The Antiviral Effect of Indonesian Medicinal Plant Extracts Against Dengue Virus In Vitro and In Silico

 and
and
Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Extract Preparation
4.2. Cell Culture and Virus
4.3. Determination of Cytotoxicity Assay
4.4. Determination of Infectivity Assay
4.5. The Selectivity Index Analysis
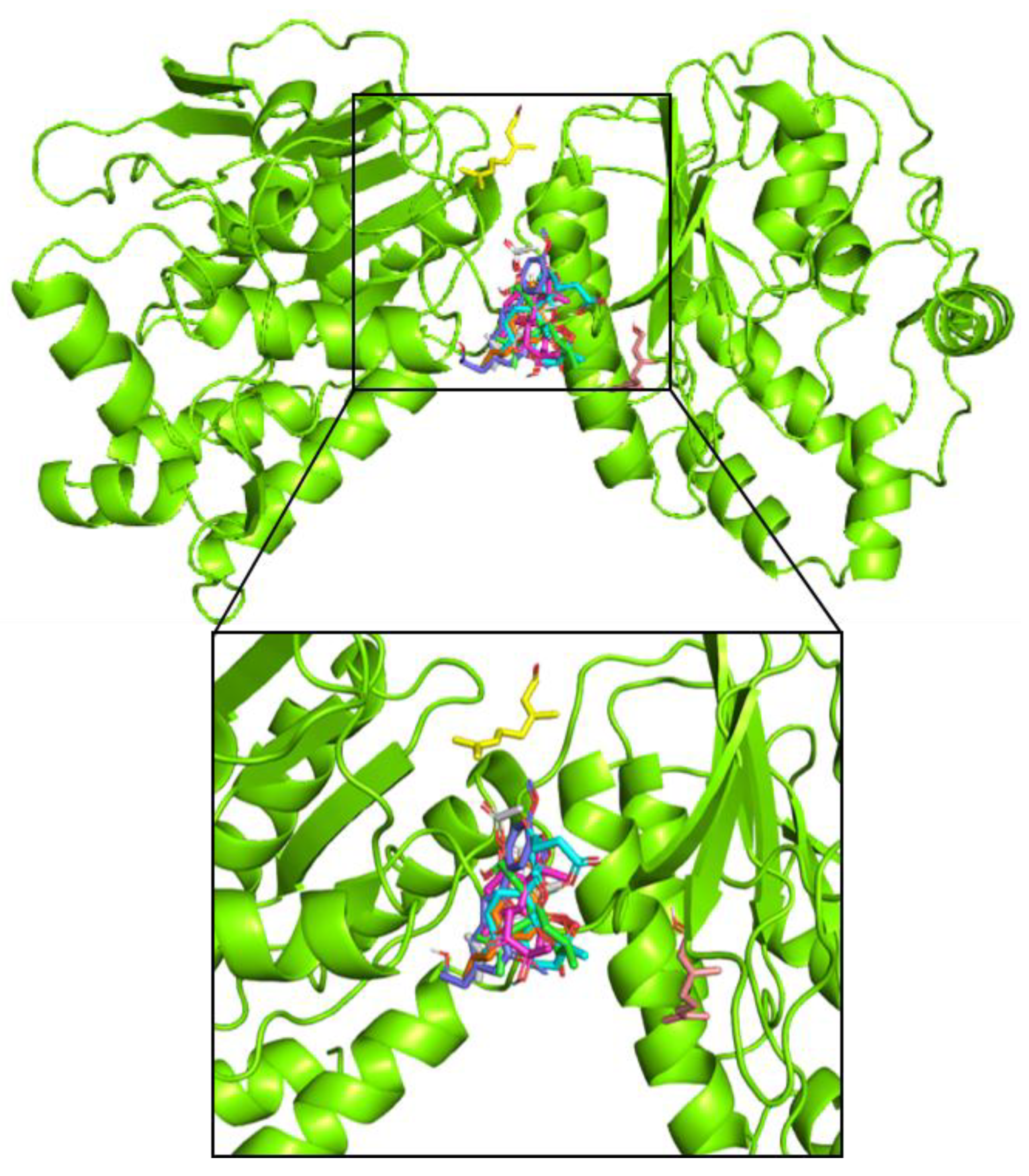
4.6. Molecular Docking Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.I.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Zhou, Z.; Wen, Z.; Liu, Y.; Zeng, C.; Xiao, D. Global Epidemiology of Dengue Outbreaks in 1990–2015: A Systematic Review and Meta-Analysis. Front. Cell. Infect. Microbiol. 2017, 7, 317. [Google Scholar] [CrossRef] [PubMed]
- Wahyono, T.Y.M.; Nealon, J.; Beucher, S.; Prayitno, A.; Moureae, A.; Nawawi, S. Indonesian dengue burden estimates: Review of evidence by an expert panel. Epidemiol. Infect. 2017, 145, 2324–2329. [Google Scholar] [CrossRef] [PubMed]
- Suwandono, A.; Kosasih, H.; Kusriastuti, R.; Harun, S.; Ma’roef, C.; Wuryadi, S.; Herianto, B.; Yuwono, D.; Porter, K.R.; Beckett, C.G.; et al. Four dengue virus serotypes found circulating during an outbreak of dengue fever and dengue haemorrhagic fever in Jakarta, Indonesia, during 2004. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Dewi, B.E.; Naiggolan, L.; Putri, D.H.; Rachmayanti, N.; Albar, S.; Indriastuti, N.T.; Sjamsuridzal, W.; Sudiro, T.M. Characterization of dengue virus serotype 4 infection in Jakarta, Indonesia. Southeast Asian J. Trop. Med. Public Health 2014, 45, 53–61. [Google Scholar] [PubMed]
- Badurdeen, S.; Valladares, D.B.; Farrar, J.; Gozzer, E.; Kroeger, A.; Kuswara, N.; Ranzinger, S.R.; Tinh, H.T.; Leite, P.; Mahendradhata, Y.; et al. Sharing experiences: Towards an evidence based model of dengue surveillance and outbreak response in Latin America and Asia. BMC Public Health 2013, 13, 607. [Google Scholar] [CrossRef] [PubMed]
