
A Comparative Analysis of the Mechanism of Toll-Like Receptor-Disruption by TIR-Containing Protein C from Uropathogenic Escherichia coli


Abstract
:1. Introduction
2. Results and Discussion
2.1. The TIR-Containing Protein C and Its Relatives
2.2. Function of TcpC and Other Bacterial Tcps
2.3. Interaction of TcpC and Other Bacterial Tcps with TLR-Components
2.4. Structure of Bacterial Tcps
2.5. TIR-Domains from Bacterial Tcps, TLRs and TLR-Adaptors Differ from Each Other
3. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Foxman, B. Epidemiology of urinary tract infections: Incidence, morbidity, and economic costs. Am. J. Med. 2002, 113, 5S–13S. [Google Scholar] [CrossRef]
- Ironmonger, D.; Edeghere, O.; Bains, A.; Loy, R.; Woodford, N.; Hawkey, P.M. Surveillance of antibiotic susceptibility of urinary tract pathogens for a population of 5.6 million over 4 years. J. Antimicrob. Chemother. 2015, 70, 1744–1750. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Basu, S.; Mukherjee, S.K.; Majumder, M. Multidrug-Resistance and Extended Spectrum Beta-Lactamase Production in Uropathogenic E. Coli which were Isolated from Hospitalized Patients in Kolkata, India. J. Clin. Diagn. Res. 2013, 7, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Broz, P.; Monack, D.M. Newly described pattern recognition receptors team up against intracellular pathogens. Nat. Rev. Immunol. 2013, 13, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Low, L.Y.; Mukasa, T.; Reed, J.C.; Pascual, J. Characterization of a TIR-like protein from Paracoccus denitrificans. Biochem. Biophys. Res. Commun. 2007, 356, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Newman, R.M.; Salunkhe, P.; Godzik, A.; Reed, J.C. Identification and characterization of a novel bacterial virulence factor that shares homology with mammalian Toll/interleukin-1 receptor family proteins. Infect. Immunity 2006, 74, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Cirl, C.; Wieser, A.; Yadav, M.; Duerr, S.; Schubert, S.; Fischer, H.; Stappert, D.; Wantia, N.; Rodriguez, N.; Wagner, H.; et al. Subversion of Toll-like receptor signaling by a unique family of bacterial Toll/interleukin-1 receptor domain-containing proteins. Nat. Med. 2008, 14, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, S.P.; Marchesini, M.I.; Lelouard, H.; Fugier, E.; Jolly, G.; Balor, S.; Muller, A.; Lapaque, N.; Demaria, O.; Alexopoulou, L.; et al. Brucella Control of Dendritic Cell Maturation Is Dependent on the TIR-Containing Protein Btp1. PLoS Pathog. 