Armillaria Root-Rot Pathogens: Species Boundaries and Global Distribution


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Genes and Genomic Regions Employed in Phylogenetic Studies of Armillaria Species
3. Curation of Sequences from GenBank and Phylogenetic Analyses
4. Relatives of Armillaria
5. Species Clusters and Phylogenetic Relationships Based on Morphological Cohesion, Phenotypic Characteristics and Phylogenetic Analyses
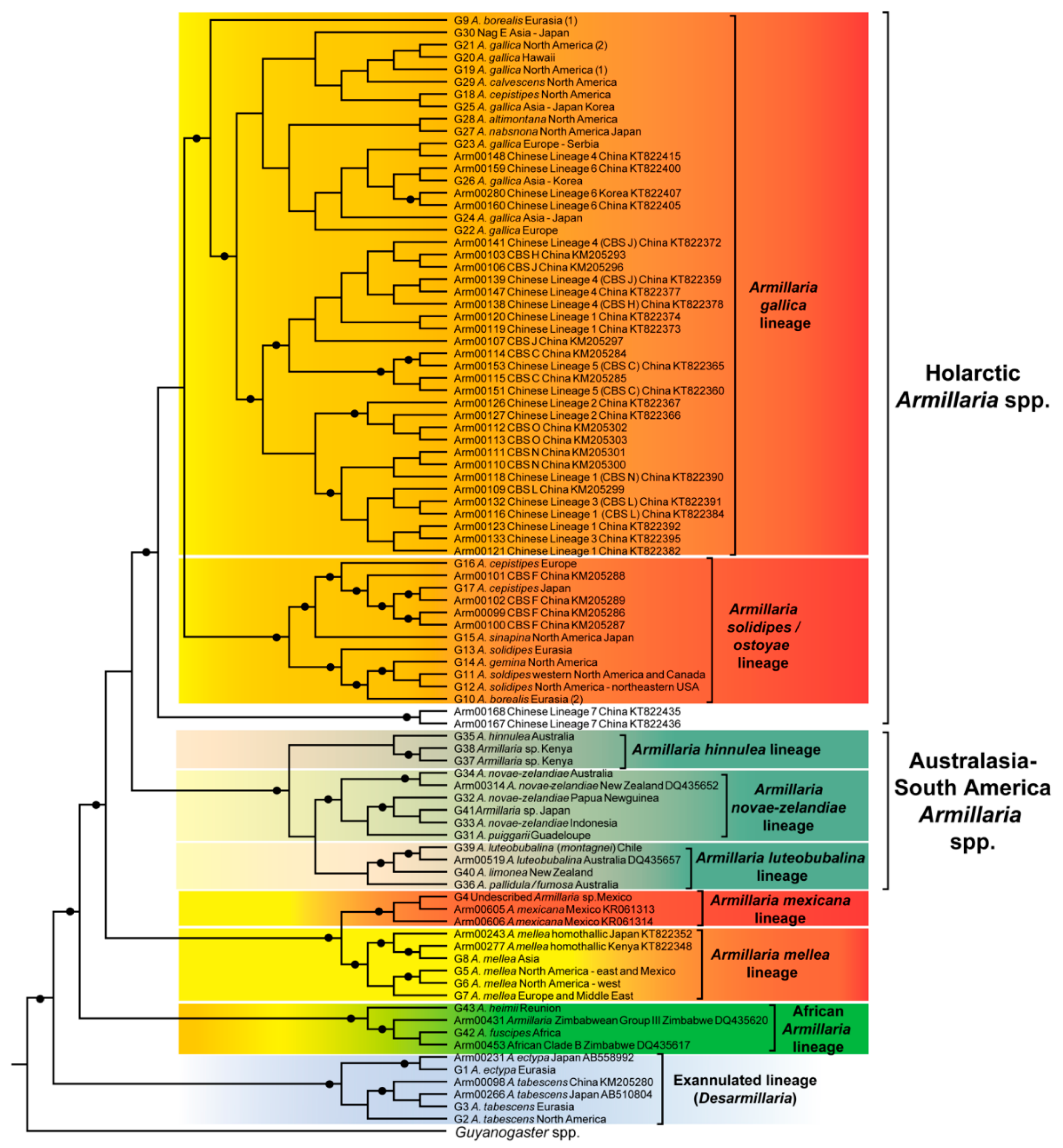
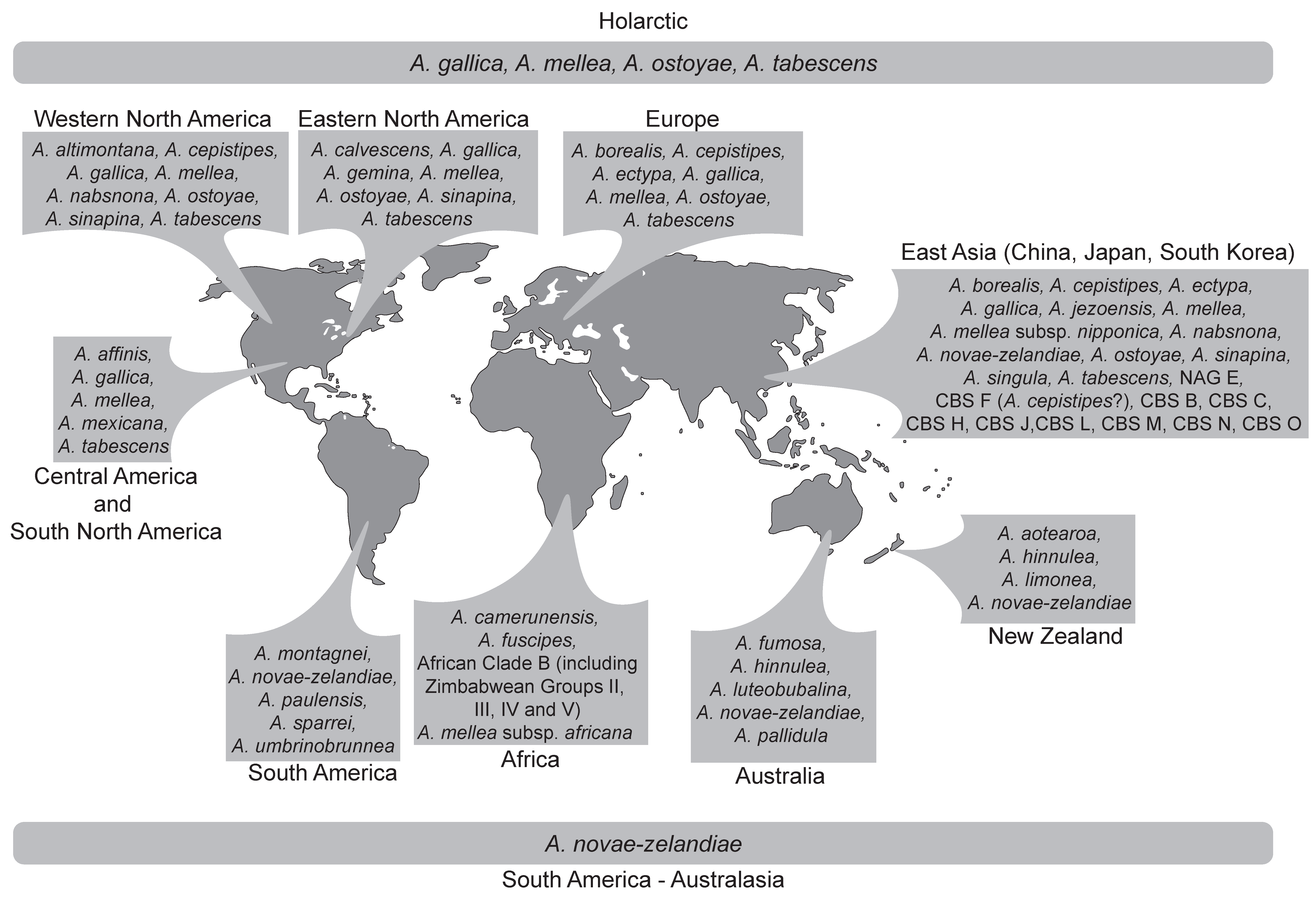
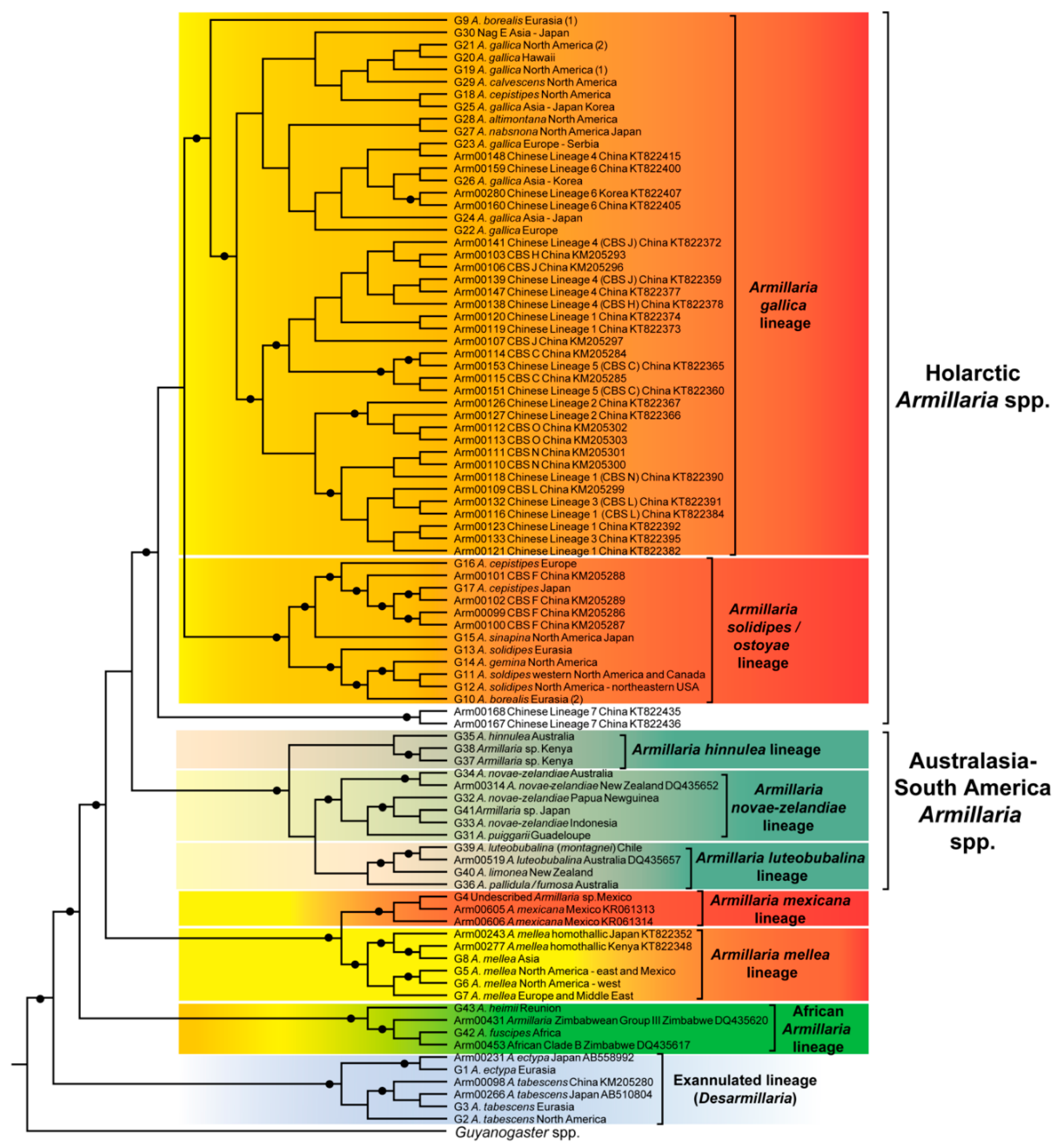
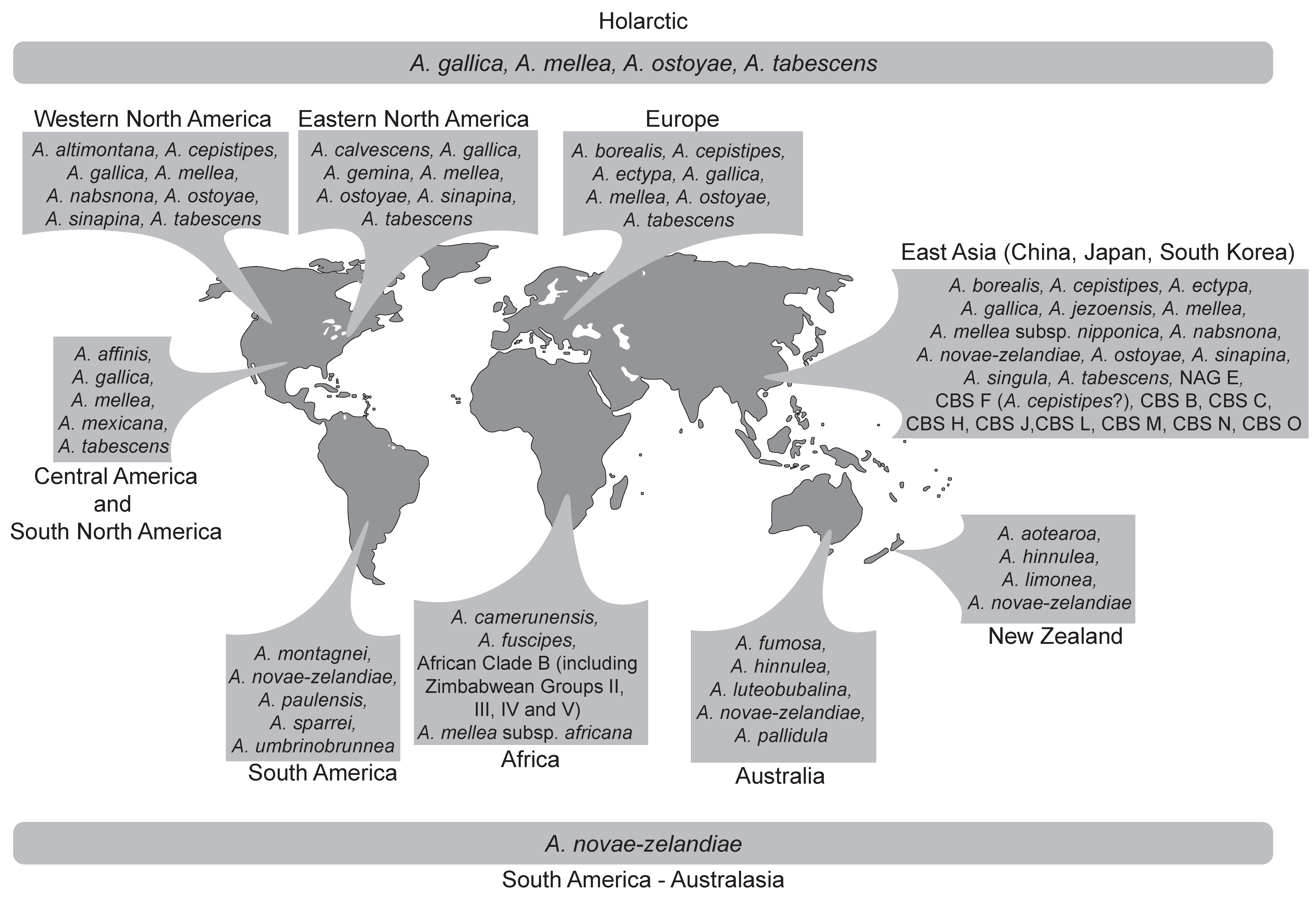
5.1. Species Lineages from the Holarctic
5.1.1. The Armillaria solidipes/ostoyae Lineage
5.1.2. The Armillaria gallica Lineage
5.1.3. The Armillaria mellea Lineage
5.1.4. The Armillaria mexicana Lineage
5.1.5. The Exannulated Lineage of Species
5.2. Species Lineages from Australasia-South America
5.2.1. The Armillaria hinnulea Lineage
5.2.2. The Armillaria novae-zelandiae Lineage
5.2.3. The Armillaria luteobubalina Lineage
5.3. The African Armillaria Lineage
6. Conclusions and Future Prospects
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References and Note
- Shaw, C.G.; Kile, G.A. Armillaria Root Disease. Agriculture Handbook No. 691; Forest Service, United States Department of Agriculture: Washington, DC, USA, 1991.
