276–283. [Google Scholar] [CrossRef]
- Helbig, J.H.; Bernander, S.; Castellani-Pastoris, M.; Etienne, J.; Gaia, V.; Lauwers, S.; Lindsay, D.; Luck, T.; Marques, S.; Mentula, S.; et al. Pan-European study on culture-proven Legionnaires’ disease: Distribution of Legionella pneumophila serogroups and monoclonal subgroups. Eur. J. Clin. Microbiol. Infect. Dis. 2002, 21, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Rajendram, D.; Ayenza, R.; Holder, F.; Moran, B.; Long, T.; Shah, H. Long-term storage and safe retrieval of DNA from microorganisms for molecular analysis using FTA matrix cards. J. Microbiol. Methods 2006, 67, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Visca, P.; D’Arezzo, S.; Ramisse, F.; Gelfand, Y.; Benson, G.; Vergnaud, G.; Fry, N.; Pourcel, C. Investigation of the population structure of Legionella pneumophila by analysis of tandem repeat copy number and internal sequence variation. Microbiology 2011, 157, 2582–2594. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pecellín, M. Structure and Virulence of Legionella pneumophila Populations from Freshwater Systems in Germany and Middle East. Ph.D. Thesis, Technische Universität Braunschweig, Braunschweig, Germany, 2016. Available online: https://publikationsserver.tu-braunschweig.de/servlets/solr/mods_nameIdentifier?q=mods.name:%22Pecell%C3%ADn%20Monjo%20Marina%22&owner=createdby:guest (accessed on 13 October 2020).
- Lück, P.C.; Fry, N.K.; Helbig, J.H.; Jarraud, S.; Harrison, T.G. Typing methods for Legionella. Recent Results Cancer Res. 2012, 954, 119–148. [Google Scholar]
- Chien, M.; Morozova, I.; Shi, S.; Sheng, H.; Chen, J.; Gomez, S.M.; Asamani, G.; Hill, K.; Nuara, J.; Feder, M.; et al. The Genomic Sequence of the Accidental Pathogen Legionella pneumophila. Science 2004, 305, 1966–1968. [Google Scholar] [CrossRef] [PubMed]
- Cazalet, C.; Rusniok, C.; Brüggemann, H.; Zidane, N.; Magnier, A.; Ma, L.; Tichit, M.; Jarraud, S.; Bouchier, C.; Vandenesch, F.; et al. Evidence in the Legionella pneumophila genome for exploitation of host cell functions and high genome plasticity. Nat. Genet. 2004, 36, 1165–1173. [Google Scholar] [CrossRef]
- Glöckner, G.; Albert-Weissenberger, C.; Weinmann, E.; Jacobi, S.; Schunder, E.; Steinert, M.; Hacker, J.; Heuner, K. Identification and characterization of a new conjugation/type IVA secretion system (trb/tra) of Legionella pneumophila Corby localized on two mobile genomic islands. Int. J. Med. Microbiol. 2008, 298, 411–428. [Google Scholar] [CrossRef]
- Zayed, A.R.; Butmeh, S.; Pecellin, M.; Salah, A.; Alalam, H.; Höfle, M.G.; Bitar, D.M.; Brettar, I. Biogeography and environmental drivers of Legionella pneumophila abundance and genotype composition across the West Bank: Relevance of a genotype-based ecology for understanding Legionella occurrence. Pathogens 2020. under review. [Google Scholar]
