Limited Interactions between Streptococcus Suis and Haemophilus Parasuis in In Vitro Co-Infection Studies



Abstract
:1. Introduction
2. Results and Discussion
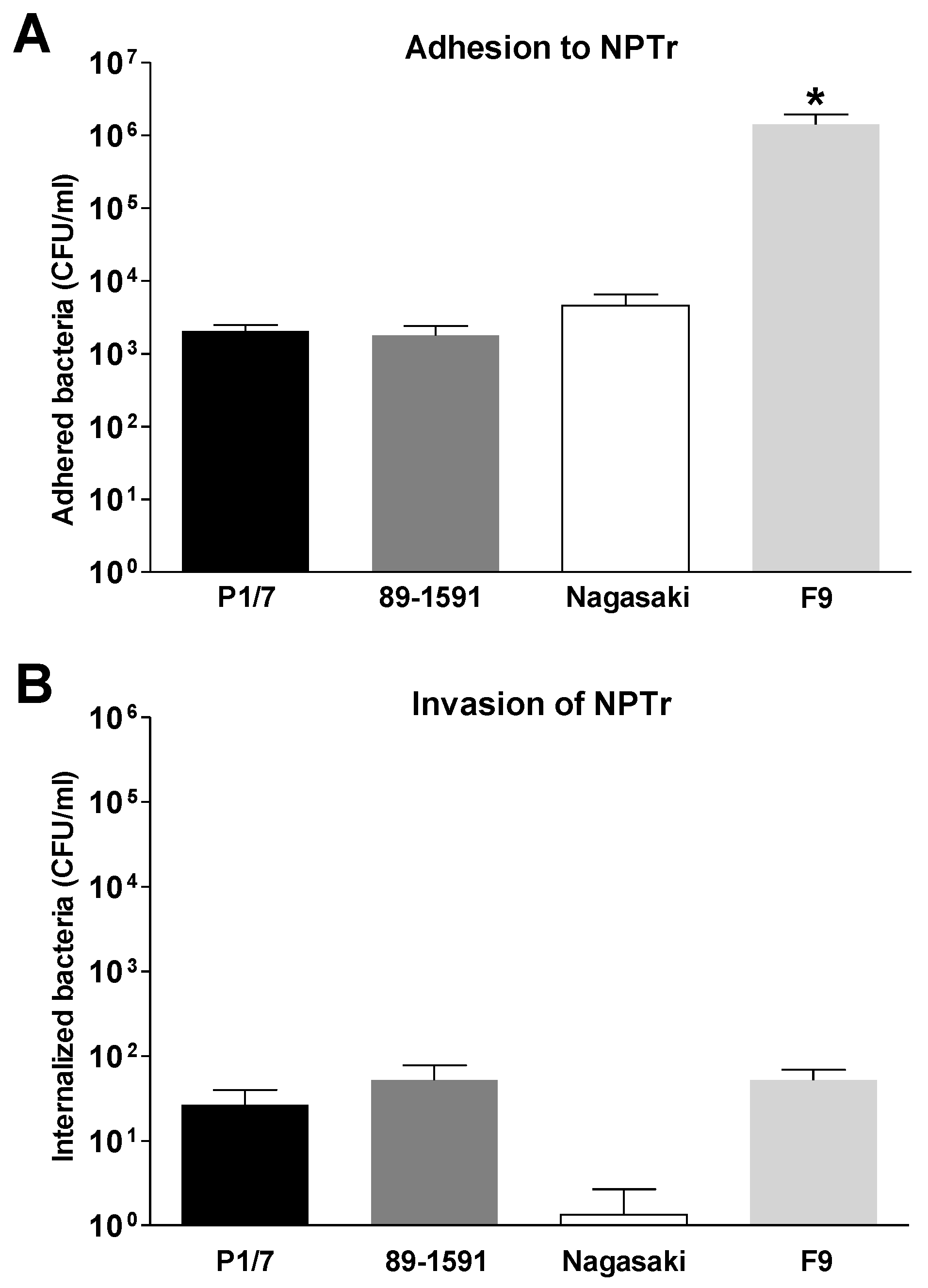
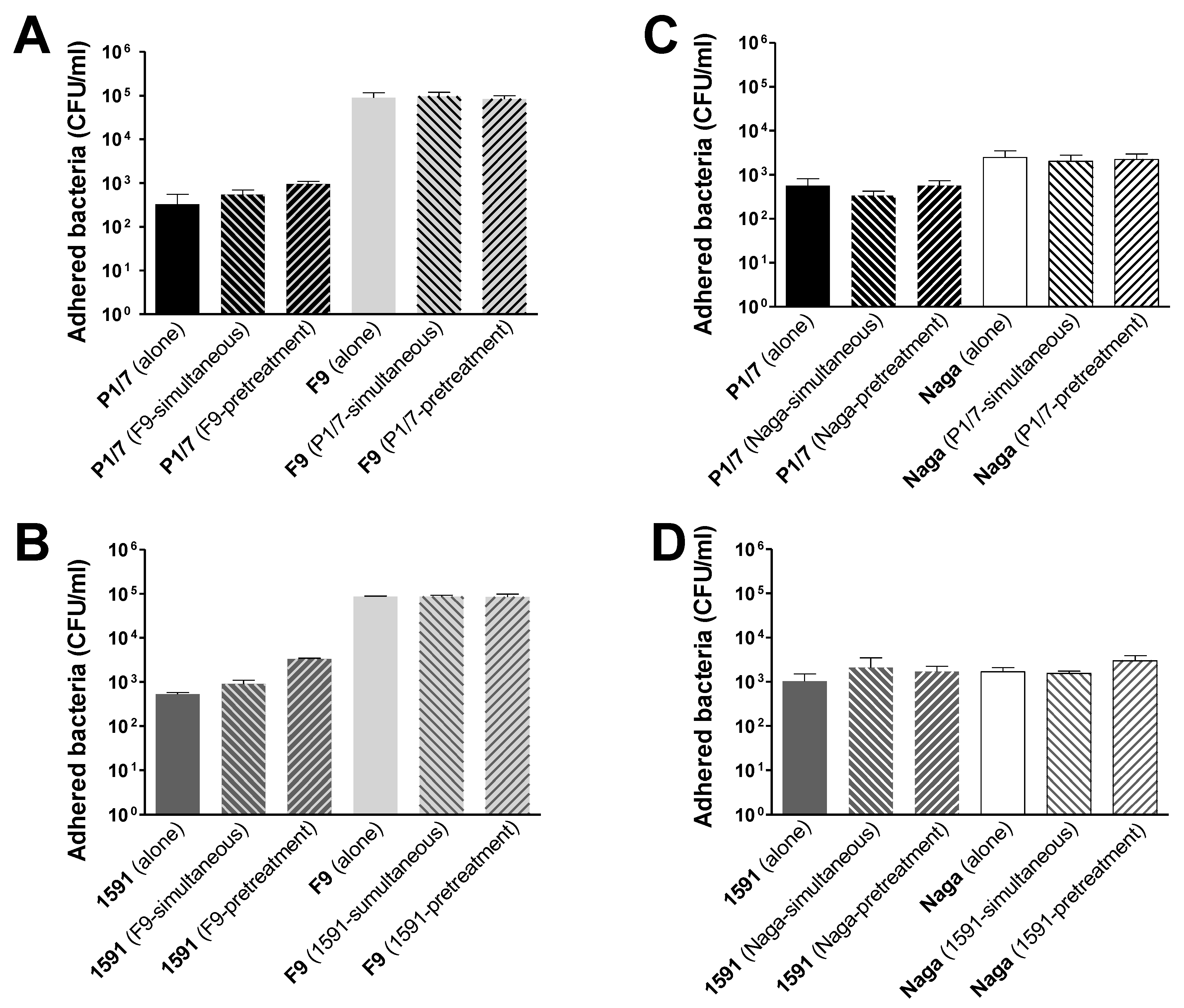
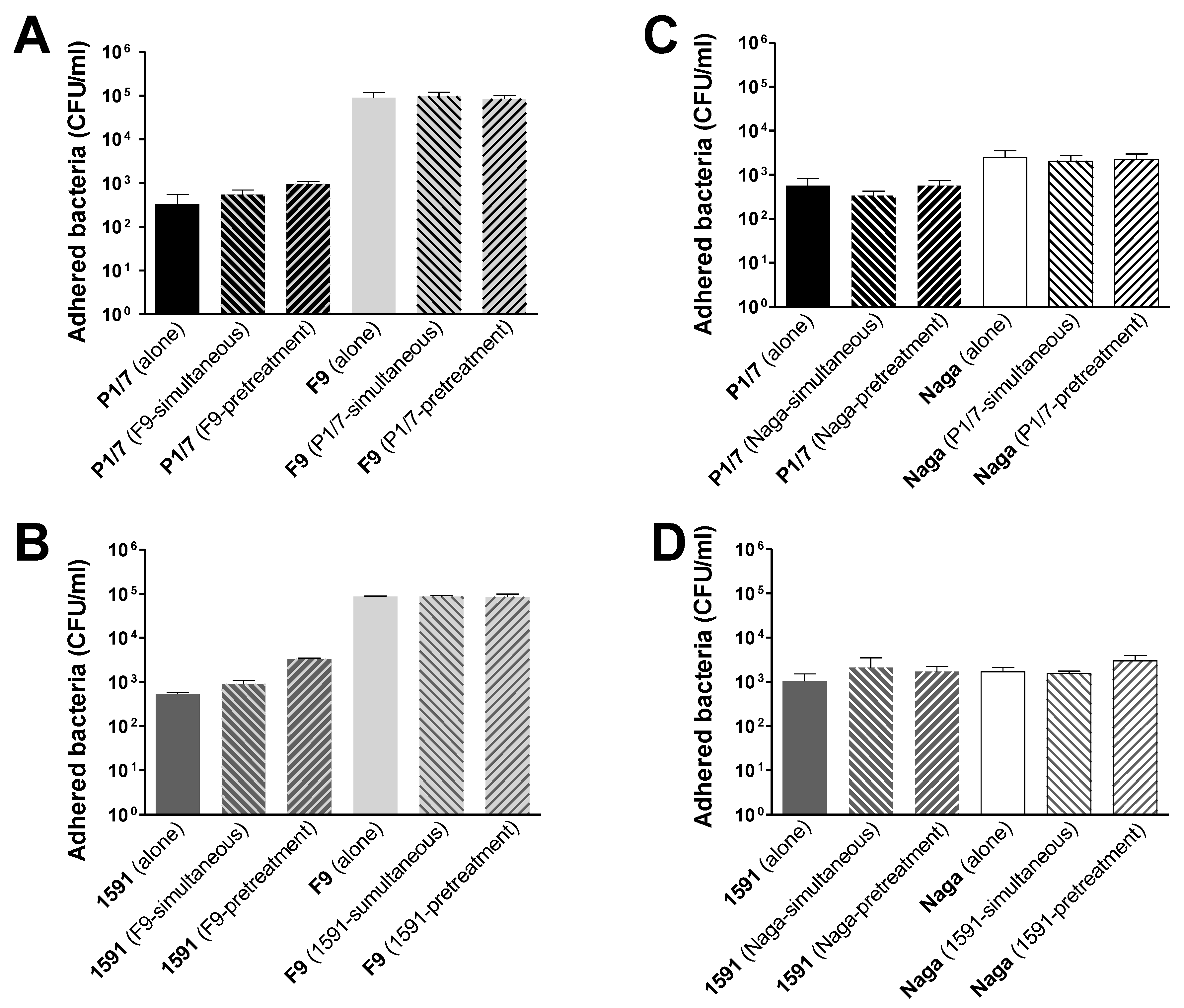
2.1. Co- or Sequential Infections of Swine Tracheal Epithelial Cells by S. suis and H. parasuis Has No Impact on Their Adhesion/Invasion Capacities
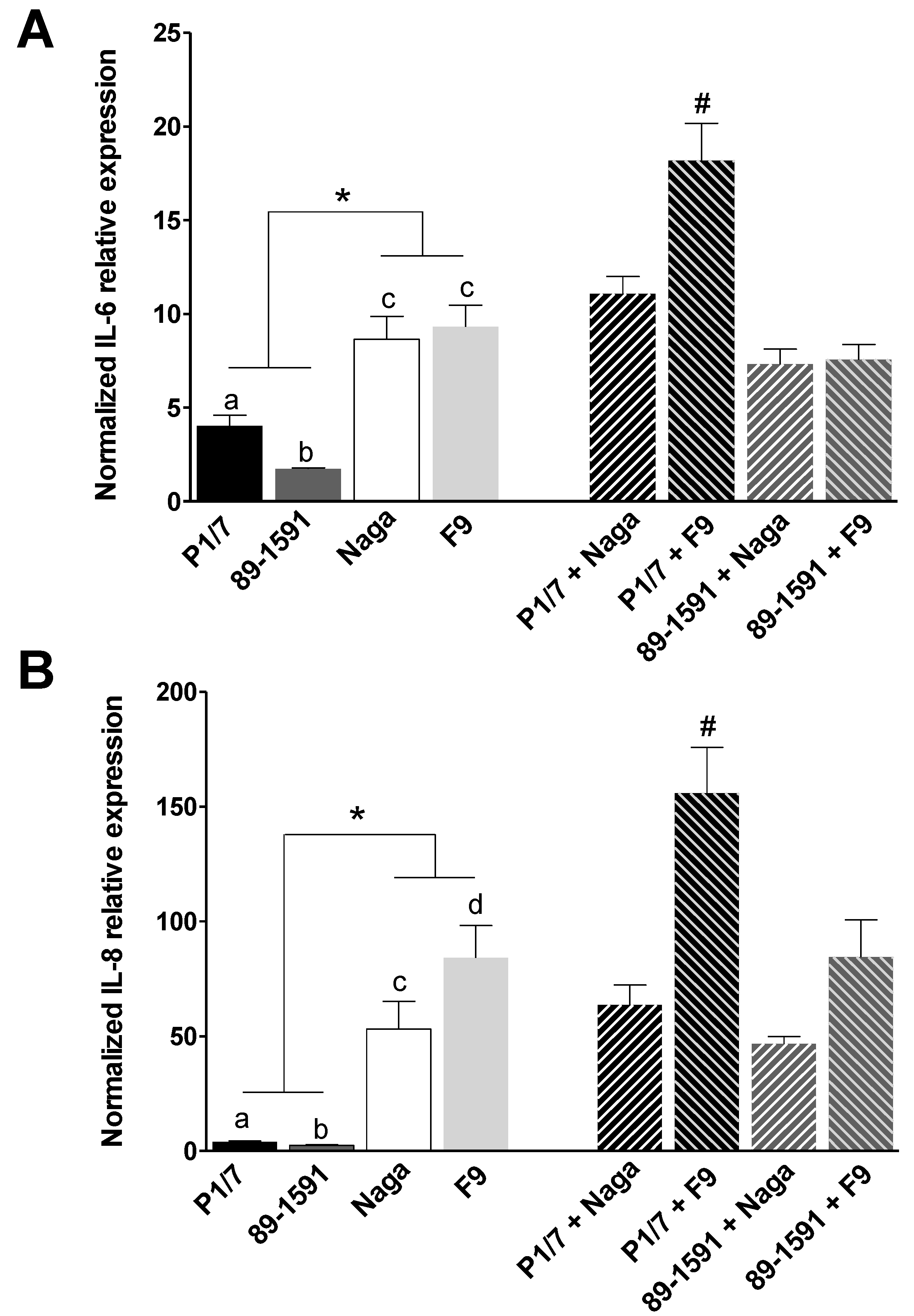
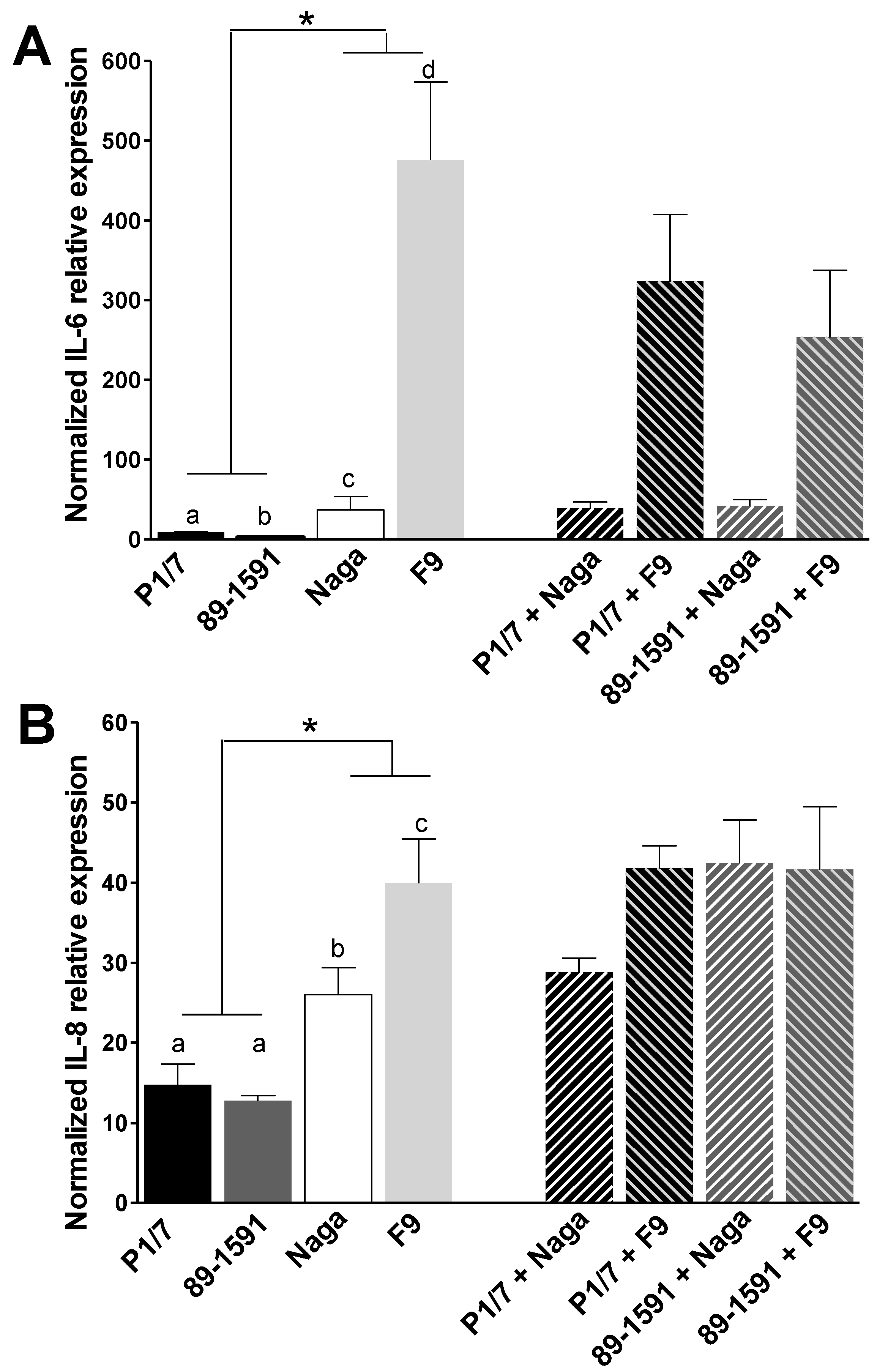
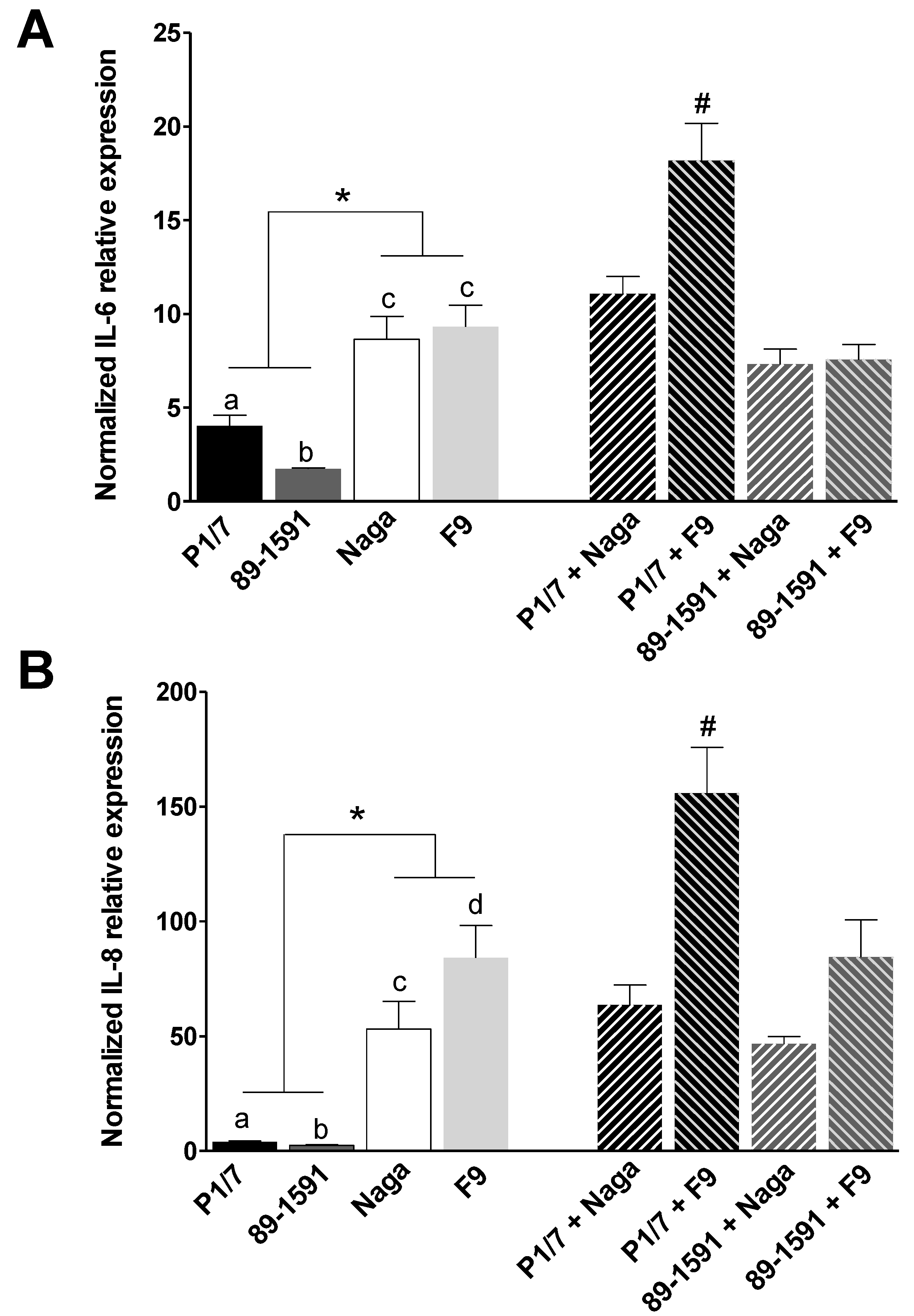
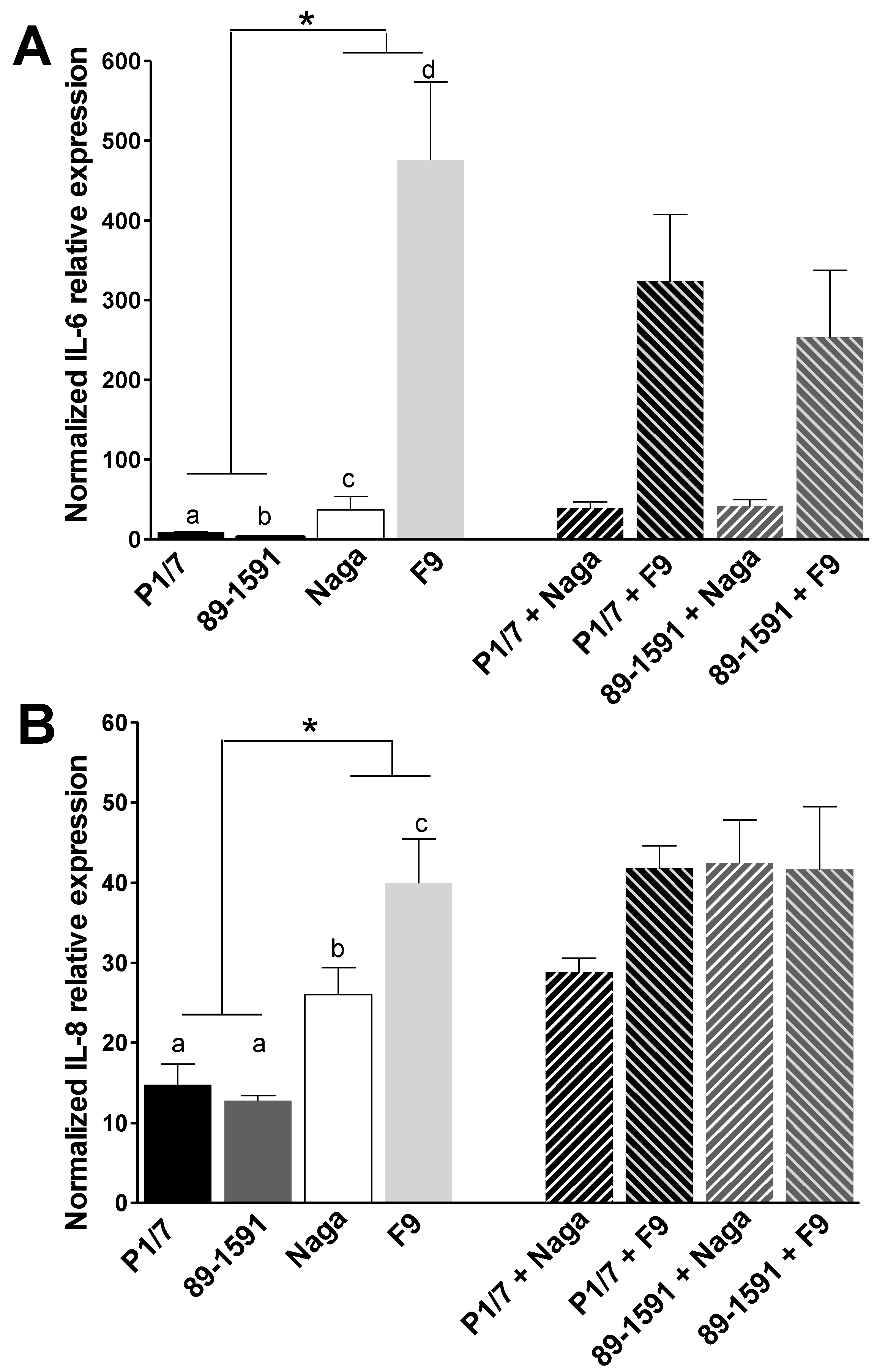
2.2. H. parasuis Induces a Higher Expression of Pro-Inflammatory Cytokines by Tracheal Porcine Epithelial Cells and Primary Alveolar Macrophages than S. suis and Co-Infections Only Partially Modulate This Expression
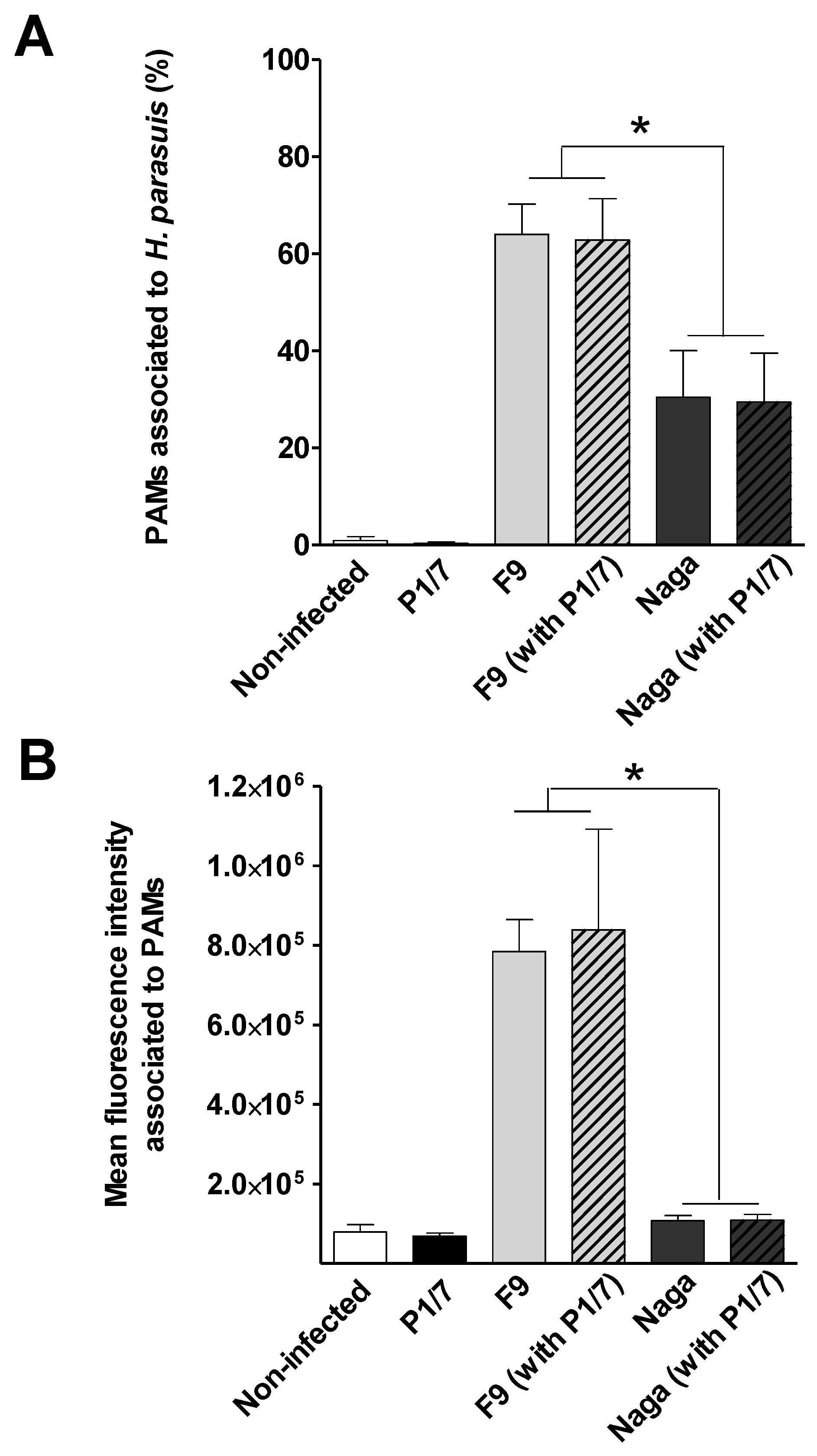
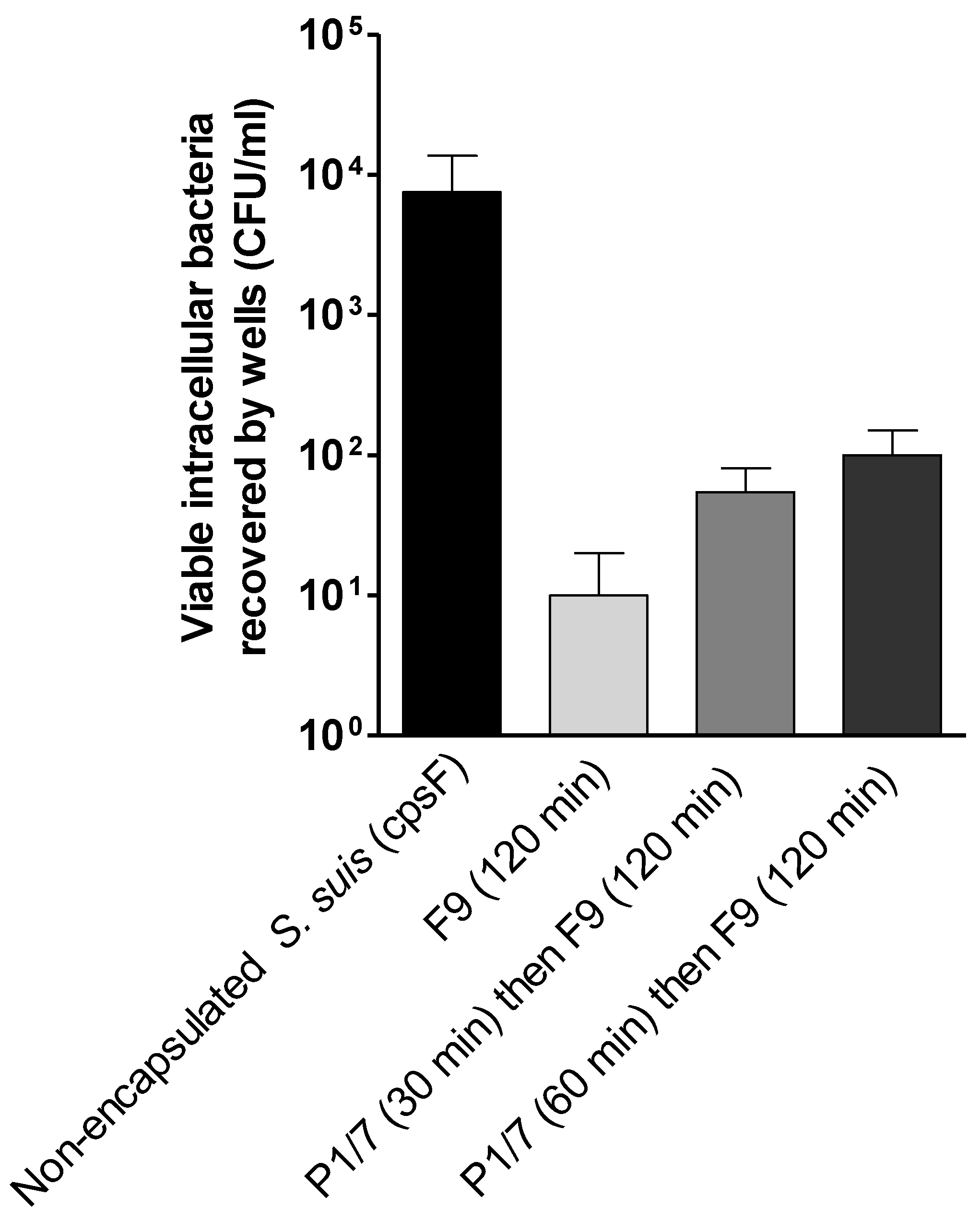
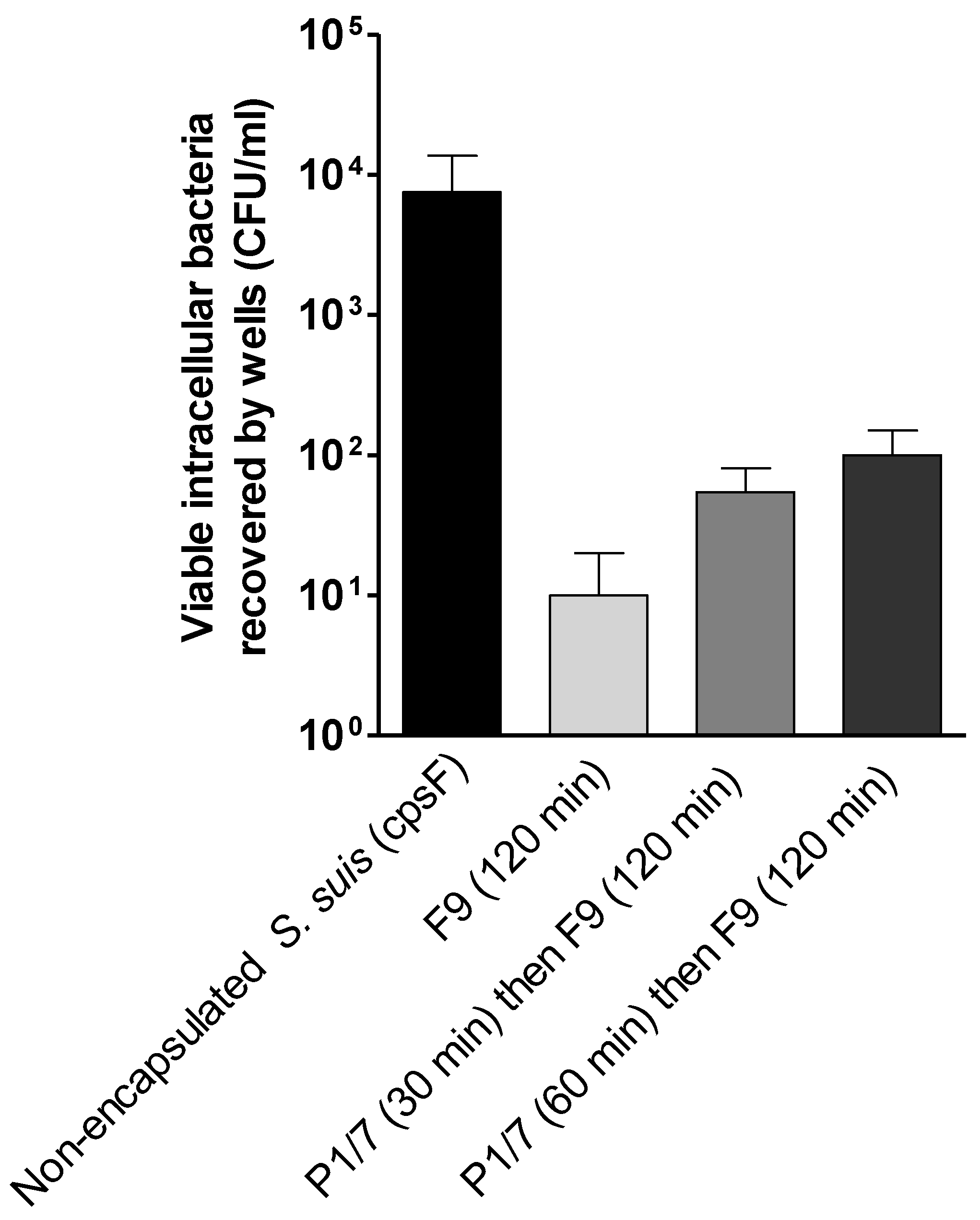
2.3. S. suis Has No Effect on H. parasuis Association to or Phagocytosis by PAMs
3. Materials and Methods
3.1. Bacterial Strains
3.2. Cell Culture
3.3. Adhesion and Invasion of NPTr by S. suis and H. parasuis
3.4. Induction of Pro-Inflammatory Cytokine Expression
3.5. RNA Extraction, cDNA Construct and RT-qPCR
3.6. Cytotoxicity
3.7. Effect of S. suis on H. parasuis Association to and Phagocytosis by PAMs
3.7.1. Association of H. parasuis to PAMs by FACS
3.7.2. Phagocytosis of H. parasuis by PAMs by the Antibiotic-Protection Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| cDNA | Complementary DNA |
| CFU | Colony forming unit |
| CPS | Capsular polysaccharide |
| DMEM | Dulbecco’s modified Eagle’s Medium |
| EDTA | Ethylenediaminetetraacetic acid |
| FACS | Fluorescence associated cell sorting |
| FBS | Fetal bovine serum |
| FITC | Fluoroscein isothiocyanate |
