Animal Models for Influenza Viruses: Implications for Universal Vaccine Development



Abstract
:1. Introduction
1.1. The Virus
Influenza
1.2. Clinical Relevance and Control Measures for Influenza Virus Infections
1.3. Animal Models Used in Influenza Virus Research
2. Pathogenesis of the Influenza Virus
2.1. Ferrets
2.2. Mice

| Clinical Signs | Animal Model | |||
|---|---|---|---|---|
| Ferret | Mouse | Pig | Nonhuman Primates | |
| Nasal discharge | YES | NOT OBSERVED | YES | YES |
| Coughing/sneezing | YES (sneezing) | NOT OBSERVED | OCCASIONALLY - intensity varies with the strain - | YES - only upon infection with highly pathogenic viruses - |
| Malaise | YES | YES | NOT OBSERVED | YES |
| Fever | YES | NO - develop hypothermia - | OCCASIONALLY - intensity varies with the strain - | OCCASIONALLY - only upon infection with highly pathogenic viruses - |
| Anorexia - weight loss - | YES - more predominant for highly pathogenic viruses - | YES | MINOR | MINOR |
| Neurological complications | OCCASIONALLY - high path avian viruses - | OCCASIONALLY - high path avian viruses - | NOT OBSERVED | NOT OBSERVED |
| Hypercytokenia - cytokine storm - | YES - in some cases of highly virulent strains - | YES - upon infection with highly pathogenic viruses - | LIMITED | YES |
2.3. Pigs
2.4. Nonhuman Primates
2.5. Other Animal Models
3. Evaluation of Broadly Protective Influenza Vaccines in Animal Models
| Species | Advantages | Disadvantages |
|---|---|---|
| Mice |
|
|
| Ferret |
|
|
| Pig |
|
|
| NHP |
|
|
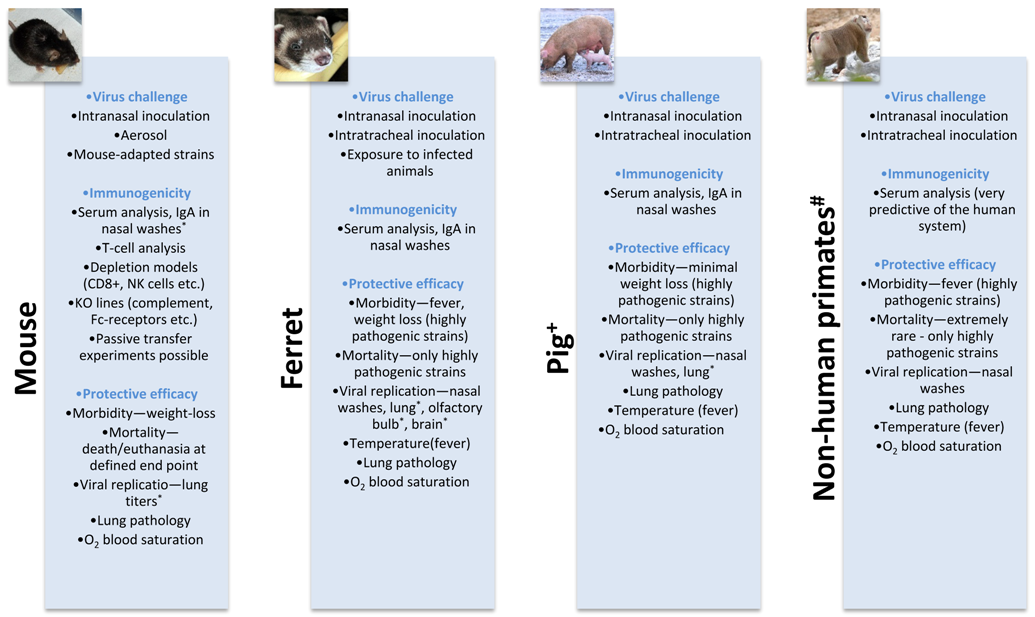
3.1. Mice
3.2. Ferrets
3.3. Pigs
3.4. Nonhuman Primates
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shaw, M.L.; Palese, P. Orthomyxoviridae: The viruses and their replication. In Fields Virology,, 6th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 1647–1689. [Google Scholar]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.; Chen, L.M.; Recuenco, S.; Ellison, J.A.; Davis, C.T.; York, I.A.; et al. A distinct lineage of influenza a virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza a viruses. PLoS Pathog 2013, 9, e1003657. [Google Scholar] [CrossRef] [PubMed]
- Cox, N.J.; Subbarao, K. Global epidemiology of influenza: Past and present. Annu. Rev. Med. 2000, 51, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, N.M.; Palese, P. The biology of influenza viruses. Vaccine 2008, 26, D49–D53. [Google Scholar] [CrossRef] [PubMed]
- Beigel, J.H.; Farrar, J.; Han, A.M.; Hayden, F.G.; Hyer, R.; de Jong, M.D.; Lochindarat, S.; Nguyen, T.K.; Nguyen, T.H.; Tran, T.H.; et al. Avian influenza a (H5N1) infection in humans. N. Engl. J. Med. 2005, 353, 1374–1385. [Google Scholar] [CrossRef] [PubMed]
- Butt, K.M.; Smith, G.J.; Chen, H.; Zhang, L.J.; Leung, Y.H.; Xu, K.M.; Lim, W.; Webster, R.G.; Yuen, K.Y.; Peiris, J.S.; et al. Human infection with an avian H9N2 influenza a virus in hong kong in 2003. J. Clin. Microbiol. 2005, 43, 5760–5767. [Google Scholar] [CrossRef] [PubMed]