- Jaber, L.; Amro, M.; Abu Tair, H.; Bahader, S.A.; AlAlam, H.; Butmeh, S.; Abu Hilal, D.; Brettar, I.; Höfle, M.G.; Bitar, D.M. Comparison of in situ sequence type analysis of Legionella pneumophila in respiratory tract secretions and environmental samples of a hospital in East Jerusalem. Epidemiol. Infect. 2018, 146, 2116–2121. [Google Scholar] [CrossRef]
- Bonetta, S.; Ferretti, E.; Balocco, F.; Carraro, E.; Bonetta, S. Evaluation of Legionella pneumophila contamination in Italian hotel water systems by quantitative real-time PCR and culture methods. J. Appl. Microbiol. 2010, 108, 1576–1583. [Google Scholar] [CrossRef]
- Beauté, J.; Robesyn, E.; De Jong, B. Legionnaires’ disease in Europe: All quiet on the eastern front? Eur. Respir. J. 2013, 42, 1454–1458. [Google Scholar] [CrossRef] [PubMed]
- Yarom, R.; Sheinman, R.; Armon, R. Legionella pneumophila serogroup 3 prevalence in drinking water survey in Israel (2003–2007). Water Supply 2010, 10, 746–752. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2005, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Katsiaflaka, A.; Pournaras, S.; Kristo, I.; Mouchtouri, V.A.; Kyritsi, M.; Velonakis, E.; Vatopoulos, A.C.; Hadjichristodoulou, C. Epidemiological investigation of Legionella pneumophila serogroup 2 to 14 isolates from water samples by amplified fragment length polymorphism and sequence-based typing and detection of virulence traits. Appl. Environ. Microbiol. 2016, 82, 6102–6108. [Google Scholar] [CrossRef]
- Ghrairi, T.; Chaftar, N.; Jarraud, S.; Berjeaud, J.M.; Hani, K.; Frère, J. Diversity of legionellae strains from Tunisian hot spring water. Res. Microbiol. 2013, 164, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Al-Matawah, Q.A.; Al-Zenki, S.F.; Qasem, J.A.; Al-Waalan, T.E.; Ben Heji, A.H. Detection and quantification of Legionella pneumophila from water systems in Kuwait residential facilities. J. Pathog. 2012, 13, 8389. [Google Scholar] [CrossRef]
- Ma’Ayeh, S.Y.; Al-Hiyasat, A.S.; Hindiyeh, M.; Khader, Y.; Hindiyeh, M.Y. Legionella pneumophila contamination of a dental unit water line system in a dental teaching centre. Int. J. Dent. Hyg. 2008, 6, 48–55. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, S.; Sharaby, Y.; Pecellín, M.; Brettar, I.; Höfle, M.; Halpern, M. Spatial distribution of Legionella pneumophila MLVA-genotypes in a drinking water system. Water Res. 2015, 77, 119–132. [Google Scholar] [CrossRef]
- Douterelo, I.; Boxall, J.B.; Deines, P.; Sekar, R.; Fish, K.E.; Biggs, C.A. Methodological approaches for studying the microbial ecology of drinking water distribution systems. Water Res. 2014, 65, 134–156. [Google Scholar] [CrossRef]
- Berry, D.; Xi, C.; Raskin, L. Microbial ecology of drinking water distribution systems. Curr. Opin. Biotechnol. 2006, 17, 297–302. [Google Scholar] [CrossRef]