| GbO3 | globotriaosylceramide |
| Hypox | hypoxanthine phosphoribosyl transferase 1 |
| IL | Interleukin |
| LDH | lactate dehydrogenase |
| LPS | Lipopolysaccharide |
| MFI | Mean fluorescence intensities |
| MOI | Multiplicity of infection |
| NPTr | Newborn Pig Tracheal cells |
| PAMs | Primary alveolar macrophages |
| PBS | Phosphate-buffered saline |
| PCR | Polymerization chain reaction |
| PPIA | Peptidylprolyl isomerase A |
| RT-PCR | Reverse transcription polymerization chain reaction |
| ST | Sequence type |
| THA | Todd-Hewitt broth agar |
| THB | Todd-Hewitt broth |
| VtaA | Trimeric autotransporter |
References
- Gottschalk, M.; Segura, M.; Xu, J. Streptococcus suis infections in humans: The chinese experience and the situation in North America. Anim. Health Res. Rev. 2007, 8, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Costa-Hurtado, M.; Aragon, V. Advances in the quest for virulence factors of Haemophilus parasuis. Vet. J. 2013, 198, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Okura, M.; Osaki, M.; Nomoto, R.; Arai, S.; Osawa, R.; Sekizaki, T.; Takamatsu, D. Current taxonomical situation of Streptococcus suis. Pathogens 2016, 5, 45. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Qiu, X.; Roy, D.; Segura, M.; Du, P.; Xu, J.; Gottschalk, M. Genotyping and investigating capsular polysaccharide synthesis gene loci of non-serotypeable Streptococcus suis isolated from diseased pigs in Canada. Vet. Res. 2017, 48, 10. [Google Scholar] [CrossRef] [PubMed]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes Infect. 2014, 3, e45. [Google Scholar] [CrossRef] [PubMed]