- Wilder-Smith, A.; Renhorn, K.E.; Tissera, H.; Abu Bakar, S.; Alphey, L.; Kittayapong, P.; Lindsay, S.; Logan, J.; Hatz, C.; Reiter, P.; et al. DengueTools: Innovative tools and strategies for the surveillance and control of dengue. Glob. Health Action 2012, 5, 17273. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Alvarez, M.; Halstead, S.B. Secondary infection as a risk factor for dengue hemorrhagic fever/dengue shock syndrome: An historical perspective and role of antibody-dependent enhancement of infection. Arch. Virol. 2013, 158, 1445–1459. [Google Scholar] [CrossRef] [PubMed]
- Cos, P.; Vlietinck, A.J.; Berghe, D.V.; Maes, L. Anti-infective potential of natural products: How to develop a stronger in vitro ‘proof of concept’. J. Ethnopharmacol. 2006, 106, 290–302. [Google Scholar] [CrossRef]
- World Health Organization. Comprehensive Guidelines for Prevention and Control of Dengue and Dengue Haemorrhagic Fever: Revised and Expanded Edition; SEARO: New Delhi, India, 2011. [Google Scholar]
- Ghosh, A.; Dar, L. Dengue vaccines: Challenges, development, current status and prospects. Indian J. Med. Microbiol. 2015, 33, 3–15. [Google Scholar]
- De Maio, F.A.; Risso, G.; Iglesias, N.G.; Shah, P.; Pozzi, B.; Gebhard, L.G.; Mammi, P.; Mancini, E.; Yanovsky, M.J.; Andino, R.; et al. The dengue virus NS5 protein intrudes in the cellular spliceosome and modulates splicing. PLoS Pathog. 2016, 12, e1005841. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Rawat, P. Evolving herbal formulations in management of dengue fever. J. Ayurveda Integr. Med. 2017, 8, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Texeira, R.R.; Pereira, W.L.; Oliveira, A.F.C.S.; Silva, A.M.; Oliveira, A.S.; Silva, M.L. Natural Products as Source of Potential Dengue Antivirals. Molecules 2014, 19, 8151–8176. [Google Scholar] [CrossRef] [PubMed]
- Monika, J.A.; Sudipta, C.; Malleswara, R.E.; Lilly, G. Effect of Hippophae Rhamnoides Leaf Extract against Dengue Virus Infection in U937 Cells. Virol-Mycol 2016, 5, 157. [Google Scholar]
- Dewi, B.E.; Angelina, M.; Meilawati, L.; Hartati, S.; Dewijanti, I.D.; Santi, M.R.; Desti, H.; Sudiro, T.M. Antiviral Effect of Pterocarpus indicus Willd Leaves Extract Against Replication of Dengue Virus (DENV) In Vitro. J. Trop. Life Sci. 2018, 2018. 8, 55–61. [Google Scholar] [CrossRef][Green Version]