2008, 4, e21. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Baghdayan, A.S.; Payne, S.J.; Shankar, N. A TIR domain protein from E. faecalis attenuates MyD88-mediated signaling and NF-kappaB activation. PLoS ONE 2014, 9, e112010. [Google Scholar] [CrossRef] [PubMed]
- Rana, R.R.; Simpson, P.; Zhang, M.; Jennions, M.; Ukegbu, C.; Spear, A.M.; Alguel, Y.; Matthews, S.J.; Atkins, H.S.; Byrne, B. Yersinia pestis TIR-domain protein forms dimers that interact with the human adaptor protein MyD88. Microb. Pathog. 2011, 51, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Norenberg, D.; Wieser, A.; Magistro, G.; Hoffmann, C.; Meyer, C.; Messerer, M.; Schubert, S. Molecular analysis of a novel Toll/interleukin-1 receptor (TIR)-domain containing virulence protein of Y. pseudotuberculosis among Far East scarlet-like fever serotype I strains. Int. J. Med. Microbiol. 2013, 303, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Cirl, C.; Miethke, T. Microbial Toll/interleukin 1 receptor proteins: A new class of virulence factors. Int. J. Med. Microbiol. 2010, 300, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Askarian, F.; van Sorge, N.M.; Sangvik, M.; Beasley, F.C.; Henriksen, J.R.; Sollid, J.U.; van Strijp, J.A.; Nizet, V.; Johannessen, M. A Staphylococcus aureus TIR domain protein virulence factor blocks TLR2-mediated NF-kappaB signaling. J. Innate Immunity 2014, 6, 485–498. [Google Scholar]
- Schubert, S.; Norenberg, D.; Clermont, O.; Magistro, G.; Wieser, A.; Romann, E.; Hoffmann, C.; Weinert, K.; Denamur, E. Prevalence and phylogenetic history of the TcpC virulence determinant in Escherichia coli. Int. J. Med. Microbiol. 2010, 300, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, G.K.; Yu, Q.; Harms, J.S.; Splitter, G.A. Brucella TIR Domain-containing Protein Mimics Properties of the Toll-like Receptor Adaptor Protein TIRAP. J. Biol. Chem. 2009, 284, 9892–9898. [Google Scholar] [CrossRef] [PubMed]