- Cha, J.Y.; Igarashi, T. Armillaria species associated with Gastrodia elata in Japan. Eur. J. For. Pathol. 1995, 25, 319–326. [Google Scholar] [CrossRef]
- Terashita, T. Biological species of Armillaria symbiotic with Galeola septentrionalis. Nippon Kingakukai Kaiho 1996, 37, 45–49. [Google Scholar]
- Cha, J.Y.; Igarashi, T. Armillaria jezoensis, a new symbiont of Galeola septentrionalis (Orchidaceae) in Hokkaido. Mycoscience 1996, 37, 21–24. [Google Scholar] [CrossRef]
- Sekizaki, H.; Kuninaga, S.; Yamamoto, M.; Asazu, S.N.; Sawa, S.; Kojoma, M.; Yokosawa, R.; Yoshida, N. Identification of Armillaria nabsnona in gastrodia tubers. Biol. Pharm. Bull. 2008, 31, 1410–1414. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Wang, H.C.; Xue, W.Q.; Zhao, J.; Yang, Z.L. Phylogenetic analyses of Armillaria reveal at least 15 phylogenetic lineages in China, seven of which are associated with cultivated Gastrodia elata. PLoS ONE 2016, 11, e0154794. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.X.; Xu, J.T. Nutrient source of sclerotia of Grifola umbellata and its relationship to Armillaria mellea. Acta Bot. Sin. 1992, 34, 576–580. [Google Scholar]