- Rajesh, K.G.; Piya, P.M.; Hindol, M.; Deepu, D.; Santhosha, D.; Snehlata, N.; Govindakarnavar, A. Herbal plants and plant preparations as remedial approach for viral diseases. Virus Dis. 2015, 26, 225–236. [Google Scholar]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Simpson, D.; Amos, S. Other Plant Metabolites. In Pharmacognosy; Academic Press: Cambridge, MA, USA, 2017; pp. 267–280. [Google Scholar]
- Megawati, M. Resorcinol Compounds Isolated form the Bark of Myristica fatua Houtt. J. Indones. Pharm. 2017, 28, 82–90. [Google Scholar] [CrossRef][Green Version]
- Fajriah, S.; Darmawan, A.; Hudiyono, S.; Kosela, S.; Hanafi, M. New cytotoxic compounds from Myristica fatua Houtt leaves against MCF-7 cell lines. Phytochem. Lett. 2017, 20, 36–39. [Google Scholar] [CrossRef]
- Xingang, Y.; Yun, L.; Songxin, G.; Wenyu, W.; Shijun, H.; Qing, Z.; Min, Z.; Kutty, S.N.; Xiaoguang, C.; Shuwen, L. Tatanan A from the Acorus calamus L. root inhibited dengue virus proliferation and infections. Phytomedicine 2018, 42, 258–267. [Google Scholar]
- Shah, G.; Shri, R.; Panchal, V.; Sharma, N.; Singh, B.; Mann, A.S. Scientific basis for the therapeutic use of Cymbopogon citratus, stapf (Lemon grass). J. Adv. Pharm. Technol. Res. 2011, 2, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Yucharoen, R.; Anuchapreeda, S.; Tragoolpua, Y. Anti-herpes simplex virus activity of extracts from the culinary herbs Ocimum sanctum L., Ocimum basilicum L. and Ocimum americanum L. Afr. J. Biotechnol. 2011, 10, 860–866. [Google Scholar]
- Saptawati, L.; Febrinasari, R.P.; Yudhani, R.D.; Yono, H.; Faza, A.G.; Luthfiani, S.; Ummiyati, H.S.; Sudiro, T.M.; Dewi, B.E. In vitro study of eight Indonesian plants extracts as anti-dengue virus. Health Sci. J. Indones. 2017, 8, 12–18. [Google Scholar] [CrossRef]
- Sanghai, V.D.N.; Kulkarni, S.R.; Sanghai, N.N. Screening of Antiviral Compounds from Plants—A Review. J. Pharm. Res. 2014, 8, 1050–1058. [Google Scholar]