- Alaidarous, M.; Ve, T.; Casey, L.W.; Valkov, E.; Ericsson, D.J.; Ullah, M.O.; Schembri, M.A.; Mansell, A.; Sweet, M.J.; Kobe, B. Mechanism of bacterial interference with TLR4 signaling by Brucella Toll/interleukin-1 receptor domain-containing protein TcpB. J. Biol. Chem. 2014, 289, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Spear, A.M.; Loman, N.J.; Atkins, H.S.; Pallen, M.J. Microbial TIR domains: not necessarily agents of subversion? Trends Microbiol. 2009, 17, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.; Zhang, J.; Fischer, H.; Huang, W.; Lutay, N.; Cirl, C.; Lum, J.; Miethke, T.; Svanborg, C. Inhibition of TIR domain signaling by TcpC: MyD88-dependent and independent effects on Escherichia coli virulence. PLoS Pathog. 2010, 6, e1001120. [Google Scholar] [CrossRef] [PubMed]
- Snyder, G.A.; Cirl, C.; Jiang, J.; Chen, K.; Waldhuber, A.; Smith, P.; Römmler, F.; Snyder, N.; Fresquez, T.; Durr, S.; et al. Molecular mechanisms for the subversion of MyD88 signaling by TcpC from virulent uropathogenic Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 6985–6990. [Google Scholar] [CrossRef] [PubMed]
- Kaplan-Turkoz, B.; Koelblen, T.; Felix, C.; Candusso, M.P.; O’Callaghan, D.; Vergunst, A.C.; Terradot, L. Structure of the Toll/interleukin 1 receptor (TIR) domain of the immunosuppressive Brucella effector BtpA/Btp1/TcpB. FEBS Lett. 2013, 587, 3412–3416. [Google Scholar] [CrossRef] [PubMed]
- Snyder, G.A.; Deredge, D.; Waldhuber, A.; Fresquez, T.; Wilkins, D.Z.; Smith, P.T.; Durr, S.; Cirl, C.; Jiang, J.; Jennings, W.; et al. Crystal structures of the Toll/Interleukin-1 receptor (TIR) domains from the Brucella protein TcpB and host adaptor TIRAP reveal mechanisms of molecular mimicry. J. Biol. Chem. 2014, 289, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, S.P.; Marchesini, M.I.; Degos, C.; Terwagne, M.; von Bargen, K.; Lepidi, H.; Herrmann, C.K.; Santos Lacerda, T.L.; Imbert, P.R.; Pierre, P.; et al. BtpB, a novel Brucella TIR-containing effector protein with immune modulatory functions. Front. Cell Infect. Microbiol. 2013, 3, 28. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, T.D.; Haro, O.D.Q.; Domann, E.; Chakraborty, T.; Tchatalbachev, S. The TIR Domain Containing Locus of Enterococcus faecalis Is Predominant among Urinary Tract Infection Isolates and Downregulates Host Inflammatory Response. Int. J. Microbiol. 