- Kikuchi, G.; Yamaji, H. Identification of Armillaria species associated with Polyporus umbellatus using ITS sequences of nuclear ribosomal DNA. Mycoscience 2010, 51, 366–372. [Google Scholar] [CrossRef]
- Lee, M.W.; Chang, K.C.; Shin, D.B.; Lee, K.R.; Im, K.H.; Jin, G.H.; Shin, P.G.; Xing, Y.M.; Chen, J.; Guo, S.X.; et al. The culture conditions for mycelial growth and sclerotial formation of Polyporus umbellatus. J. Mushroom Sci. Prod. 2013, 11, 194–200. [Google Scholar] [CrossRef]
- Xing, X.; Men, J.; Guo, S. Phylogenetic constrains on Polyporus umbellatus-Armillaria associations. Sci. Rep. 2017, 7, 4226. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.L.; Bruhn, J.N.; Anderson, J.B. The fungus Armillaria bulbosa is among the largest and oldest living organisms. Nature 1992, 356, 428–431. [Google Scholar] [CrossRef]
- Ferguson, B.A.; Dreisbach, T.A.; Parks, C.G.; Filip, G.M.; Schmitt, C.L. Coarse-scales population structure of pathogenic Armillaria species in a mixed-conifer forest in the Blue Mountains of northeast Oregon. Can. J. For. Res. 2003, 33, 612–623. [Google Scholar] [CrossRef]
- Vahl, M. (1787–1799). Flora Danica. Fasc. 16–21, Tab. 901–1260.—(1794). Nogle iagttagelser ved en reise giennem Norge til dets nordlige dele (2). - Skr. Naturhist. Selsk.