- Fittipaldi, N.; Xu, J.; Lacouture, S.; Tharavichitkul, P.; Osaki, M.; Sekizaki, T.; Takamatsu, D.; Gottschalk, M. Lineage and virulence of Streptococcus suis serotype 2 isolates from North America. Emerg. Infect. Dis. 2011, 17, 2239–2244. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Calzas, C.; Grenier, D.; Gottschalk, M. Initial steps of the pathogenesis of the infection caused by Streptococcus suis: Fighting against nonspecific defenses. FEBS Lett. 2016, 590, 3772–3799. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical streptococcus suis virulence factors: Are they all really critical? Trends Microbiol. 2017, 25, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Houde, M.; Gottschalk, M.; Gagnon, F.; Van Calsteren, M.R.; Segura, M. Streptococcus suis capsular polysaccharide inhibits phagocytosis through destabilization of lipid microdomains and prevents lactosylceramide-dependent recognition. Infect. Immun. 2012, 80, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Lachance, C.; Wang, Y.; Gagnon, C.A.; Savard, C.; Segura, M.; Grenier, D.; Gottschalk, M. Transcriptional approach to study porcine tracheal epithelial cells individually or dually infected with Swine Influenza virus and Streptococcus suis. BMC Vet. Res. 2014, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.; Pijoan, C. Haemophilus parasuis: New trends on diagnosis, epidemiology and control. Vet. Microbiol. 2004, 99, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wang, L.; Chu, Y.; Li, X.; Cui, Y.; Chen, S.; Zhou, J.; Li, C.; Lu, Z.; Liu, J.; et al. Characterization of chinese Haemophilus parasuis isolates by traditional serotyping and molecular serotyping methods. PLoS ONE 2016, 11, e0168903. [Google Scholar] [CrossRef] [PubMed]
- Lacouture, S.; Rodriguez, E.; Strutzberg-Minder, E.; Gottschalk, M. Serotyping of Haemophilus parasuis field isolates from diseased pigs in quebec by indirect hemagglutination assay and multiplex polymerase chain reaction (pcr). Can. Vet. J. 2017, 58, 802–804. [Google Scholar] [PubMed]