- Reuter, S.; Harrison, T.G.; Köser, C.U.; Ellington, M.J.; Smith, G.P.; Parkhill, J.; Peacock, S.J.; Bentley, S.D.; Török, M.E. A pilot study of rapid whole-genome sequencing for the investigation of a Legionella outbreak. BMJ Open 2013, 3, e002175. [Google Scholar] [CrossRef] [PubMed]
- Whiley, H.; Keegan, A.; Fallowfield, H.; Ross, K. Uncertainties associated with assessing the public health risk from Legionella. Front. Microbiol. 2014, 5, 501. [Google Scholar] [CrossRef]
- Oliver, J.D. The viable but nonculturable state in bacteria. J. Microbiol. 2005, 93–100. [Google Scholar]
- Lee, H.K.; Kang, Y.H.; Yu, J.Y. Genomic diversity of Legionella pneumophila serogroup 1 from environmental water sources and clinical specimens using pulsed-field gel electrophoresis (PFGE) from 1985 to 2007, Korea. J. Microbiol. 2010, 48, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Uzel, A.; Uçar, F.; Hameş-Kocabaş, E.E. Prevalence of Legionella pneumophila serogroup 1 in water distribution systems in İzmir province of Turkey. APMIS 2005, 113, 664–669. [Google Scholar] [CrossRef]
- Harrison, T.G.; Afshar, B.; Doshi, N.; Fry, N.K.; Lee, J.V. Distribution of Legionella pneumophila serogroups, monoclonal antibody subgroups and DNA sequence types in recent clinical and environmental isolates from England and Wales (2000–2008). Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 781–791. [Google Scholar] [CrossRef]
- Doleans, A.; Aurell, H.; Reyrolle, M.; Lina, G.; Freney, J.; Vandenesch, F.; Etienne, J.; Jarraud, S. Clinical and environmental distributions of Legionella strains in France are different. J. Clin. Microbiol. 2004, 42, 458–460. [Google Scholar] [CrossRef]
- Coscolla, M.; González-Candelas, F. Comparison of clinical and environmental samples of Legionella pneumophila at the nucleotide sequence level. Infect. Genet. Evol. 2009, 9, 882–888. [Google Scholar] [CrossRef]
- Martin-Loeches, I.; Torres, A.; Rinaudo, M.; Terraneo, S.; De Rosa, F.; Ramirez, P.; Díaz, E.; Fernandez-Barat, L.; Bassi, G.L.L.; Ferrer, M. Resistance patterns and outcomes in intensive care unit (ICU)-acquired pneumonia. Validation of European Centre for Disease Prevention and Control (ECDC) and the Centers for Disease Control and Prevention (CDC) classification of multidrug resistant organisms. J. Infect. 2015, 70, 213–222. [Google Scholar] [CrossRef]
- Yu, V.L.; Plouffe, J.F.; Pastoris, M.C.; Stout, J.E.; Schousboe, M.; Widmer, A.; Summersgill, J.; File, T.; Heath, C.M.; Paterson, D.L.; et al. Distribution of Legionella species and serogroups isolated by culture in patients with sporadic community-acquired Legionellosis: An international collaborative survey. J. Infect. Dis. 2002, 186, 127–128. [Google Scholar] [CrossRef]
- Fendukly, F.; Bernander, S.; Hanson, H.-S. Nosocomial Legionnaires’ disease caused by Legionella pneumophila serogroup 6: Implication of the sequence-based typing method (SBT). Scand. J. Infect. Dis. 2007, 39, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhou, H.; Chen, R.; Qin, T.; Ren, H.; Liu, B.; Ding, X.; Sha, D.; Zhou, W. Legionnaires’ Disease caused by Legionella pneumophila serogroups 5 and 10, China. Emerg. Infect. Dis. 2014, 20, 1242–1243. [Google Scholar] [CrossRef] [PubMed]