2014, 2014, 918143. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.L.; Low, L.Y.; Hsu, S.; Li, S.; Liu, T.; Santelli, E.; le Negrate, G.; Reed, J.C.; Woods, V.L., Jr.; Pascual, J. Molecular mimicry in innate immunity: Crystal structure of a bacterial TIR domain. J. Biol. Chem. 2009, 284, 21386–21392. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.; Koblansky, A.; Gaines, J.; Brown, T.; West, A.P.; Zhang, D.; Nishikawa, T.; Park, S.G.; Roop, R.M., II; et al. Subversion of innate immune responses by Brucella through the targeted degradation of the TLR signaling adapter, MAL. J. Immunol. 2010, 184, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Yamamoto, M.; Akira, S.; Beutler, B.; Svanborg, C. Mechanism of pathogen-specific TLR4 activation in the mucosa: Fimbriae, recognition receptors and adaptor protein selection. Eur. J. Immunol. 2006, 36, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Tao, X.; Shen, B.; Horng, T.; Medzhitov, R.; Manley, J.L.; Tong, L. Structural basis for signal transduction by the Toll/interleukin-1 receptor domains. Nature 2000, 408, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Ve, T.; Williams, S.J.; Kobe, B. Structure and function of Toll/interleukin-1 receptor/resistance protein (TIR) domains. Apoptosis 2015, 20, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Rana, R.R.; Zhang, M.; Spear, A.M.; Atkins, H.S.; Byrne, B. Bacterial TIR-containing proteins and host innate immune system evasion. Med. Microbiol. Immunol. 2013, 202, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nyman, T.; Stenmark, P.; Flodin, S.; Johansson, I.; Hammarstrom, M.; Nordlund, P. The crystal structure of the human toll-like receptor 10 cytoplasmic domain reveals a putative signaling dimer. J. Biol. Chem. 2008, 283, 11861–11865. [Google Scholar] [CrossRef] [PubMed]