- Volk, T.J.; Burdsall, H.H. A Nomenclatural Study of Armillaria and Armillariella Species (Basidiomycotina, Tricholomataceae); Fungiflora: Førde, Norway, 1995; p. 121. [Google Scholar]
- Watling, R.; Kile, G.A.; Gregory, N.M. The genus Armillaria–nomenclature, typification, the identity of Armillaria mellea and species differentiation. Trans. Br. Mycol. Soc. 1982, 78, 271–285. [Google Scholar] [CrossRef]
- Anderson, J.B.; Stasovski, E. Molecular phylogeny of Northern Hemisphere species of Armillaria. Mycologia 1992, 84, 505–516. [Google Scholar] [CrossRef]
- Coetzee, M.P.; Wingfield, B.D.; Bloomer, P.; Wingfield, M.J. Phylogenetic analyses of DNA sequences reveal species partitions amongst isolates of Armillaria from Africa. Mycol. Res. 2005, 109, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Chillali, M.; Wipf, D.; Guillaumin, J.-J.; Mohammed, C.; Botton, B. Delineation of the European Armillaria species based on the sequences of the internal transcribed spacer (ITS) of ribosomal DNA. New Phytol. 1998, 138, 553–561. [Google Scholar] [CrossRef]
- Mulholland, V.; MacAskill, G.A.; Laue, B.E.; Steele, H.; Kenyon, D.; Green, S. Development and verification of a diagnostic assay based on EF-1 a for the identification of Armillaria species in Northern Europe. For. Pathol. 2012, 42, 229–238. [Google Scholar] [CrossRef]
- Coetzee, M.P.A.; Wingfield, B.D.; Bloomer, P.; Ridley, G.S.; Kile, G.A.; Wingfield, M.J. Phylogenetic relationships of Australian and New Zealand Armillaria species. Mycologia 2001, 93, 887–896. [Google Scholar] [CrossRef]
- Dunne, C.P.; Glen, M.; Tommerup, I.C.; Shearer, B.L.; Hardy, G.E.S.J. Sequence variation in the rDNA ITS of Australian Armillaria species and intra-specific variation in A. luteobubalina. Australas. Plant Pathol. 2002, 31, 241–251. [Google Scholar] [CrossRef]
- Coetzee, M.P.A.; Wingfield, B.D.; Bloomer, P.; Ridley, G.S.; Wingfield, M.J. Molecular identification and phylogeny of Armillaria isolates from South America and Indo-Malaysia. Mycologia 2003, 95, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Pildain, M.; Coetzee, M.; Rajchenberg, M.; Petersen, R.; Wingfield, M.; Wingfield, B. Molecular phylogeny of Armillaria from the Patagonian Andes. Mycol. Prog. 2009, 8, 181–194. [Google Scholar] [CrossRef]
- Terashima, K.; Cha, J.Y.; Yajima, T.; Igarashi, T.; Miura, K. Phylogenetic analysis of Japanese Armillaria based on the intergenic spacer (IGS) sequences of their ribosomal DNA. Eur. J. For. Pathol. 1998, 28, 11–19. [Google Scholar] [CrossRef]
- Coetzee, M.P.A.; Wingfield, B.D.; Zhao, J.; van Coller, S.J.; Wingfield, M.J. Phylogenetic relationships among biological species of Armillaria from China. Mycoscience 2015, 56, 530–541. [Google Scholar] [CrossRef]
- Maphosa, L.; Wingfield, B.D.; Coetzee, M.P.A.; Mwenje, E.; Wingfield, M.J. Phylogenetic relationships among Armillaria species inferred from partial elongation factor 1-alpha DNA sequence data. Australas. Plant Pathol. 2006, 35, 513–520. [Google Scholar] [CrossRef]
- Coetzee, M.P.A.; Bloomer, P.; Wingfield, M.J.; Wingfield, B.D. Paleogene radiation of a plant pathogenic mushroom. PLoS ONE 2011, 6, e28545. [Google Scholar] [CrossRef] [PubMed]
- Klopfenstein, N.B.; Stewart, J.E.; Ota, Y.; Hanna, J.W.; Richardson, B.A.; Ross-Davis, A.L.; Elías-Román, R.D.; Korhonen, K.; Keča, N.; Iturritxa, E.; et al. Insights into the phylogeny of Northern Hemisphere Armillaria: Neighbor-net and Bayesian analyses of translation elongation factor 1-α gene sequences. Mycologia 2017, 109, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Koch, R.A.; Wilson, A.W.; Séné, O.; Henkel, T.W.; Aime, M.C. Resolved phylogeny and biogeography of the root pathogen Armillaria and its gasteroid relative, Guyanagaster. BMC Evol. Biol. 2017, 17, 33. [Google Scholar] [CrossRef] [PubMed]
- Morrone, J.J. Biogeographical regionalisation of the world: A reappraisal. Aust. Syst. Bot. 2015, 28, 81–90. [Google Scholar] [CrossRef]
- Kile, G.A.; Guillaumin, J.-J.; Mohammed, C.; Watling, R. Biogeography and pathology of Armillaria. In Proceedings of the Eigth International Conference on Root and Butt Rots, Wik, Sweden; Haikko, Finland, 9–16 August 1993; Swedish University of Agricultural Science: Uppsala, Sweden, 1994; pp. 411–436. [Google Scholar]
- Coetzee, M.P.A.; Wingfield, B.D.; Kirisits, T.; Chhetri, D.B.; Bloomer, P.; Wingfield, M.J. Identification of Armillaria isolates from Bhutan based on DNA sequence comparisons. Plant Pathol. 2005, 54, 36–45. [Google Scholar] [CrossRef]
- Hanna, J.W.; Klopfenstein, N.B.; Kim, M.S.; McDonald, G.I.; Moore, J.A. Phylogeographic patterns of Armillaria ostoyae in the western United States. For. Pathol. 2007, 37, 192–216. [Google Scholar] [CrossRef]
- Coetzee, M.P.A.; Wingfield, B.D.; Coutinho, T.A.; Wingfield, M.J. Identification of the causal agent of Armillaria root rot of Pinus species in South Africa. Mycologia 2000, 92, 777–785. [Google Scholar] [CrossRef]
