
Neutrophil Migration into the Infected Uroepithelium Is Regulated by the Crosstalk between Resident and Helper Macrophages


{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
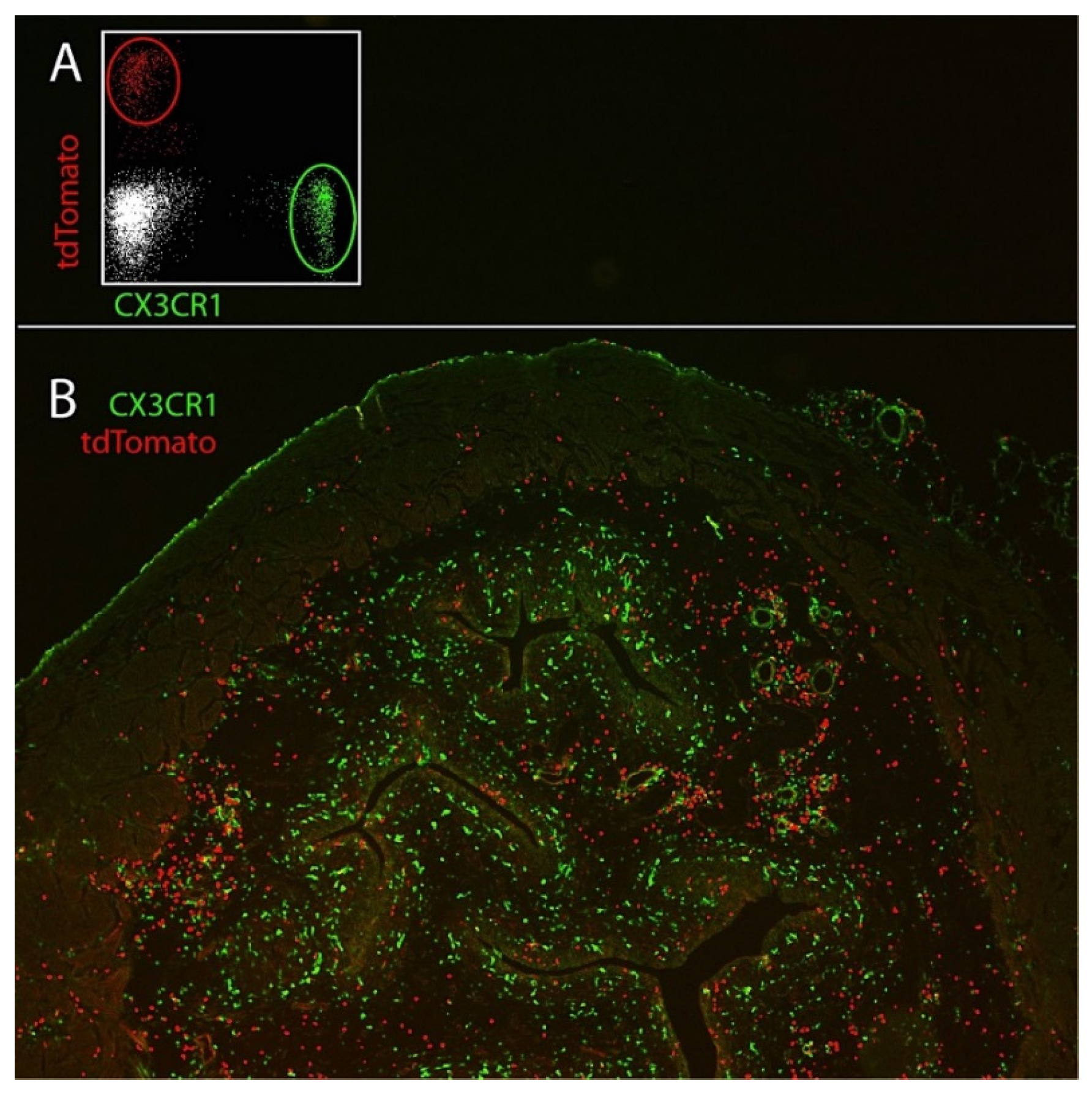
2.1. Neutrophils Internalize Uropathogenic E. coli
2.2. TNF-Dependent Neutrophil Transepithelial Migration
2.3. Ly6C− Macrophages Produce CXCL2
2.4. Neutrophil Migration across Epithelium Depends on MMP-9
2.5. Three-Cellular Crosstalk between Ly6C− and Ly6C+ Macrophages with Neutrophils
3. Experimental Section
3.1. Mice
3.2. UPEC Model
3.3. Histology
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chin, A.C.; Parkos, C.A. Neutrophil transepithelial migration and epithelial barrier function in IBD: Potential targets for inhibiting neutrophil trafficking. Ann. N. Y. Acad. Sci. 2006, 1072, 276–287. [Google Scholar] [PubMed]
- Quinones, M.P.; Kalkonde, Y.; Estrada, C.A.; Jimenez, F.; Ramirez, R.; Mahimainathan, L.; Mummidi, S.; Choudhury, G.G.; Martinez, H.; Adams, L.; et al. Role of astrocytes and chemokine systems in acute TNFalpha induced demyelinating syndrome: CCR2-dependent signals promote astrocyte activation and survival via NF-kappaB and Akt. Mol. Cell. Neurosci. 2008, 37, 96–109. [Google Scholar] [PubMed]
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Nourshargh, S.; Hordijk, P.L.; Sixt, M. Breaching multiple barriers: Leukocyte motility through venular walls and the interstitium. Nature reviews. Mol. Cell. Biol. 2010, 11, 366–378. [Google Scholar]
- Voisin, M.B.; Probstl, D.; Nourshargh, S. Venular basement membranes ubiquitously express matrix protein low-expression regions: Characterization in multiple tissues and remodeling during inflammation. Am. J. Pathol. 2010, 176, 482–495. [Google Scholar] [PubMed]
- Wang, S.; Voisin, M.B.; Larbi, K.Y.; Dangerfield, J.; Scheiermann, C.; Tran, M.; Maxwell, P.H.; Sorokin, L.; Nourshargh, S. Venular basement membranes contain specific matrix protein low expression regions that act as exit points for emigrating neutrophils. J. Exp. Med. 2006, 203, 1519–1532. [Google Scholar] [PubMed]
- Ichiyasu, H.; McCormack, J.M.; McCarthy, K.M.; Dombkowski, D.; Preffer, F.I.; Schneeberger, E.E. Matrix metalloproteinase-9-deficient dendritic cells have impaired migration through tracheal epithelial tight junctions. Am. J. Respir. Cell Mol. Biol. 2004, 30, 761–770. [Google Scholar] [PubMed]
- LeBleu, V.S.; Macdonald, B.; Kalluri, R. Structure and function of basement membranes. Exp. Biol. Med. (Maywood) 2007, 232, 1121–1129. [Google Scholar] [PubMed]