- Frandoloso, R.; Martinez-Martinez, S.; Gutierrez-Martin, C.B.; Rodriguez-Ferri, E.F. Haemophilus parasuis serovar 5 nagasaki strain adheres and invades pk-15 cells. Vet. Microbiol. 2012, 154, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Olvera, A.; Ballester, M.; Nofrarias, M.; Sibila, M.; Aragon, V. Differences in phagocytosis susceptibility in Haemophilus parasuis strains. Vet. Res. 2009, 40, 24. [Google Scholar] [CrossRef] [PubMed]
- Boerlin, P.; Poljak, Z.; Gallant, J.; Chalmers, G.; Nicholson, V.; Soltes, G.A.; MacInnes, J.I. Genetic diversity of Haemophilus parasuis from sick and healthy pigs. Vet. Microbiol. 2013, 167, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Galofre-Mila, N.; Correa-Fiz, F.; Lacouture, S.; Gottschalk, M.; Strutzberg-Minder, K.; Bensaid, A.; Pina-Pedrero, S.; Aragon, V. A robust pcr for the differentiation of potential virulent strains of Haemophilus parasuis. BMC Vet. Res. 2017, 13, 124. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gagnon, C.A.; Savard, C.; Music, N.; Srednik, M.; Segura, M.; Lachance, C.; Bellehumeur, C.; Gottschalk, M. Capsular sialic acid of Streptococcus suis serotype 2 binds to swine influenza virus and enhances bacterial interactions with virus-infected tracheal epithelial cells. Infect. Immun. 2013, 81, 4498–4508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrando, M.L.; Fuentes, S.; de Greeff, A.; Smith, H.; Wells, J.M. Apua, a multifunctional alpha-glucan-degrading enzyme of Streptococcus suis, mediates adhesion to porcine epithelium and mucus. Microbiology 2010, 156, 2818–2828. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.H.; Meng, F.; Seitz, M.; Valentin-Weigand, P.; Herrler, G. Sialic acid-dependent interactions between influenza viruses and Streptococcus suis affect the infection of porcine tracheal cells. J. Gen. Virol. 2015, 96, 2557–2568. [Google Scholar] [CrossRef] [PubMed]