- Matheny, P.B.; Wang, Z.; Binder, M.; Curtis, J.M.; Lim, Y.W.; Nilsson, R.H.; Hughes, K.W.; Hofstetter, V.; Ammirati, J.F.; Schoch, C.; et al. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogen. Evol. 2007, 43, 430–451. [Google Scholar] [CrossRef] [PubMed]
- Kauserud, H.; Schumacher, T. Outcrossing or inbreeding: DNA markers provide evidence for type of reproductive mode in Phellinus nigrolimitatus (Basidiomycota). Mycol. Res. 2001, 105, 676–683. [Google Scholar] [CrossRef]
- Baumgartner, K.; Bhat, R.; Fujiyoshi, P. A rapid infection assay for Armillaria and real-time PCR quantitation of the fungal biomass in planta. Fungal Biol 2010, 114, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, E.; Ota, Y.; Hattori, T.; Kikuchi, T. Sequence-based identification of Japanese Armillaria species using the elongation factor-1 alpha gene. Mycologia 2010, 102, 890–910. [Google Scholar] [CrossRef]
- Brazee, N.J.; Hulvey, J.P.; Wick, R.L. Evaluation of partial tef1, rpb2, and nLSU sequences for identification of isolates representing Armillaria calvescens and Armillaria gallica from northeastern North America. Fungal Biol. 2011, 115, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Ross-Davis, A.L.; Hanna, J.W.; Klopfenstein, N.B.; Kim, M.-S. Advances toward DNA-based identification and phylogeny of North American Armillaria species using elongation factor-1 alpha gene. Mycoscience 2012, 53, 161–165. [Google Scholar] [CrossRef]
- Burdsall, H.H.; Volk, T.J. The state of taxonomy of the genus Armillaria. McIlvainea 1993, 11, 4–12. [Google Scholar]
- Zhao, Z.; Liu, H.; Luo, Y.; Zhou, S.; An, L.; Wang, C.; Jin, Q.; Zhou, M.; Xu, J.-R. Molecular evolution and functional divergence of tubulin superfamily in the fungal tree of life. Sci. Rep. 2014, 4, 6746. [Google Scholar] [CrossRef] [PubMed]
- Piercey-Normore, M.D.; Egger, K.N.; Bérubé, J.A. Molecular phylogeny and evolutionary divergence of North American Biological Species of Armillaria. Mol. Phylogen. Evol. 1998, 10, 49–66. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, bbx108. [Google Scholar] [CrossRef] [PubMed]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.5. Available online: http://www.mesquiteproject.org (accessed on 21 October 2018).
- Moncalvo, J.-M.; Lutzoni, F.M.; Rhener, S.A.; Johnson, J.; Vilgalys, R. Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences. Syst. Biol. 2000, 49, 278–305. [Google Scholar] [CrossRef] [PubMed]
- Moncalvo, J.-M.; Vilgalys, R.; Redhead, S.A.; Johnson, J.E.; James, T.Y.; Catherine Aime, M.; Hofstetter, V.; Verduin, S.J.W.; Larsson, E.; Baroni, T.J.; et al. One hundred and seventeen clades of euagarics. Mol. Phylogen. Evol. 2002, 23, 357–400. [Google Scholar] [CrossRef]
- Matheny, P.B.; Curtis, J.M.; Hofstetter, V.; Aime, M.C.; Moncalvo, J.-M.; Ge, Z.-W.; Yang, Z.-L.; Slot, J.C.; Ammirati, J.F.; Baroni, T.J.; et al. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 982–995. [Google Scholar] [CrossRef] [PubMed]
- Garnica, S.; Weiss, M.; Walther, G.; Oberwinkler, F. Reconstructing the evolution of agarics from nuclear gene sequences and basidiospore ultrastructure. Mycol. Res. 2007, 111, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Henkel, T.W.; Smith, M.E.; Aime, M.C. Guyanagaster, a new wood-decaying sequestrate fungal genus related to Armillaria (Physalacriaceae, Agaricales, Basidiomycota). Am. J. Bot. 2010, 97, 1471–1484. [Google Scholar] [CrossRef] [PubMed]
- Antonín, V.; Jankovský, L.; Lochman, J.; Tomšovský, M. Armillaria socialis morphological anatomical and ecological characteristics, pathology, distribution in the Czech Republic and Europe and remarks on its genetic variation. Czech Mycol. 2006, 58, 209–224. [Google Scholar]
- Pegler, D.N. Taxonomy, nomenclature and description of Armillaria. In Armillaria Root Rot: Biology and Control of Honey Fungus; Fox, R.T.V., Ed.; Intercept Limited: Andover, UK, 2000; pp. 81–93. [Google Scholar]
- Zolciak, A.; Bouteville, R.J.; Tourvieille, J.; Roeckel-Drevet, P.; Nicolas, P.; Guillaumin, J.J. Occurrence of Armillaria ectypa (Fr.) Lamoure in peat bogs of the Auvergne—The reproduction system of the species. Cryptogam. Mycol. 1997, 18, 299–313. [Google Scholar]
- Korhonen, K. Armillaria since Elias Fries. Acta Universitatis Upsaliensis Symbolae Botanicae Upsalienses 1995, 30, 153–161. [Google Scholar]
- Burdsall, H.H.; Volk, T.J. Armillaria solidipes, an older name for the fungus called Armillaria ostoyae. N. Am. Fungi 2008, 3, 261–267. [Google Scholar] [CrossRef]
- Hunt, R.S.; Morrison, D.J.; Bérubé, J. Armillaria solidipes is not a replacement name for A. ostoyae. For. Pathol. 2011, 41, 253–254. [Google Scholar] [CrossRef]
- Guillaumin, J.J.; Mohammed, C.; Anselmi, N.; Courtecuisse, R.; Gregory, S.C.; Holdenrieder, O.; Intini, M.; Lung, B.; Marxmuller, H.; Morrison, D.; et al. Geographical distribution and ecology of the Armillaria species in western Europe. Eur. J. For. Pathol. 1993, 23, 321–341. [Google Scholar] [CrossRef]