- Lammermann, T.; Bader, B.L.; Monkley, S.J.; Worbs, T.; Wedlich-Soldner, R.; Hirsch, K.; Keller, M.; Forster, R.; Critchley, D.R.; Fassler, R.; et al. Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature 2008, 453, 51–55. [Google Scholar] [PubMed]
- Shulman, Z.; Shinder, V.; Klein, E.; Grabovsky, V.; Yeger, O.; Geron, E.; Montresor, A.; Bolomini-Vittori, M.; Feigelson, S.W.; Kirchhausen, T.; et al. Lymphocyte crawling and transendothelial migration require chemokine triggering of high-affinity LFA-1 integrin. Immunity 2009, 30, 384–396. [Google Scholar] [PubMed]
- Chabot, V.; Reverdiau, P.; Iochmann, S.; Rico, A.; Senecal, D.; Goupille, C.; Sizaret, P.Y.; Sensebe, L. CCL5-enhanced human immature dendritic cell migration through the basement membrane in vitro depends on matrix metalloproteinase-9. J. Leukoc. Biol. 2006, 79, 767–778. [Google Scholar] [PubMed]
- Chakrabarti, S.; Patel, K.D. Regulation of matrix metalloproteinase-9 release from IL-8-stimulated human neutrophils. J. Leukoc. Biol. 2005, 78, 279–288. [Google Scholar] [PubMed]
- Luca, M.; Huang, S.; Gershenwald, J.E.; Singh, R.K.; Reich, R.; Bar-Eli, M. Expression of interleukin-8 by human melanoma cells up-regulates MMP-2 activity and increases tumor growth and metastasis. Am. J. Pathol. 1997, 151, 1105–1113. [Google Scholar] [PubMed]
- Van den Steen, P.E.; Dubois, B.; Nelissen, I.; Rudd, P.M.; Dwek, R.A.; Opdenakker, G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9). Crit. Rev. Biochem. Mol. Biol. 2002, 37, 375–536. [Google Scholar] [PubMed]
- Benoit, M.; Desnues, B.; Mege, J.L. Macrophage polarization in bacterial infections. J. Immunol. 2008, 181, 3733–3739. [Google Scholar] [PubMed]
- Kopydlowski, K.M.; Salkowski, C.A.; Cody, M.J.; van Rooijen, N.; Major, J.; Hamilton, T.A.; Vogel, S.N. Regulation of macrophage chemokine expression by lipopolysaccharide in vitro and in vivo. J. Immunol. 1999, 163, 1537–1544. [Google Scholar] [PubMed]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Soehnlein, O.; Lindbom, L. Phagocyte partnership during the onset and resolution of inflammation. Nat. Rev. Immunol. 2010, 10, 427–439. [Google Scholar] [PubMed]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science 2010, 330, 841–845. [Google Scholar] [PubMed]
- Schulz, C.; Gomez Perdiguero, E.; Chorro, L.; Szabo-Rogers, H.; Cagnard, N.; Kierdorf, K.; Prinz, M.; Wu, B.; Jacobsen, S.E.; Pollard, J.W.; et al. A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science 2012, 336, 86–90. [Google Scholar] [PubMed]