- Tenenbaum, T.; Asmat, T.; Seitz, M.; Schroten, H.; Schwerk, C. Biological activities of suilysin: Role in Streptococcus suis pathogenesis. Future Microbiol. 2016, 11, 941–954. [Google Scholar] [CrossRef] [PubMed]
- Seitz, M.; Baums, C.G.; Neis, C.; Benga, L.; Fulde, M.; Rohde, M.; Goethe, R.; Valentin-Weigand, P. Subcytolytic effects of suilysin on interaction of Streptococcus suis with epithelial cells. Vet. Microbiol. 2013, 167, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, B.; Vanier, G.; Jacques, M.; Auger, E.; Gottschalk, M. Studies on the interactions of Haemophilus parasuis with porcine epithelial tracheal cells: Limited role of los in apoptosis and pro-inflammatory cytokine release. Microb. Pathog. 2009, 46, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Auger, E.; Deslandes, V.; Ramjeet, M.; Contreras, I.; Nash, J.H.; Harel, J.; Gottschalk, M.; Olivier, M.; Jacques, M. Host-pathogen interactions of Actinobacillus pleuropneumoniae with porcine lung and tracheal epithelial cells. Infect. Immun. 2009, 77, 1426–1441. [Google Scholar] [CrossRef] [PubMed]
- De Greeff, A.; Benga, L.; Wichgers Schreur, P.J.; Valentin-Weigand, P.; Rebel, J.M.; Smith, H.E. Involvement of nf-kappab and map-kinases in the transcriptional response of alveolar macrophages to Streptococcus suis. Vet. Microbiol. 2010, 141, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Costa-Hurtado, M.; Olvera, A.; Martinez-Moliner, V.; Galofre-Mila, N.; Martinez, P.; Dominguez, J.; Aragon, V. Changes in macrophage phenotype after infection of pigs with Haemophilus parasuis strains with different levels of virulence. Infect. Immun. 2013, 81, 2327–2333. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Yuan, F.; Zhang, M.; Tan, C.; Chen, H.; Bei, W. Cloning, expression and characterization of a cell wall surface protein, 6-phosphogluconate dehydrogenase, of Haemophilus parasuis. Res. Vet. Sci. 2012, 93, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, T.; Langford, P.; Hua, K.; Zhou, S.; Zhai, Y.; Xiao, H.; Luo, R.; Bi, D.; Jin, H.; et al. Haemophilus parasuis induces activation of nf-kappab and map kinase signaling pathways mediated by toll-like receptors. Mol. Immunol. 2015, 65, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Lecours, M.P.; Segura, M.; Fittipaldi, N.; Rivest, S.; Gottschalk, M. Immune receptors involved in Streptococcus suis recognition by dendritic cells. PLoS ONE 2012, 7, e44746. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, S.; Hua, K.; Xiao, H.; Li, Z.; Liu, M.; Luo, R.; Bi, D.; Zhou, R.; Jin, H. Haemophilus parasuis infection activates chemokine rantes in pk-15 cells. Mol. Immunol. 2015, 67, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Van Calsteren, M.R.; Gagnon, F.; Lacouture, S.; Fittipaldi, N.; Gottschalk, M. Structure determination of Streptococcus suis serotype 2 capsular polysaccharide. Biochem. Cell Biol. 2010, 88, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.P.; Fittipaldi, N.; Benoit-Biancamano, M.O.; Segura, M.; Gottschalk, M. Virulence studies of different sequence types and geographical origins of Streptococcus suis serotype 2 in a mouse model of infection. Pathogens 2016, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Vanier, G.; Segura, M.; Friedl, P.; Lacouture, S.; Gottschalk, M. Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infect. Immun. 2004, 72, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.P.; Christodoulides, M.; Segura, M.; Xu, J.; Gottschalk, M. Interactions of Streptococcus suis serotype 2 with human meningeal cells and astrocytes. BMC Res. Notes 2015, 8, 607. [Google Scholar] [CrossRef] [PubMed]