- Cazalet, C.; Jarraud, S.; Ghavi-Helm, Y.; Kunst, F.; Glaser, P.; Etienne, J.; Buchrieser, C. Multigenome analysis identifies a worldwide distributed epidemic Legionella pneumophila clone that emerged within a highly diverse species. Genome Res. 2008, 18, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Kozak-Muiznieks, N.A.; Lucas, C.E.; Brown, E.; Pondo, T.; Taylor, T.H.; Frace, M.; Miskowski, D.; Winchell, J.M. Prevalence of sequence types among clinical and environmental isolates of Legionella pneumophila serogroup 1 in the United States from 1982 to 2012. J. Clin. Microbiol. 2013, 52, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Amemura-Maekawa, J.; Kikukawa, K.; Helbig, J.H.; Kaneko, S.; Suzuki-Hashimoto, A.; Furuhata, K.; Chang, B.; Murai, M.; Ichinose, M.; Ohnishi, M.; et al. Distribution of monoclonal antibody subgroups and sequence-based types among Legionella pneumophila serogroup 1 isolates derived from cooling tower water, bathwater, and soil in Japan. Appl. Environ. Microbiol. 2012, 78, 4263–4270. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.K.; Shim, J.I.; Kim, H.E.; Yu, J.Y.; Kang, Y.H. Distribution of Legionella species from environmental water sources of public facilities and genetic diversity of L. pneumophila serogroup 1 in South Korea. Appl. Environ. Microbiol. 2010, 76, 6547–6554. [Google Scholar] [CrossRef] [PubMed]
- Borchardt, J.; Helbig, J.H.; Lück, P.C. Occurrence and distribution of sequence types among Legionella pneumophila strains isolated from patients in Germany: Common features and differences to other regions of the world. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 27, 29–36. [Google Scholar] [CrossRef]
- Chasqueira, M.J.; Rodrigues, L.; Nascimento, M.; Ramos, M.; Marques, T. Genetic diversity and evolutionary relationships among Legionella pneumophila clinical isolates, Portugal, 1987 to 2012. Eurosurveillance 2014, 19, 20965. [Google Scholar] [CrossRef]
- Sánchez-Busó, L.; Coscollá, M.; Pinto-Carbó, M.; Catalán, V.; González-Candelas, F. Genetic characterization of Legionella pneumophila isolated from a common watershed in Comunidad Valenciana, Spain. PLoS ONE 2013, 8, e61564. [Google Scholar] [CrossRef]
- David, S.; Afshar, B.; Mentasti, M.; Ginevra, C.; Podglajen, I.; Harris, S.R.; Chalker, V.J.; Jarraud, S.; Harrison, T.G.; Parkhill, J. Seeding and establishment of Legionella pneumophila in hospitals: Implications for genomic investigations of nosocomial Legionnaires’ disease. Clin. Infect. Dis. 2017, 64, 1251–1259. [Google Scholar] [CrossRef]