- Serbina, N.V.; Pamer, E.G. Monocyte emigration from bone marrow during bacterial infection requires signals mediated by chemokine receptor CCR2. Nat. Immunol. 2006, 7, 311–317. [Google Scholar] [PubMed]
- Landsman, L.; Varol, C.; Jung, S. Distinct differentiation potential of blood monocyte subsets in the lung. J. Immunol. 2007, 178, 2000–2007. [Google Scholar] [PubMed]
- Foxman, B. Epidemiology of urinary tract infections: Incidence, morbidity, and economic costs. Am. J. Med. 2002, 113, 5–13. [Google Scholar] [CrossRef]
- Mulvey, M.A.; Lopez-Boado, Y.S.; Wilson, C.L.; Roth, R.; Parks, W.C.; Heuser, J.; Hultgren, S.J. Induction and evasion of host defenses by type 1-piliated uropathogenic Escherichia coli. Science 1998, 282, 1494–1497. [Google Scholar] [PubMed]
- Martinez, J.J.; Mulvey, M.A.; Schilling, J.D.; Pinkner, J.S.; Hultgren, S.J. Type 1 pilus-mediated bacterial invasion of bladder epithelial cells. EMBO J. 2000, 19, 2803–2812. [Google Scholar] [PubMed]
- Kurts, C.; Panzer, U.; Anders, H.J.; Rees, A.J. The immune system and kidney disease: Basic concepts and clinical implications. Nat. Rev. Immunol. 2013, 13, 738–753. [Google Scholar] [PubMed]
- Svensson, M.; Yadav, M.; Holmqvist, B.; Lutay, N.; Svanborg, C.; Godaly, G. Acute pyelonephritis and renal scarring are caused by dysfunctional innate immunity in mCxcr2 heterozygous mice. Kidney Int. 2011, 80, 1064–1072. [Google Scholar] [PubMed]
- Frendeus, B.; Godaly, G.; Hang, L.; Karpman, D.; Lundstedt, A.C.; Svanborg, C. Interleukin 8 receptor deficiency confers susceptibility to acute experimental pyelonephritis and may have a human counterpart. J. Exp. Med. 2000, 192, 881–890. [Google Scholar] [PubMed]
- Belge, K.U.; Dayyani, F.; Horelt, A.; Siedlar, M.; Frankenberger, M.; Frankenberger, B.; Espevik, T.; Ziegler-Heitbrock, L. The proinflammatory CD14+CD16+DR++ monocytes are a major source of TNF. J. Immunol. 2002, 168, 3536–3542. [Google Scholar] [PubMed]
- Engel, D.R.; Maurer, J.; Tittel, A.P.; Weisheit, C.; Cavlar, T.; Schumak, B.; Limmer, A.; van Rooijen, N.; Trautwein, C.; Tacke, F.; Kurts, C. CCR2 mediates homeostatic and inflammatory release of Gr1(high) monocytes from the bone marrow, but is dispensable for bladder infiltration in bacterial urinary tract infection. J. Immunol. 2008, 181, 5579–5586. [Google Scholar] [PubMed]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [PubMed]