- Qin, G.F.; Zhao, J.; Korhonen, K. A study on intersterility groups of Armillaria in China. Mycologia 2007, 99, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Oh, S.Y.; Park, M.S.; Kim, M.S.; Klopfenstein, N.B.; Kim, N.K.; Park, J.Y.; Kim, J.J.; Han, S.K.; Lee, J.K.; et al. Re-evaluation of Armillaria and Desarmillaria in South Korea based on ITS/tef1 sequences and morphological characteristics. For. Pathol. 2018, e12447. [Google Scholar] [CrossRef]
- Anderson, J.B.; Bailey, S.S.; Pukkila, P.J. Variation in ribosomal DNA among biological species of Armillaria, a genus of root-infecting fungi. Evolution 1989, 43, 1652–1662. [Google Scholar] [PubMed]
- Antonín, V.; Tomšovský, M.; Sedlák, P.; Májek, T.; Jankovský, L. Morphological and molecular characterization of the Armillaria cepistipes—A. gallica complex in the Czech Republic and Slovakia. Mycol. Prog. 2009, 8, 259–271. [Google Scholar] [CrossRef]
- Kim, M.S.; Klopfenstein, N.B.; Hanna, J.W.; McDonald, G.I. Characterization of North American Armillaria species: Genetic relationships determined by ribosomal DNA sequences and AFLP markers. For. Pathol. 2006, 36, 145–164. [Google Scholar] [CrossRef]
- Tsykun, T.; Rigling, D.; Prospero, S. A new multilocus approach for a reliable DNA-based identification of Armillaria species. Mycologia 2013, 105, 1059–1076. [Google Scholar] [CrossRef] [PubMed]
- Elías-Román, R.D.; Guzmán-Plazola, R.A.; Klopfenstein, N.B.; Alvarado-Rosales, D.; Calderón-Zavala, G.; Mora-Aguilera, J.A.; Kim, M.S.; García-Espinosa, R. Incidence and phylogenetic analyses of Armillaria spp. associated with root disease in peach orchards in the State of Mexico, Mexico. For. Pathol. 2013, 43, 390–401. [Google Scholar] [CrossRef]
- Keča, N.; Klopfenstein, N.B.; Kim, M.S.; Solheim, H.; Woodward, S. Initial characterization of an unidentified Armillaria isolate from Serbia using LSU-IGS1 and TEF-1-α genes. For. Pathol. 2015, 45, 120–126. [Google Scholar] [CrossRef]
- Motta, J.J.; Korhonen, K. A note on Armilaria mellea and Armillaria bulbosa from the Middle Atlantic States. Mycologia 1986, 78, 471–474. [Google Scholar] [CrossRef]
- Elias-Roman, R.D.; Guzman-Plazola, R.A.; Alvarado-Rosales, D.; Calderon-Zavala, G.; Mora-Aguilera, J.A.; Garcia-Espinosa, R.; Kim, M.S.; Ross-Davis, A.L.; Hanna, J.W.; Klopfenstein, N.B. Armillaria root disease in peach orchards of the state of Mexico, Mexico: Characterization of Armillaria species and assessment of disease impact. Phytopathology 2013, 103, 39. [Google Scholar]
- Baumgartner, K.; Travadon, R.; Bruhn, J.; Bergemann, S.E. Contrasting patterns of genetic diversity and population structure of Armillaria mellea sensu stricto in the eastern and western United States. Phytopathology 2010, 100, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Asef, M.R.; Mohammadi Goltapeh, E.; Alizadeh, A. Identification of Armillaria biological species in Iran. Fungal Divers. 2003, 14, 51–60. [Google Scholar]
- Ota, Y.; Matsushita, N.; Nagasawa, E.; Terashita, T.; Fukuda, K.; Suzuki, K. Biological species of Armillaria in Japan. Plant Dis. 1998, 82, 537–543. [Google Scholar] [CrossRef]
- Coetzee, M.P.A.; Musasira, N.Y.; Roux, J.; Roets, F.; van der Merwe, N.A.; Wingfield, M.J. Armillaria root rot spreading into a natural woody ecosystem in South Africa. Plant Pathol. 2018, 67, 883–891. [Google Scholar] [CrossRef]
- Guillaumin, J.-J.; Mohammed, C.; Abomo-Ndongo, S. Vegetative incompatibility and sexual systems of Armillaria isolates from tropical Africa. In Proceedings of the Eighth International Conference on Root and Butt Rots, Wik, Sweden; Haikko, Finland, 9–16 August 1993; Swedish University of Agricultural Science: Uppsala, Sweden, 1994; pp. 349–354. [Google Scholar]
- Mohammed, C.; Guillaumin, J.-J.; Botton, B.; Intini, M. Species of Armillaria in tropical Africa. In Proceedings of the Eight International Conference on Root and Butt Rots, Wik, Sweden; Haikko, Finland, 9–16 August 1993; Swedish University of Agricultural Science: Uppsala, Sweden, 1994; pp. 402–410. [Google Scholar]
- Abomo-Ndongo, S.; Mohammed, C.; Guillaumin, J.-J. Sexual behaviour of Armillaria heimii and A. mellea isolates from Africa. Eur. J. For. Pathol. 1997, 27, 207–224. [Google Scholar] [CrossRef]
- Coetzee, M.P.A.; Wingfield, B.D.; Harrington, T.C.; Steimel, J.; Coutinho, T.A.; Wingfield, M.J. The root rot fungus Armillaria mellea introduced into South Africa by early Dutch settlers. Mol. Ecol. 2001, 10, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, M.P.A.; Wingfield, B.D.; Roux, J.; Crous, P.W.; Denman, S.; Wingfield, M.J. Discovery of two northern hemisphere Armillaria species on Proteaceae in South Africa. Plant Pathol. 2003, 52, 604–612. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Coetzee, M.P.A.; Crous, P.W.; Six, D.; Wingfield, B.D. Fungal phoenix rising from the ashes? IMA Fungus 2011, 1, 149–153. [Google Scholar] [CrossRef]