- Zandi, K.; Lim, T.H.; Rahim, N.A.; Shu, M.H.; Teoh, B.T.; Sam, S.S.; Danlami, M.B.; Tan, K.K.; Abubakar, S. Extract of Scutellaria baicalensis inhibits dengue virus replication. BMC Complement. Altern. Med. 2013, 13, 91. [Google Scholar] [CrossRef] [PubMed]
- Viveka, M.R.; Chandrashekar, K.R. Antioxidant and Antibacterial Activities of Myristica Fatua var. Magnifica (beddome) Sinclair. Asian J. Pharm. Clin. Res. 2016, 9, 235–239. [Google Scholar]
- Astani, A.; Reichling, J.; Schnitzler, P. Screening for antiviral activities of isolated compounds from essential oils. Evid. Based Complement. Altern. Med. 2011, 2011, 253643. [Google Scholar] [CrossRef] [PubMed]
- Rasoanaivo, P.; Wright, C.W.; Willcox, M.L.; Gilbert, B. Whole plant extracts versus single compounds for the treatment of malaria: Synergy and positive interactions. Malar. J. 2011, 10, S4. [Google Scholar] [CrossRef]
- Ulrich-Merzenich, G.; Panek, D.; Zeitler, H.; Vetter, H.; Wagner, H. Drug development from natural products: Exploiting synergistic effects. Indian J. Exp. Biol. 2010, 48, 208–219. [Google Scholar]
- Kohn, L.K.; Foglio, M.A.; Rodrigues, R.A.; Sousa, I.D.O.; Martini, M.C.; Padilla, M.A.; de Lima Neto, D.F.; Arns, C.W. In-vitro antiviral activities of extracts of plants of the Brazilian cerrado against the avian metapneumovirus (aMPV). Braz. J. Poult. Sci. 2015, 17, 275–280. [Google Scholar] [CrossRef]
- Prasasty, V.D.; Hanafi, M. Novel Cinchona Alkaloid Derivatives as Potential Antimalarial Agents through Receptor–Inhibitor Interaction Fingerprint and Biosynthesis Design. Orient. J. Chem. 2018, 34, 2643–2650. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methanolic Extract | Cell Viability (%) |
|---|---|
| Acorus calamus | 96.5 |
| Cymbopogon citratus | 98.9 |
| Myristica fatua | 122.7 |
| Sample | Average Infectivity (%) | Average Inhibition (%) |
|---|---|---|
| Acorus calamus (leaves methanol extract) | 73.4 | 26.6 |
| Cymbopogon citratus (root methanol extract) | 47.8 | 52.2 |
| Myristica fatua (methanol extract) | 21.6 | 78.4 |
| Plant Extract | CC50 (µg.mL−1) | EC50 (µg.mL−1) | SI |