- Fontana, S.; Scaturro, M.; Rota, M.C.; Caporali, M.G.; Ricci, M.L. Molecular typing of Legionella pneumophila serogroup 1 clinical strains isolated in Italy. Int. J. Med. Microbiol. 2014, 304, 597–602. [Google Scholar] [CrossRef] [PubMed]
- García-Vidal, C.; Labori, M.; Viasus, D.; Simonetti, A.; Garcia-Somoza, D.; Dorca, J.; Gudiol, F.; Carratalà, J. Rainfall is a risk factor for sporadic cases of Legionella pneumophila pneumonia. PLoS ONE 2013, 8, e61036. [Google Scholar] [CrossRef] [PubMed]
- Gaia, V.; Fry, N.K.; Harrison, T.G.; Peduzzi, R. Sequence-based typing of Legionella pneumophila serogroup 1 offers the potential for true portability in Legionellosis outbreak investigation. J. Clin. Microbiol. 2003, 41, 2932–2939. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, C.; Reyrolle, M.; Dubrou, S.; Forey, F.; Decludt, B.; Goulvestre, C.; Matsiota-Bernard, P.; Etienne, J.; Nauciel, C. Single clonal origin of a high proportion of Legionella pneumophila serogroup 1 isolates from patients and the environment in the area of Paris, France, over a 10-year period. J. Clin. Microbiol. 1999, 37, 2652–2655. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Rusniok, C.; Mentasti, M.; Gomez-Valero, L.; Harris, S.R.; Lechat, P.; Lees, J.; Ginevra, C.; Glaser, P.; Ma, L.; et al. Multiple major disease-associated clones of Legionella pneumophila have emerged recently and independently. Genome Res. 2016, 26, 1555–1564. [Google Scholar] [CrossRef]
- David, S.; Mentasti, M.; Tewolde, R.; Aslett, M.; Harris, S.R.; Afshar, B.; Underwood, A.; Fry, N.K.; Parkhill, J.; Harrison, T.G. Evaluation of an optimal epidemiological typing scheme for Legionella pneumophila with whole-genome sequence data using validation guidelines. J. Clin. Microbiol. 2016, 54, 2135–2148. [Google Scholar] [CrossRef]
- Sharaby, Y.; Rodríguez-Martínez, S.; Pecellin, M.; Sela, R.; Peretz, A.; Höfle, M.G.; Halpern, M.; Brettar, I. Virulence traits of environmental and clinical Legionella pneumophila Multilocus Variable-Number Tandem-Repeat Analysis (MLVA) Genotypes. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed]
- Sharaby, Y.; Nitzan, O.; Brettar, I.; Höfle, M.G.; Peretz, A.; Halpern, M. Antimicrobial agent susceptibilities of Legionella pneumophila MLVA-8 genotypes. Sci. Rep. 2019, 9, 6138. [Google Scholar] [CrossRef]
- Sharaby, Y.; Rodríguez-Martínez, S.; Oks, O.; Pecellin, M.; Mizrahi, H.; Peretz, A.; Brettar, I.; Höfle, M.G.; Halpern, M. Temperature-dependent growth modeling of environmental and clinical Legionella pneumophila Multilocus Variable-Number Tandem-Repeat Analysis (MLVA) genotypes. Appl. Environ. Microbiol. 2017, 83, e03295-16. [Google Scholar] [CrossRef]
- García, K.; Gavilán, R.G.; Höfle, M.G.; Martínez-Urtaza, J.; Espejo, R.T. Microevolution of pandemic Vibrio parahaemolyticus assessed by the number of repeat units in short sequence tandem repeat regions. PLoS ONE 2012, 7, e30823. [Google Scholar] [CrossRef] [PubMed]