- Athey, T.B.; Teatero, S.; Takamatsu, D.; Wasserscheid, J.; Dewar, K.; Gottschalk, M.; Fittipaldi, N. Population structure and antimicrobial resistance profiles of Streptococcus suis serotype 2 sequence type 25 strains. PLoS ONE 2016, 11, e0150908. [Google Scholar] [CrossRef] [PubMed]
- Holden, M.T.; Hauser, H.; Sanders, M.; Ngo, T.H.; Cherevach, I.; Cronin, A.; Goodhead, I.; Mungall, K.; Quail, M.A.; Price, C.; et al. Rapid evolution of virulence and drug resistance in the emerging zoonotic pathogen Streptococcus suis. PLoS ONE 2009, 4, e6072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morozumi, T.; Nicolet, J. Some antigenic properties of Haemophilus parasuis and a proposal for serological classification. J. Clin. Microbiol. 1986, 23, 1022–1025. [Google Scholar] [PubMed]
- Ferrari, M.; Scalvini, A.; Losio, M.N.; Corradi, A.; Soncini, M.; Bignotti, E.; Milanesi, E.; Ajmone-Marsan, P.; Barlati, S.; Bellotti, D.; et al. Establishment and characterization of two new pig cell lines for use in virological diagnostic laboratories. J. Virol. Methods 2003, 107, 205–212. [Google Scholar] [CrossRef]
- McCaig, W.D.; Loving, C.L.; Hughes, H.R.; Brockmeier, S.L. Characterization and vaccine potential of outer membrane vesicles produced by Haemophilus parasuis. PLoS ONE 2016, 11, e0149132. [Google Scholar] [CrossRef] [PubMed]
- Lecours, M.P.; Segura, M.; Lachance, C.; Mussa, T.; Surprenant, C.; Montoya, M.; Gottschalk, M. Characterization of porcine dendritic cell response to Streptococcus suis. Vet. Res. 2011, 42, 72. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | ST or Serovar | Country | Host | Virulence/Sly | Reference |