- Aamir, U.B.; Naeem, K.; Ahmed, Z.; Obert, C.A.; Franks, J.; Krauss, S.; Seiler, P.; Webster, R.G. Zoonotic potential of highly pathogenic avian H7N3 influenza viruses from pakistan. Virology 2009, 390, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Munster, V.J.; de Wit, E.; van Riel, D.; Beyer, W.E.; Rimmelzwaan, G.F.; Osterhaus, A.D.; Kuiken, T.; Fouchier, R.A. The molecular basis of the pathogenicity of the dutch highly pathogenic human influenza a H7N7 viruses. J. Infect. Dis. 2007, 196, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.H.; Yang, J.R.; Wu, H.S.; Chang, M.C.; Lin, J.S.; Lin, C.Y.; Liu, Y.L.; Lo, Y.C.; Yang, C.H.; Chuang, J.H.; et al. Human infection with avian influenza a h6n1 virus: An epidemiological analysis. Lancet Respir. Med. 2013, 1, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Palese, P. Universal influenza virus vaccines: Need for clinical trials. Nature Immunol. 2013, 15, 3–5. [Google Scholar] [CrossRef]
- To, K.K.; Tsang, A.K.; Chan, J.F.; Cheng, V.C.; Chen, H.; Yuen, K.Y. Emergence in china of human disease due to avian influenza a(H10N8)—cause for concern? J. Infect. 2014, 68, 205–215. [Google Scholar] [CrossRef]
- Bouvier, N.M.; Lowen, A.C. Animal models for influenza virus pathogenesis and transmission. Viruses 2010, 2, 1530–1563. [Google Scholar] [PubMed]
- Connor, R.J.; Kawaoka, Y.; Webster, R.G.; Paulson, J.C. Receptor specificity in human, avian, and equine h2 and h3 influenza virus isolates. Virology 1994, 205, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Baum, L.G.; Paulson, J.C. Sialyloligosaccharides of the respiratory epithelium in the selection of human influenza virus receptor specificity. Acta Histochem. Suppl. 1990, 40, 35–38. [Google Scholar] [PubMed]
- Yu, H.; Gao, Z.; Feng, Z.; Shu, Y.; Xiang, N.; Zhou, L.; Huai, Y.; Feng, L.; Peng, Z.; Li, Z.; et al. Clinical characteristics of 26 human cases of highly pathogenic avian influenza a (H5N1) virus infection in china. PLoS One 2008, 3, e2985. [Google Scholar] [CrossRef] [PubMed]
- Rimmelzwaan, G.F.; Kuiken, T.; van Amerongen, G.; Bestebroer, T.M.; Fouchier, R.A.; Osterhaus, A.D. A primate model to study the pathogenesis of influenza a (H5N1) virus infection. Avian Dis. 2003, 47, 931–933. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.Y.; Poon, L.L.; Lau, A.S.; Luk, W.; Lau, Y.L.; Shortridge, K.F.; Gordon, S.; Guan, Y.; Peiris, J.S. Induction of proinflammatory cytokines in human macrophages by influenza a (H5N1) viruses: A mechanism for the unusual severity of human disease? Lancet 2002, 360, 1831–1837. [Google Scholar] [CrossRef]
- Korteweg, C.; Gu, J. Pathology, molecular biology, and pathogenesis of avian influenza a (H5N1) infection in humans. Am. J. Pathol. 2008, 172, 1155–1170. [Google Scholar] [CrossRef] [PubMed]
- Palese, P. Influenza: Old and new threats. Nat. Med. 2004, 10, S82–S87. [Google Scholar] [CrossRef] [PubMed]
- WHO. Influenza (seasonal). Available online: http://www.who.int/mediacentre/factsheets/fs211/en (accessed on 10 October 2014).