- Valkov, E.; Stamp, A.; Dimaio, F.; Baker, D.; Verstak, B.; Roversi, P.; Kellie, S.; Sweet, M.J.; Mansell, A.; Gay, N.J.; et al. Crystal structure of Toll-like receptor adaptor MAL/TIRAP reveals the molecular basis for signal transduction and disease protection. Proc. Natl. Acad. Sci. USA 2011, 108, 14879–14884. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Lu, J.; Zhou, W.; Shen, Y. Structural insights into TIR domain specificity of the bridging adaptor Mal in TLR4 signaling. PLoS One 2012, 7, e34202. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.R.; Kim, S.; Shoelson, S.E.; Park, S. X-ray Crystallographic Structure of TIR-Domain from the Human TIR-Domain Containing Adaptor Protein/MyD88 Adaptor-Like Protein (TIRAP/MAL). Bull. Korean Chem. Soc. 2013, 33, 3091–3094. [Google Scholar] [CrossRef]
- Snyder, G.A.; Sundberg, E.J. Molecular interactions in interleukin and toll-like receptor signaling pathways. Curr. Pharm. Des. 2014, 20, 1244–1258. [Google Scholar] [CrossRef] [PubMed]
- Mills, K.H. TLR-dependent T cell activation in autoimmunity. Nat. Rev. Immunol. 2011, 11, 807–822. [Google Scholar] [CrossRef] [PubMed]
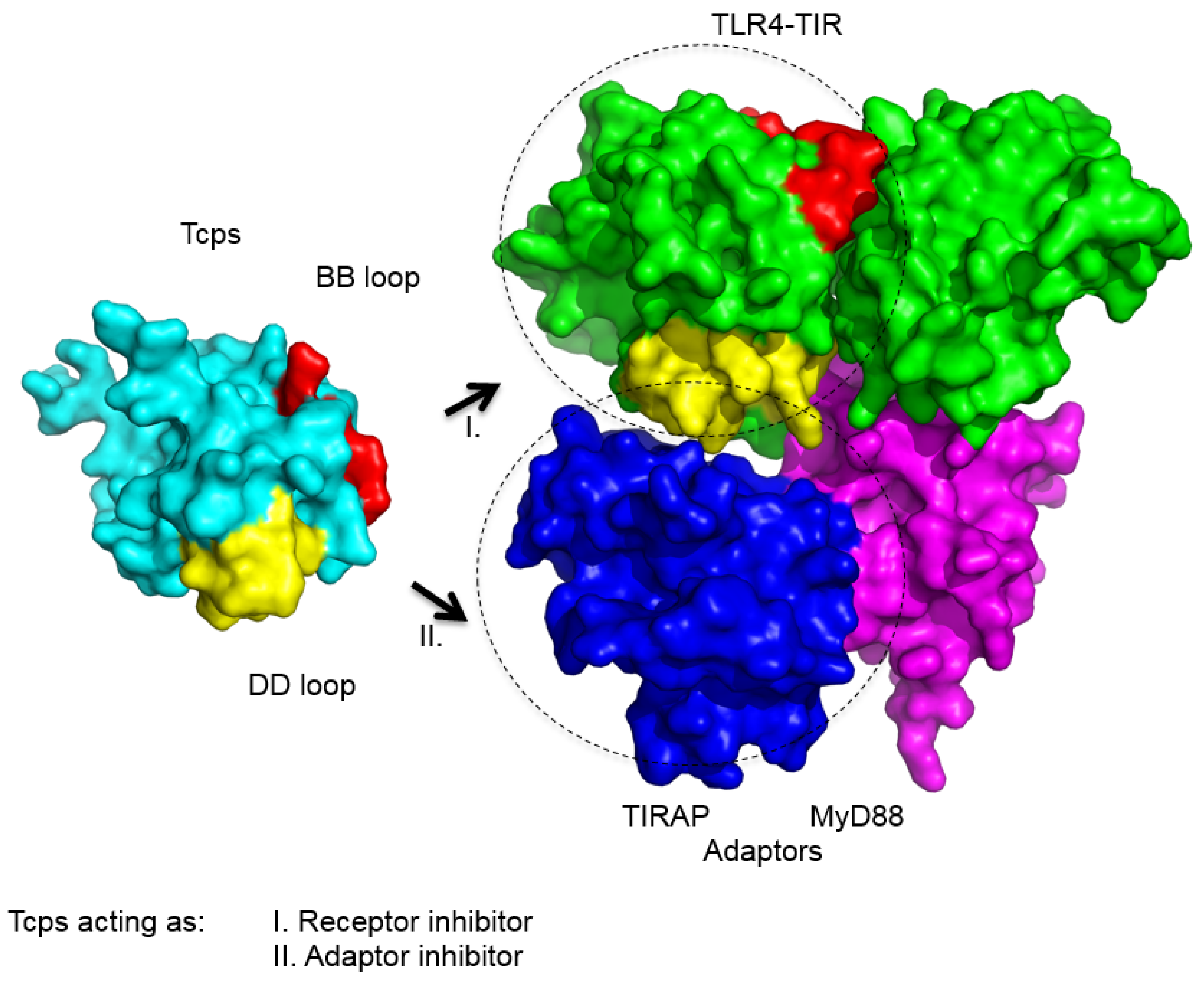

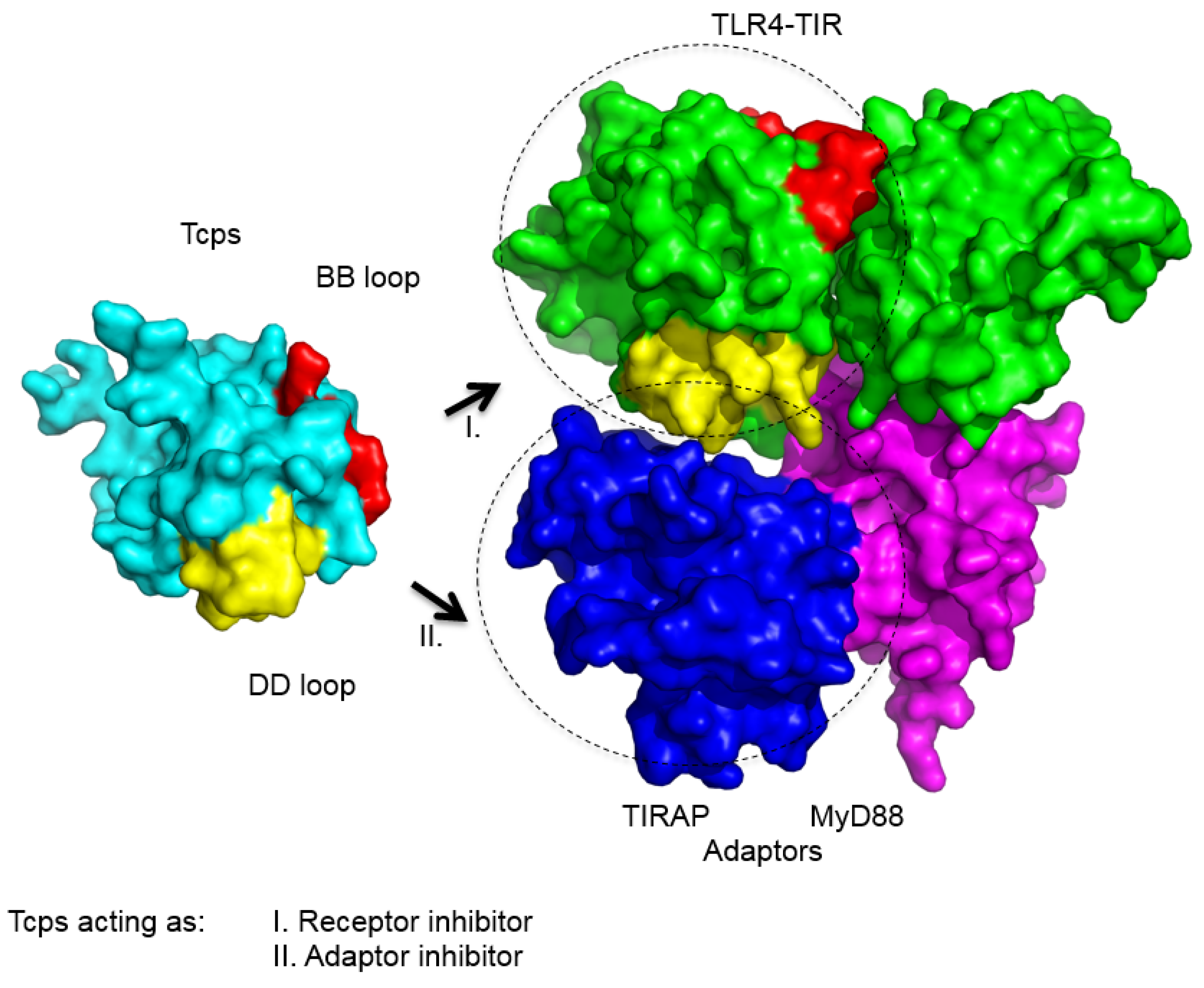{kind=link}
| Bacterial TIR Protein | Organism | TIR Protein Function in vitro | TIR Protein Function in vivo | Protein Interaction with | Protein Structure | References |
|---|---|---|---|---|---|---|
| TcpC | Escherichia coli CFT073 | suppresses NF-κB activation, promotes bacterial survival in RAW264.7 macrophages | promotes virulence in UTI mouse model (bacterial burden, kidney disease) | MyD88, TLR4 | not known | Cirl et al., 2008 [7]; Yadav et al., 2010 [18]; Snyder et al., 2013 [19] |
| TcpB (Btp1/BtpA) | Brucella sp. | suppresses NF-κB activation, inhibition of dendritic cell maturation | promotes virulence during early stages of infection in mice (systemic spread) | MyD88, TIRAP, TLR4, Microtubule | PDB IDs: 4C7M, 4LZP, 4LQC | Cirl et al., 2008 [7]; Salcedo et al., 2008 [8]; Radhakrishnan et al., 2009 [15]; Kaplan-Türköz et al., 2013 [20]; Snyder et al., 2014 [21]; Alaidarous et al., 2014 [16]; |
| BtpB | Brucella sp. | suppresses TLR2, TLR4, TLR5, TLR9 mediated activation of NF-κB | promotes virulence in mice (survival) | not known | not known | Salcedo et al., 2013 [22]; |
| TlpA | Salmonella enterica serovar Enteriditis | suppresses NF-κB activation, promotes bacterial survival in human THP1 macrophages | promotes virulence in mice (bacterial survival, burden and lethality) | not known | not known | Newman et al., 2006 [6]; |