|---|---|---|---|---|---|
| S. suis serotype 2 | |||||
| P1/7 | 1 | UK | Pig | High virulent/Yes | [35] |
| 89-1591 | 25 | Canada | Pig | Intermediate virulent/No | [36,37] |
| H. parasuis | |||||
| F9 | 6 | Spain | Pig | Low-virulent | [27] |
| Nagasaki | 5 | Japan | Pig | Virulent | [38] |
| Gene Name | Forward | Reverse |
|---|---|---|
| IL-6 | ACTCCCTCTCCACAAGCGCCTT | TGGCATCTTCTTCCAGGCGTCCC |
| IL-8 | TGTGAGGCTGCAGTTCTGGCAAG | GGGTGGAAAGGTGTGGAATGCGT |
| Hypox | GCAGCCCCAGCGTCGTGATT | CGAGCAAGCCGTTCAGTCCTGT |
| PPIA | TGCAGACAAAGTTCCAAAGACAG | GCCACCAGTGCCATTATGG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathieu-Denoncourt, A.; Letendre, C.; Auger, J.-P.; Segura, M.; Aragon, V.; Lacouture, S.; Gottschalk, M. Limited Interactions between Streptococcus Suis and Haemophilus Parasuis in In Vitro Co-Infection Studies. Pathogens 2018, 7, 7. https://doi.org/10.3390/pathogens7010007
Mathieu-Denoncourt A, Letendre C, Auger J-P, Segura M, Aragon V, Lacouture S, Gottschalk M. Limited Interactions between Streptococcus Suis and Haemophilus Parasuis in In Vitro Co-Infection Studies. Pathogens. 2018; 7(1):7. https://doi.org/10.3390/pathogens7010007
Chicago/Turabian StyleMathieu-Denoncourt, Annabelle, Corinne Letendre, Jean-Philippe Auger, Mariela Segura, Virginia Aragon, Sonia Lacouture, and Marcelo Gottschalk. 2018. "Limited Interactions between Streptococcus Suis and Haemophilus Parasuis in In Vitro Co-Infection Studies" Pathogens 7, no. 1: 7. https://doi.org/10.3390/pathogens7010007
APA StyleMathieu-Denoncourt, A., Letendre, C., Auger, J.-P., Segura, M., Aragon, V., Lacouture, S., & Gottschalk, M. (2018). Limited Interactions between Streptococcus Suis and Haemophilus Parasuis in In Vitro Co-Infection Studies. Pathogens, 7(1), 7. https://doi.org/10.3390/pathogens7010007