- Monto, A.S. Epidemiology and virology of influenza illness. Am. J. Manag. Care 2000, 6, S255–S264. [Google Scholar]
- Tamerius, J.; Nelson, M.I.; Zhou, S.Z.; Viboud, C.; Miller, M.A.; Alonso, W.J. Global influenza seasonality: Reconciling patterns across temperate and tropical regions. Environ. Health Perspect. 2011, 119, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Lowen, A.; Palese, P. Transmission of influenza virus in temperate zones is predominantly by aerosol, in the tropics by contact: A hypothesis. PLoS Curr. 2009, 1, RRN1002. [Google Scholar] [CrossRef] [PubMed]
- Lowen, A.C.; Steel, J.; Mubareka, S.; Palese, P. High temperature (30 degrees c) blocks aerosol but not contact transmission of influenza virus. J. Virol. 2008, 82, 5650–5652. [Google Scholar] [CrossRef] [PubMed]
- Ison, M.G. Clinical use of approved influenza antivirals: Therapy and prophylaxis. Influ. Other Respir. Viruses 2013, 7, 7–13. [Google Scholar] [CrossRef]
- Kamali, A.; Holodniy, M. Influenza treatment and prophylaxis with neuraminidase inhibitors: A review. Infect. Drug Resist. 2013, 6, 187–198. [Google Scholar] [PubMed]
- Hsu, J.; Santesso, N.; Mustafa, R.; Brozek, J.; Chen, Y.L.; Hopkins, J.P.; Cheung, A.; Hovhannisyan, G.; Ivanova, L.; Flottorp, S.A.; et al. Antivirals for treatment of influenza: A systematic review and meta-analysis of observational studies. Ann. Intern. Med. 2012, 156, 512–524. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E. Antiviral agents active against influenza a viruses. Nat. Rev. Drug Discov. 2006, 5, 1015–1025. [Google Scholar]
- Nichol, K.L. Efficacy and effectiveness of influenza vaccination. Vaccine 2008, 26, D17–D22. [Google Scholar] [CrossRef] [PubMed]
- Palese, P. Making better influenza virus vaccines? Emerg. Infect. Dis. 2006, 12, 61–65. [Google Scholar] [CrossRef]
- Barr, I.G.; Jelley, L.L. The coming era of quadrivalent human influenza vaccines: Who will benefit? Drugs 2012, 72, 2177–2185. [Google Scholar] [CrossRef]
- Ambrose, C.S.; Levin, M.J. The rationale for quadrivalent influenza vaccines. Hum. Vaccin. Immunother. 2012, 8, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Wilde, J.A.; McMillan, J.A.; Serwint, J.; Butta, J.; O'Riordan, M.A.; Steinhoff, M.C. Effectiveness of influenza vaccine in health care professionals: A randomized trial. JAMA: J. Am. Med. Assoc. 1999, 281, 908–913. [Google Scholar] [CrossRef]
- Belshe, R.B.; Mendelman, P.M.; Treanor, J.; King, J.; Gruber, W.C.; Piedra, P.; Bernstein, D.I.; Hayden, F.G.; Kotloff, K.; Zangwill, K.; et al. The efficacy of live attenuated, cold-adapted, trivalent, intranasal influenzavirus vaccine in children. N. Engl. J. Med. 1998, 338, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Yohannes, K.; Roche, P.; Spencer, J.; Hampson, A. Annual report of the national influenza surveillance scheme, 2002. Commun. Dis. Intell. Q. Rep. 2003, 27, 162–172. [Google Scholar]