- Coetzee, M.P.A.; Wingfield, B.D.; Harrington, T.C.; Dalevi, D.; Coutinho, T.A.; Wingfield, M.J. Geographical diversity of Armillaria mellea s. s. based on phylogenetic analysis. Mycologia 2000, 92, 105–113. [Google Scholar] [CrossRef]
- Ota, Y.; Fukuda, K.; Suzuki, K. The nonheterothallic life cycle of Japanese Armillaria mellea. Mycologia 1998, 90, 396–405. [Google Scholar] [CrossRef]
- Ota, Y.; Intini, M.; Hattori, T. Genetic characterization of heterothallic and non-heterothallic Armillaria mellea sensu stricto. Mycol. Res. 2000, 104, 1046–1054. [Google Scholar] [CrossRef]
- Cha, J.Y.; Igarashi, T. A note on Armillaria mellea subsp. nipponica subsp. nov. in Japan. Mycoscience 1995, 36, 143–146. [Google Scholar] [CrossRef]
- Elías-Román, R.D.; Medel-Ortiz, R.; Alvarado-Rosales, D.; Hanna, J.W.; Ross-Davis, A.L.; Kim, M.-S.; Klopfenstein, N.B. Armillaria mexicana, a newly described species from Mexico. Mycologia 2018, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Stasińska, M. Armillaria ectypa, a rare fungus of mire in Poland. Acta Mycol. 2015, 50, 1–6. [Google Scholar] [CrossRef]
- Sesli, E.; Denchev, C.M. Checklists of the myxomycetes, larger ascomycetes, and larger basidiomycetes in Turkey. Mycotaxon 2008, 106, 65–67. [Google Scholar]
- Zhang, M. The Fungi in the Region of Hengduan Mountains; Science Press: Beijing, China, 1996. [Google Scholar]
- Kudo, S.; Nagasawa, E. Armillaria ectypa rediscovered in Aomori Prefecture, northern Japan. Rep. Tottori Mycol. Inst. 2003, 41, 26–34, (In Japanese with English Abstract). [Google Scholar]
- Ito, S. Occurrence of Armillaria ectypa in Aomori prefecture. Newsl. Mycol. Soc. Jpn. 2004, 4, 4–5. (In Japanese) [Google Scholar]
- Bruhn, J.N.; Wetteroff, J.J., Jr.; Mihail, J.D.; Kabrick, J.M.; Pickens, J.B. Distribution of Armillaria species in upland Ozark Mountain forests with respect to site, overstory species composition and oak decline. For. Pathol. 2000, 30, 43–60. [Google Scholar] [CrossRef]
- Schnabel, G.; Ash, J.S.; Bryson, P.K. Identification and characterization of Armillaria tabescens from the southeastern United States. Mycol. Res. 2005, 109, 1208–1222. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Klopfenstein, N.B.; Hanna, J.W.; Cannon, P.; Medel, R.; López, A. First report of armillaria root disease caused by Armillaria tabescens on Araucaria araucana in Veracruz, Mexico. Plant Dis. 2010, 94, 784. [Google Scholar] [CrossRef]
- Cha, J.Y.; Lee, S.Y.; Chun, K.W.; Lee, S.Y.; Ohga, S. Armillaria root rot caused by Armillaria tabescens on Prunus salicina in a Korean garden. J. Fac. Agric. Kyushu Univ. 2009, 54, 273–277. [Google Scholar]
- Lee, S.K.; Seo, S.T. First report of Armillaria root disease caused by Armillaria tabescens on Carpinus tschonoskii in South Korea. Plant Dis. 2016, 100, 213. [Google Scholar] [CrossRef]
- Dai, Y.C.; Cui, B.K.; Yuan, H.S.; Li, B.D. Pathogenic wood-decaying fungi in China. For. Pathol. 2007, 37, 105–120. [Google Scholar] [CrossRef]
- Ota, Y.; Kim, M.-S.; Neda, H.; Klopfenstein, N.B.; Hasegawa, E. The phylogenetic position of an Armillaria species from Amami-Oshima, a subtropical island of Japan, based on elongation factor and ITS sequences. Mycoscience 2011, 52, 53–58. [Google Scholar] [CrossRef]
- Darmono, T.W.; Burdsall, H.H.; Volk, T.J. Interfertility among isolates of Armillaria tabescens in North America. Sydowia 1992, 42, 105–116. [Google Scholar]
- Harrington, T.C.; Wingfield, B.D. A PCR-based identification method for species of Armillaria. Mycologia 1995, 87, 280–288. [Google Scholar] [CrossRef]
- Kile, G.A.; Watling, R. Armillaria species from south-eastern Australia. Trans. Br. Mycol. Soc. 1983, 81, 129–140. [Google Scholar] [CrossRef]
- Ramsfield, T.D.; Power, M.W.P.; Ridley, O.S. A comparison of populations of Armillaria hinnulea in New Zealand and Australia. N. Z. Plant Prot. 2008, 61, 41–47. [Google Scholar]
- Hood, I.A.; Ramsfield, T.D. Armillaria aotearoa species nova. N. Z. J. For. Sci. 2016, 46, 1–11. [Google Scholar] [CrossRef]
- Pildain, M.B.; Coetzee, M.P.A.; Wingfield, B.D.; Wingfield, M.J.; Rajchenberg, M. Taxonomy of Armillaria in the Patagonian forests of Argentina. Mycologia 2010, 102, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Guillaumin, J.-J.; Mohammed, C.; Berthelay, S. Armillaria novae-zelandiae in Papua New Guinea. Mycol. Res. 1992, 96, 278–280. [Google Scholar] [CrossRef]
- Stevenson, G. The Agaricales of New Zealand V. Tricholomataceae. Kew Bull. 1964, 19, 1–59. [Google Scholar] [CrossRef]
- Kile, G.A.; Watling, R. Identification and occurrence of Australian Armillaria species, including A. pallidula sp. nov. and comparative studies between them and non-Australian tropical and Indian Armillaria. Trans. Br. Mycol. Soc. 1988, 91, 305–315. [Google Scholar] [CrossRef]
- Singer, R. Mycoflora Australis. Beihefte zur Nova Hedwigia 1969, 29, 40–49. [Google Scholar]
- Horak, E. Flora criptogámica de Tierra del Fuego. Fungi: Basidiomycetes Agaricales y Gasteromycetes Secotioides; CONICET-FECIC: Buenos Aires, Argentina, 1979.