- Husseini, H. Palestinian Water Authority: Developments and Challenges-Legal Framework and Capacity. In Water Resources in the Middle East; Springer Science and Business Media LLC: New York, NY, USA, 2007; pp. 301–308. [Google Scholar]
- Sharaby, Y.; Rodríguez-Martínez, S.; Höfle, M.; Brettar, I.; Halpern, M. Quantitative microbial risk assessment of Legionella pneumophila in a drinking water supply system in Israel. Sci. Total. Environ. 2019, 671, 404–410. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hospital | Water Type | HPC (CFU/L) at 37 °C ± SD | HPC (CFU/L) at 22 °C ± SD | Leg. Count (CFU/L) ± SD | Tempe-rature °C ± SD | pH ± SD | Conductivity µS/cm ± SD | Chlorine mg/L ± SD | Hardness (mg/L) |
|---|---|---|---|---|---|---|---|---|---|
| A | Cold | 2.1 × 104 ± 1.6 × 102 | 7.4 × 103 ± 7.1 × 101 | 4.3 × 101 ± 1.06 × 102 | 23.1 ± 2.4 | 7.7 ± 0.2 | 780 ± 158 | 0.5 ± 0.3 | 290 ± 36 |
| Hot | NA | NA | NA | NA | NA | NA | NA | NA | |
| B | Cold | 2.0 × 104 ± 2.2 × 102 | 1.4 × 103 ± 1.4 × 102 | BD | 24.3 ± 2 | 8.0 ± 0.2 | 639 ± 86 | 0.4 ± 0.2 | 299 ± 16 |
| Hot | 2.2 × 102 ± 2.9 × 101 | 1.1 × 102 ± 1.4 × 101 | BD | 51.9 ± 16.3 | 7.8 ± 0.2 | 700 ± 30 | NA | NA | |
| C | Cold | 1.5 × 104 ± 2.0 × 102 | 1.3 × 103 ± 2.0 × 102 | BD | 23.1 ± 2.5 | 8.0 ± 0 | 707 ± 60 | 0.5 ± 0 | 299 ± 16 |
| Hot | NA | NA | NA | NA | NA | NA | NA | NA | |
| D | Cold | 1.6 × 105 ± 8.7 × 103 | 1.2 × 104 ± 8.9 × 103 | BD | 22.5 ± 1.6 | 7.6 ± 0.1 | 509 ± 118 | 0.7 ± 0.35 | 229 ± 26 |
| Hot | NA | NA | NA | NA | NA | NA | NA | NA | |
| E | Cold | 1.7 × 104 ± 2.0 × 102 | 1.5 × 103 ± 2.0 × 101 | BD | 25.2 ± 5.2 | 7.8 ± 0.3 | 699 ± 82 | 0.2 ± 0.05 | 269 ± 11 |
| Hot | 1.6 × 104 ± 3.5 × 102 | 4.1 × 102 ± 4.2 × 101 | BD | 47.9 ± 5.1 | 7.8 ± 0.2 | 761 ± 30 | NA | NA | |
| F | Cold | 1.1 × 105 ± 1.5 × 102 | 6.3 × 104 ± 7.1 × 102 | 1.5 × 102 ± 2.3 × 102 | 24.7 ± 2.9 | 7.8 ± 0.2 | 556 ± 106 | 0.4 ± 0.2 | 261 ± 3 |
| Hot | 2.5 × 104 ± 2.6 × 102 | 3.2 × 103 ± 3.3 × 101 | 9.1 × 101 ± 1.3 × 102 | 43.3 ± 5.7 | 7.8 ± 0.2 | 583 ± 93 | NA | NA | |
| G | Cold | 2.9 × 104 ± 2.0 × 102 | 2.2 × 103 ± 2.2 × 102 | 8.3 × 100 ± 2.0 × 101 | 21.6 ± 2.7 | 8.0 ± 0.3 | 648 ± 179 | 0.4 ± 0.2 | 266 ± 19 |
| Hot | 1.8 × 104 ± 1.5 × 102 | 2.7 × 103 ± 1.8 × 101 | BD | 38.6 ± 6.8 | 8.0 ± 0.3 | 596 ± 190 | NA | NA | |
| H | Cold | 2.7 × 104 ± 2.5 × 102 | 1.4 × 103 ± 1.3 × 102 | BD | 20.8 ± 1.3 | 7.8 ± 0.1 | 707 ± 191 | 0.4 ± 0.2 | 266 ± 19 |
| Hot | NA | NA | NA | NA | NA | NA | NA | NA |
| CDA 1 | CIA 2 | ||||||
|---|---|---|---|---|---|---|---|
| Year | L. pneumo-phila Isolates/Total Number 3 | % of Isolates | Leg. Counts (Mean of Cold Water) (CFU/l)±SD | Legionella spp (Lgsp) Positive Samples/Total Number | % of Positive Samples | L. pneumo-phila (Lpn) Positive Samples/Total Number | % of Positive Samples |
| 2012 | 96/409 | 23.5 | NA | 43/53 | 81.1 | 36/53 | 67.9 |
| 2013 | 30/346 | 8.7 | NA | 64/106 | 60.4 | 55/106 | 51.9 |
| 2014 | 71/453 | 15.7 | NA | 102/138 | 73.9 | 79/138 | 57.2 |