|---|---|---|---|
| Acorus calamus | 424.93 | n.d.* | n.d. |
| Cymbopogon citratus | 183.74 | 29.37 | 6.26 |
| Myristica fatua | 474.42 | 25.33 | 18.73 |
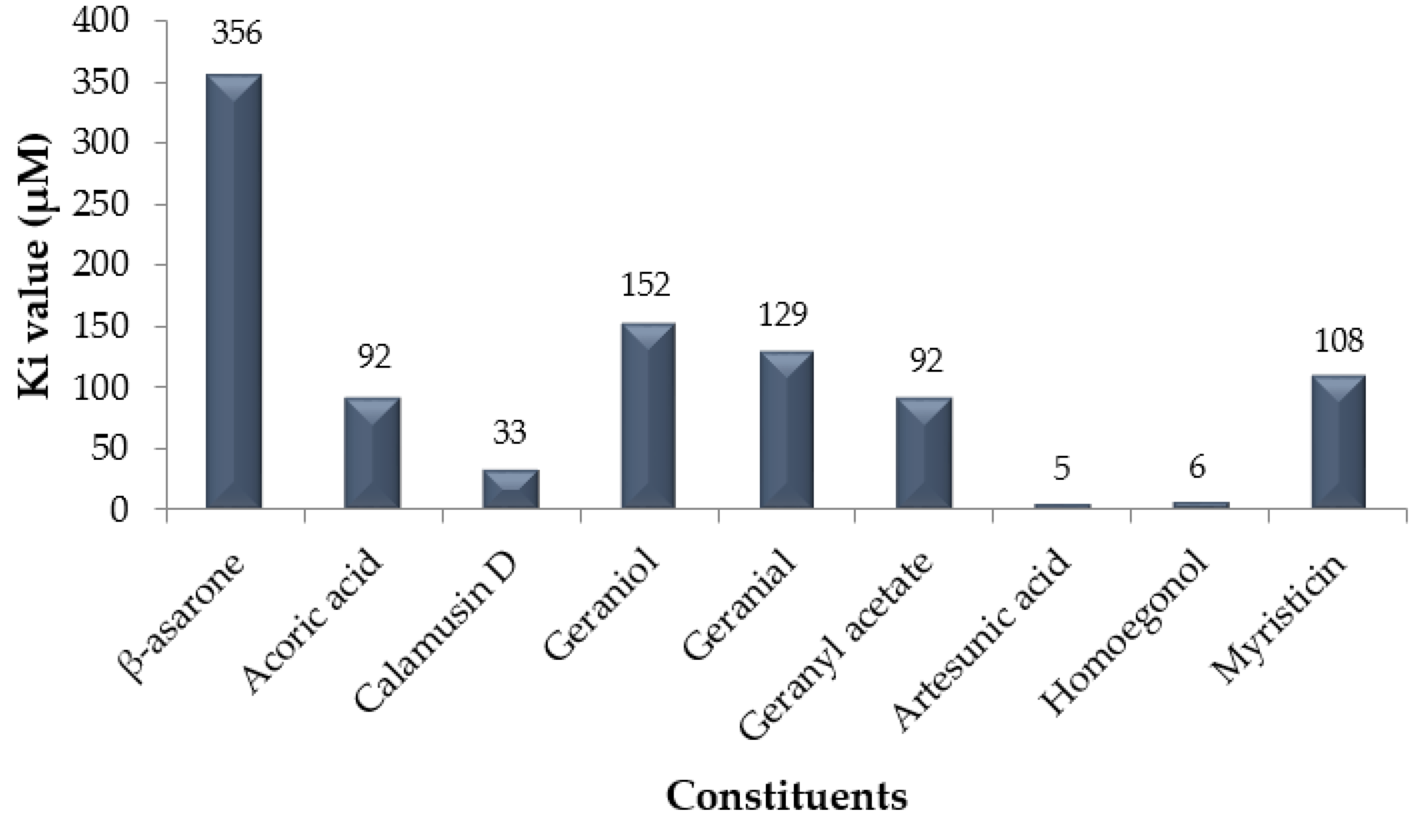
| No. | Constituent | Plant Source | Free energy of Binding (kcal/mol) | References |
|---|---|---|---|---|
| 1 | β-asarone | Acorus calamus | −4.7 | 26 |
| 2 | Acoric acid | Acorus calamus | −5.5 | 26 |
| 3 | Calamusin D | Acorus calamus | −6.1 | 26 |
| 4 | Geraniol | Cymbopogon citratus | −5.2 | 27 |
| 5 | Geranial | Cymbopogon citratus | −5.3 | 28 |
| 6 | Geranyl acetate | Cymbopogon citratus | −5.5 | 28 |
| 7 | Artesunic acid | Myristica fatua | −7.2 | 29 |
| 8 | Homoegonol | Myristica fatua | −7.1 | 29 |
| 9 | Myristicin | Myristica fatua | −5.4 | 29 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosmalena, R.; Elya, B.; Dewi, B.E.; Fithriyah, F.; Desti, H.; Angelina, M.; Hanafi, M.; Lotulung, P.D.; Prasasty, V.D.; Seto, D. The Antiviral Effect of Indonesian Medicinal Plant Extracts Against Dengue Virus In Vitro and In Silico. Pathogens 2019, 8, 85. https://doi.org/10.3390/pathogens8020085
Rosmalena R, Elya B, Dewi BE, Fithriyah F, Desti H, Angelina M, Hanafi M, Lotulung PD, Prasasty VD, Seto D. The Antiviral Effect of Indonesian Medicinal Plant Extracts Against Dengue Virus In Vitro and In Silico. Pathogens. 2019; 8(2):85. https://doi.org/10.3390/pathogens8020085
Chicago/Turabian StyleRosmalena, Rosmalena, Berna Elya, Beti E. Dewi, Fithriyah Fithriyah, Hidayati Desti, Marissa Angelina, Muhammad Hanafi, Puspa D. Lotulung, Vivitri D. Prasasty, and Dimas Seto. 2019. "The Antiviral Effect of Indonesian Medicinal Plant Extracts Against Dengue Virus In Vitro and In Silico" Pathogens 8, no. 2: 85. https://doi.org/10.3390/pathogens8020085
APA StyleRosmalena, R., Elya, B., Dewi, B. E., Fithriyah, F., Desti, H., Angelina, M., Hanafi, M., Lotulung, P. D., Prasasty, V. D., & Seto, D. (2019). The Antiviral Effect of Indonesian Medicinal Plant Extracts Against Dengue Virus In Vitro and In Silico. Pathogens, 8(2), 85. https://doi.org/10.3390/pathogens8020085