- Boesewinkel, H.J. New plant disease records in New Zealand: Records in the period 1969–1976. N. Z. J. Agric. Res. 1977, 20, 583–589. [Google Scholar] [CrossRef]
- Lima, M.L.A.; Asai, T.; Capelari, M. Armillaria paulensis: A new South American species. Mycol. Res. 2008, 112, 1122–1128. [Google Scholar] [CrossRef] [PubMed]
- Singer, R. The Armillariella mellea group. Lloydia 1956, 19, 176–187. [Google Scholar]
- Podger, F.D.; Kile, G.A.; Watling, R.; Fryer, J. Spread and effects of Armillaria luteobubalina sp. nov. in an Australian Eucaluptus regnans plantation. Trans. Br. Mycol. Soc. 1978, 71, 77–87. [Google Scholar] [CrossRef]
- Pérez-Sierra, A.; Guillaumin, J.-J.; Spooner, B.M.; Bridge, P.D. Characterization of Armillaria heimii from Africa. Plant Pathol. 2004, 53, 220–230. [Google Scholar] [CrossRef]
- Abomo-Ndongo, S.; Tourvieille, J.; Guillaumin, J.J. The Buller phenomenon in Armillaria heimii Pegler, a bipolar diploid basidiomycete. Cryptogam. Mycol. 2002, 23, 335–347. [Google Scholar]
- Guillaumin, J.-J.; Legrand, P. Armillaria root rots. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CABI Publishing: Oxfordshire, UK, 2013; pp. 159–177. [Google Scholar]
- Gezahgne, A.; Coetzee, M.P.A.; Wingfield, B.D.; Wingfield, M.J.; Roux, J. Identification of the Armillaria root rot pathogen in Ethiopian plantations. For. Pathol. 2004, 34, 133–145. [Google Scholar] [CrossRef]
- Mwenje, E.; Wingfield, B.D.; Coetzee, M.P.; Wingfield, M.J. Molecular characterisation of Armillaria species from Zimbabwe. Mycol. Res. 2003, 107, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, B.D.; Maphosa, L.; Coetzee, M.P.A.; Mwenje, E.; Wingfield, M.J. Characterisation of Zimbabwean Armillaria using IGS-1 sequences and AFLP analysis. Fungal Divers. 2009, 34, 187–196. [Google Scholar]
- Mwenje, E.; Wingfield, B.D.; Coetzee, M.P.A.; Nemato, H.; Wingfield, M.J. Armillaria species on tea in Kenya identified using isozyme and DNA sequence comparisons. Plant Pathol. 2006, 55, 343–350. [Google Scholar] [CrossRef]
- Mwangi, L.M.; Lin, D.; Hubbes, M. Identification of Kenyan Armillaria isolates by cultural morphology intersterility tests and analysis of isozyme profiles. Eur. J. For. Pathol. 1989, 19, 399–406. [Google Scholar] [CrossRef]
- Agustian, A.; Mohammed, C.; Guillaumin, J.-J.; Botton, B. Discrimination of some African Armillaria species by isozyme electrophoretic analysis. New Phytol. 1994, 128, 135–143. [Google Scholar] [CrossRef]
- Otieno, W.; Pérez Sierra, A.; Termorshuizen, A. Characterization of Armillaria isolates from tea (Camellia sinensis) in Kenya. Mycologia 2003, 95, 160–175. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Dettman, J.R.; Jacobson, D.J.; Taylor, J.W. A multilocus genealogical approach to phylogenetic species recognition in the model eukaryote Neurospora. Evolution 2003, 57, 2703–2720. [Google Scholar] [CrossRef] [PubMed]
- Rokas, A.; Williams, B.L.; King, N.; Carroll, S.B. Genome-scale approaches to resolving incongruence in molecular phylogenies. Nature 2003, 425, 798–804. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Luo, J.; Bhattacharya, D. Chapter Eight—Advances in fungal phylogenomics and their impact on fungal systematics. In Advances in Genetics; Townsend, J.P., Wang, Z., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 100, pp. 309–328. [Google Scholar]
- Wetterstrand, K. DNA Sequencing Costs: Data from the NHGRI Genome Sequencing Program (GSP). Available online: http//:www.genome.gov/sequencingcostsdata (accessed on 21 October 2018).
- Lemmon, E.M.; Lemmon, A.R. High-throughput genomic data in systematics and phylogenetics. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 99–121. [Google Scholar] [CrossRef]
- Sipos, G.; Prasanna, A.N.; Walter, M.C.; O’Connor, E.; Bálint, B.; Krizsán, K.; Kiss, B.; Hess, J.; Varga, T.; Slot, J.; et al. Genome expansion and lineage-specific genetic innovations in the forest pathogenic fungi Armillaria. Nat. Ecol. Evol. 2017, 1, 1931–1941. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coetzee, M.P.A.; Wingfield, B.D.; Wingfield, M.J. Armillaria Root-Rot Pathogens: Species Boundaries and Global Distribution. Pathogens 2018, 7, 83. https://doi.org/10.3390/pathogens7040083
Coetzee MPA, Wingfield BD, Wingfield MJ. Armillaria Root-Rot Pathogens: Species Boundaries and Global Distribution. Pathogens. 2018; 7(4):83. https://doi.org/10.3390/pathogens7040083
Chicago/Turabian StyleCoetzee, Martin P.A., Brenda D. Wingfield, and Michael J. Wingfield. 2018. "Armillaria Root-Rot Pathogens: Species Boundaries and Global Distribution" Pathogens 7, no. 4: 83. https://doi.org/10.3390/pathogens7040083
APA StyleCoetzee, M. P. A., Wingfield, B. D., & Wingfield, M. J. (2018). Armillaria Root-Rot Pathogens: Species Boundaries and Global Distribution. Pathogens, 7(4), 83. https://doi.org/10.3390/pathogens7040083