| Hospital | |||||||
| A | 30/150 | 20 | 4.3 × 101 ± 1.1 × 102 | 29/36 | 80.6 | 23/36 | 63.9 |
| B | 35/156 | 22.4 | BD | 28/42 | 66.7 | 20/42 | 47.6 |
| C | 5/150 | 3.3 | BD | 19/36 | 52.8 | 11/36 | 30.6 |
| D | 18/90 | 20 | BD | 16/21 | 76.2 | 13/21 | 61.9 |
| E | 11/156 | 7.1 | BD | 17/42 | 40.5 | 13/42 | 31 |
| F | 41/156 | 26.3 | 1.5 × 102 ± 2.3 × 102 | 37/42 | 88.1 | 34/42 | 81 |
| G | 35/156 | 22.4 | 8.3 × 100 ± 2.0 × 102 | 39/42 | 92.9 | 38/42 | 90.5 |
| H | 6/150 | 4 | BD | 24/36 | 66.7 | 18/36 | 50 |
| AQU | 16/44 | 36.4 | NA | NA | NA | NA | NA |
| Sample type | |||||||
| Water | 6/72 | 8.3 | 42/72 | 58.3 | 36/72 | 50 | |
| Biofilm | 191/1136 | 16.8 | 167/225 | 74.2 | 134/225 | 59.5 |
| West Bank | |||
|---|---|---|---|
| VNTR | No. of Repeats | HGDI 1 (CI 95%) | Null Alleles (%) |
| Lpms1 | 4 | 0.528 (0.459–0.596) | 0 |
| Lpms3 | 2 | 0.461 (0.420–0.502) | 0 |
| Lpms13 | 5 | 0.579 (0.506–0.652) | 0 |
| Lpms17 | 2 | 0.115 (0.053–0.178) | 0 |
| Lpms19 | 2 | 0.022 (1.000–0.053) | 1.11 |
| Lpms31 | 6 | 0.576 (0.513–0.639) | 1.67 |
| Lpms33 | 4 | 0.575 (0.506–0.643) | 0 |
| Lpms34 | 4 | 0.503 (0.429–0.577) | 2.22 |
| Lpms35 | 6 | 0.687 (0.641–0.733) | 1.67 |
| Lpms38 | 3 | 0.249 (0.168–0.330) | 4.44 |
| Lpms39 | 3 | 0.509 (0.445–0.574) | 0 |
| Lpms40 | 3 | 0.493 (0.444–0.541) | 3.33 |
| Lpms44 | 3 | 0.498 (0.463–0.533) | 0 |
| Serogroup | mAb 1 Subgroup | L. pneumophila Isolates | |
|---|---|---|---|
| No. | Frequency (%) | ||
| 1 | OLDA | 10 | 5.6 |
| 1 | NA 2 | 101 | 56.1 |
| Total Sg1 | 111 | 61.6 | |
| 6 | Dresden | 54 | 30.0 |
| 8 | NA 2 | 6 | 3.3 |
| 10 | NA 2 | 2 | 1.1 |
| (2–14) | NA 2 | 7 | 3.9 |
| Total non-Sg1 | 69 | 38.3 | |
| Total | 180 | 100 | |
| Location | MLVA-8 (12)-Genotype | No of Strains per Genotype (%) | Sg-MAb | MLVA- Clonal Complex (VACC) | No. of Strains per VACC (%) |
|---|---|---|---|---|---|
| Hospital A | Gt4(17) | 20 (71) | 1 | VACC1 | 20(71) |
| Gt16(1) | 4 (14) | 6 Dresden | VACC5 | 5(18) | |
| Gt13(72) | 3 (11) | 6 Dresden | VACC2 | 3(11) | |
| Gt16(6) | 1 (4) | (2-14) | 0 | 0 | |
| Total | 28(100) | 0 | 0 | 28(100) | |
| Hospital B | Gt4(17) | 21(66) | 1 | VACC1 | 24(75) |
| Gt9(92) | 7(22) | 6 Dresden | VACC11 | 7(22) | |
| Gt63(83) | 2(6) | 1 | VACC2 | 1(3) | |
| Gt4(16) | 1(3) | 1 | 0 | 0 | |
| Gt13(72) | 1(3) | 6 Dresden | 0 | 0 | |
| Total | 32(100) | 0 | 0 | 32(100) | |
| Hospital C | Gt4(17) | 4(80) | 1 | VACC1 | 4(80) |
| Gt10(93) | 1(20) | 6 Dresden | VACC11 | 1(20) | |
| Total | 5(100) | 0 | 0 | 5(100) | |
| Hospital D | Gt4(17) | 15(83) | 1 | VACC1 | 15(83) |
| Gt13(72) | 3(17) | 6 Dresden | VACC2 | 3(17) | |
| Total | 18(100) | 0 | 0 | 18(100) | |
| Hospital E | Gt13(143) | 2(20) | 10 | VACC2 | 5(50) |
| Gt64(72) | 2(20) | 6 Dresden | VACC5 | 3(30) | |
| Gt8(7) | 2(20) | (2-14) | VACC1 | 2(20) | |
| Gt4(17) | 2(20) | 1 | 0 | 0 | |
| Gt24(68) | 1(10) | (2-14) | 0 | 0 | |