- Schiwon, M.; Weisheit, C.; Franken, L.; Gutweiler, S.; Dixit, A.; Meyer-Schwesinger, C.; Pohl, J.M.; Maurice, N.J.; Thiebes, S.; Lorenz, K.; et al. Crosstalk between Sentinel and Helper Macrophages Permits Neutrophil Migration into Infected Uroepithelium. Cell 2014, 156, 456–468. [Google Scholar] [PubMed]
- Jung, S.; Aliberti, J.; Graemmel, P.; Sunshine, M.J.; Kreutzberg, G.W.; Sher, A.; Littman, D.R. Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion. Mol. Cell. Biol. 2000, 20, 4106–4114. [Google Scholar] [PubMed]
- Hasenberg, A.; Hasenberg, M.; Mann, L.; Neumann, F.; Borkenstein, L.; Stecher, M.; Kraus, A.; Engel, D.R.; Klingberg, A.; Seddigh, P.; et al. Catchup: A mouse model for imaging-based tracking and modulation of neutrophil granulocytes. Nat. Methods 2015.
- Pfeffer, K.; Matsuyama, T.; Kundig, T.M.; Wakeham, A.; Kishihara, K.; Shahinian, A.; Wiegmann, K.; Ohashi, P.S.; Kronke, M.; Mak, T.W. Mice deficient for the 55 kd tumor necrosis factor receptor are resistant to endotoxic shock, yet succumb to L. monocytogenes infection. Cell 1993, 73, 457–467. [Google Scholar] [PubMed]
- Flynn, J.L.; Goldstein, M.M.; Chan, J.; Triebold, K.J.; Pfeffer, K.; Lowenstein, C.J.; Schreiber, R.; Mak, T.W.; Bloom, B.R. Tumor necrosis factor-alpha is required in the protective immune response against Mycobacterium tuberculosis in mice. Immunity 1995, 2, 561–572. [Google Scholar] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zec, K.; Volke, J.; Vijitha, N.; Thiebes, S.; Gunzer, M.; Kurts, C.; Engel, D.R. Neutrophil Migration into the Infected Uroepithelium Is Regulated by the Crosstalk between Resident and Helper Macrophages. Pathogens 2016, 5, 15. https://doi.org/10.3390/pathogens5010015
Zec K, Volke J, Vijitha N, Thiebes S, Gunzer M, Kurts C, Engel DR. Neutrophil Migration into the Infected Uroepithelium Is Regulated by the Crosstalk between Resident and Helper Macrophages. Pathogens. 2016; 5(1):15. https://doi.org/10.3390/pathogens5010015
Chicago/Turabian StyleZec, Kristina, Julia Volke, Nirojah Vijitha, Stephanie Thiebes, Matthias Gunzer, Christian Kurts, and Daniel Robert Engel. 2016. "Neutrophil Migration into the Infected Uroepithelium Is Regulated by the Crosstalk between Resident and Helper Macrophages" Pathogens 5, no. 1: 15. https://doi.org/10.3390/pathogens5010015
APA StyleZec, K., Volke, J., Vijitha, N., Thiebes, S., Gunzer, M., Kurts, C., & Engel, D. R. (2016). Neutrophil Migration into the Infected Uroepithelium Is Regulated by the Crosstalk between Resident and Helper Macrophages. Pathogens, 5(1), 15. https://doi.org/10.3390/pathogens5010015