| TcpYI | Yersinia pseudotuberculosis | promotes bacterial survival in murine macrophages | promotes virulence in mouse model of peritonitis (survival inside the spleen) | not known | not known | Nörenberg et al., 2013 [11]; |
| YpTIR | Yersinia pestis | suppresses NF-κB activation | no role in virulence | MyD88 | not known | Rana et al., 2011 [10]; |
| TcpF | Enteroccocus faecalis | suppresses NF-κB activation | promotes virulence in mice (bacterial burden) | MyD88 | not known | Kraemer et al., 2014 [23]; Zou et al., 2014 [9]; |
| TirS | Staphylococcus aureus MSSA476 | suppresses NF-κB and MAP-kinase activation | Promotes virulence in mice (organ co-lonization) | not known | not known | Askarian et al., 2014 [13]; |
| PdTlp | Paracoccus denitrificans | not known | not known | MyD88, TLR4 | PDB ID: 3H16 | Low et al., 2007 [5]; Chan et al., 2009 [24]; |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waldhuber, A.; Snyder, G.A.; Römmler, F.; Cirl, C.; Müller, T.; Xiao, T.S.; Svanborg, C.; Miethke, T. A Comparative Analysis of the Mechanism of Toll-Like Receptor-Disruption by TIR-Containing Protein C from Uropathogenic Escherichia coli. Pathogens 2016, 5, 25. https://doi.org/10.3390/pathogens5010025
Waldhuber A, Snyder GA, Römmler F, Cirl C, Müller T, Xiao TS, Svanborg C, Miethke T. A Comparative Analysis of the Mechanism of Toll-Like Receptor-Disruption by TIR-Containing Protein C from Uropathogenic Escherichia coli. Pathogens. 2016; 5(1):25. https://doi.org/10.3390/pathogens5010025
Chicago/Turabian StyleWaldhuber, Anna, Greg A. Snyder, Franziska Römmler, Christine Cirl, Tina Müller, Tsan Sam Xiao, Catharina Svanborg, and Thomas Miethke. 2016. "A Comparative Analysis of the Mechanism of Toll-Like Receptor-Disruption by TIR-Containing Protein C from Uropathogenic Escherichia coli" Pathogens 5, no. 1: 25. https://doi.org/10.3390/pathogens5010025
APA StyleWaldhuber, A., Snyder, G. A., Römmler, F., Cirl, C., Müller, T., Xiao, T. S., Svanborg, C., & Miethke, T. (2016). A Comparative Analysis of the Mechanism of Toll-Like Receptor-Disruption by TIR-Containing Protein C from Uropathogenic Escherichia coli. Pathogens, 5(1), 25. https://doi.org/10.3390/pathogens5010025