| Gt16(3) | 1(10) | (2-14) | 0 | 0 | |
| Total | 10(100) | 0 | 0 | 10(100) | |
| Hospital F | Gt10(93) | 14(44) | 6 Dresden | VACC11 | 21(66) |
| Gt10(141) | 6(19) | 6 Dresden | VACC1 | 6(19) | |
| Gt4(17) | 6(19) | 1 | VACC2 | 5(16) | |
| Gt64(74) | 3(9) | 6 Dresden | 0 | 0 | |
| Gt55(94) | 1(3) | 6 Dresden | 0 | 0 | |
| Gt64(72) | 1(3) | 6 Dresden | 0 | 0 | |
| Gt38(109) | 1(3) | 1 | 0 | 0 | |
| Total | 32(100) | 0 | 0 | 32(100) | |
| Hospital G | Gt6(18) | 30(88) | 1 | VACC1 | 31(91) |
| Gt6(15) | 1(3) | 1 | VACC5 | 3(9) | |
| Gt16(1) | 1(3) | 6 Dresden | 0 | 0 | |
| Gt8(142) | 1(3) | (2-14) | 0 | 0 | |
| Gt8(23) | 1(3) | (2-14) | 0 | 0 | |
| Total | 34(100) | 0 | 0 | 34(100) | |
| Hospital H | Gt40(47) | 3(50) | 6 Dresden | VACC5 | 3(50) |
| Gt10(93) | 1(17) | 6 Dresden | VACC11 | 2(33) | |
| Gt9(92) | 1(17) | 6 | VACC1 | 1(17) | |
| Gt63(83) | 1(17) | 1 | 0 | 0 | |
| Total | 6(100) | 0 | 0 | 6(100) | |
| AQU * | Gt4(17) | 6(40) | 1 OLDA | VACC1 | 13(87) |
| Gt12(84) | 5(33) | 8 | VACC2 | 2(13) | |
| Gt4(20) | 1(7) | 1 OLDA | 0 | 0 | |
| Gt11(87) | 1(7) | 8 | 0 | 0 | |
| Gt13(106) | 1(7) | 6 | 0 | 0 | |
| Gt84(106) | 1(7) | 6 | 0 | 0 | |
| Total | 15(100) | 0 | 0 | 15(100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zayed, A.R.; Pecellin, M.; Salah, A.; Alalam, H.; Butmeh, S.; Steinert, M.; Lesnik, R.; Brettar, I.; Höfle, M.G.; Bitar, D.M. Characterization of Legionella pneumophila Populations by Multilocus Variable Number of Tandem Repeats (MLVA) Genotyping from Drinking Water and Biofilm in Hospitals from Different Regions of the West Bank. Pathogens 2020, 9, 862. https://doi.org/10.3390/pathogens9110862
Zayed AR, Pecellin M, Salah A, Alalam H, Butmeh S, Steinert M, Lesnik R, Brettar I, Höfle MG, Bitar DM. Characterization of Legionella pneumophila Populations by Multilocus Variable Number of Tandem Repeats (MLVA) Genotyping from Drinking Water and Biofilm in Hospitals from Different Regions of the West Bank. Pathogens. 2020; 9(11):862. https://doi.org/10.3390/pathogens9110862
Chicago/Turabian StyleZayed, Ashraf R., Marina Pecellin, Alaa Salah, Hanna Alalam, Suha Butmeh, Michael Steinert, Rene Lesnik, Ingrid Brettar, Manfred G. Höfle, and Dina M. Bitar. 2020. "Characterization of Legionella pneumophila Populations by Multilocus Variable Number of Tandem Repeats (MLVA) Genotyping from Drinking Water and Biofilm in Hospitals from Different Regions of the West Bank" Pathogens 9, no. 11: 862. https://doi.org/10.3390/pathogens9110862
APA StyleZayed, A. R., Pecellin, M., Salah, A., Alalam, H., Butmeh, S., Steinert, M., Lesnik, R., Brettar, I., Höfle, M. G., & Bitar, D. M. (2020). Characterization of Legionella pneumophila Populations by Multilocus Variable Number of Tandem Repeats (MLVA) Genotyping from Drinking Water and Biofilm in Hospitals from Different Regions of the West Bank. Pathogens, 9(11), 862. https://doi.org/10.3390/pathogens9110862