- Bridges, C.B.; Thompson, W.W.; Meltzer, M.I.; Reeve, G.R.; Talamonti, W.J.; Cox, N.J.; Lilac, H.A.; Hall, H.; Klimov, A.; Fukuda, K. Effectiveness and cost-benefit of influenza vaccination of healthy working adults: A randomized controlled trial. JAMA: J. Am. Med. Assoc. 2000, 284, 1655–1663. [Google Scholar] [CrossRef]
- Borse, R.H.; Shrestha, S.S.; Fiore, A.E.; Atkins, C.Y.; Singleton, J.A.; Furlow, C.; Meltzer, M.I. Effects of vaccine program against pandemic influenza a (H1N1) virus, united states, 2009–2010. Emerg. Infect. Dis. 2013, 19, 439–448. [Google Scholar] [CrossRef]
- Pica, N.; Palese, P. Toward a universal influenza virus vaccine: Prospects and challenges. Annu. Rev. Med. 2013, 64, 189–202. [Google Scholar] [CrossRef]
- Krammer, F.; Palese, P. Influenza virus hemagglutinin stalk-based antibodies and vaccines. Current opinion in virology 2013, 3, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.; Manch, M.D.; Andrews, C.H.; Lond, M.D.; Laidlaw, P.P. A virus obtained from influenza patients. Lancet 1933, 222, 66–68. [Google Scholar] [CrossRef]
- The virus of influenza. Lancet 1933, 222, 83.
- MacInnes, H.; Zhou, Y.; Gouveia, K.; Cromwell, J.; Lowery, K.; Layton, R.C.; Zubelewicz, M.; Sampath, R.; Hofstadler, S.; Liu, Y.; et al. Transmission of aerosolized seasonal H1N1 influenza a to ferrets. PLoS One 2011, 6, e24448. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.M.; Song, B.M.; Lee, J.S.; Kim, H.S.; Seo, S.H. Pandemic H1N1 influenza virus causes a stronger inflammatory response than seasonal H1N1 influenza virus in ferrets. Arch. Virol. 2011, 156, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Banner, D.; Fang, Y.; Ng, D.C.; Kanagasabai, T.; Kelvin, D.J.; Kelvin, A.A. Comparative analyses of pandemic H1N1 and seasonal H1N1, H3N2, and influenza b infections depict distinct clinical pictures in ferrets. PLoS One 2011, 6, e27512. [Google Scholar] [PubMed]
- Meunier, I.; Embury-Hyatt, C.; Stebner, S.; Gray, M.; Bastien, N.; Li, Y.; Plummer, F.; Kobinger, G.P.; von Messling, V. Virulence differences of closely related pandemic 2009 H1N1 isolates correlate with increased inflammatory responses in ferrets. Virology 2012, 422, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Van den Brand, J.M.; Stittelaar, K.J.; van Amerongen, G.; Reperant, L.; de Waal, L.; Osterhaus, A.D.; Kuiken, T. Comparison of temporal and spatial dynamics of seasonal H3N2, pandemic H1N1 and highly pathogenic avian influenza H5N1 virus infections in ferrets. PLoS One 2012, 7, e42343. [Google Scholar]
- Malakhov, M.P.; Aschenbrenner, L.M.; Smee, D.F.; Wandersee, M.K.; Sidwell, R.W.; Gubareva, L.V.; Mishin, V.P.; Hayden, F.G.; Kim, D.H.; Ing, A.; et al. Sialidase fusion protein as a novel broad-spectrum inhibitor of influenza virus infection. Antimicrob. Agents Chemother. 2006, 50, 1470–1479. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.L.; Lamirande, E.W.; Yang, C.F.; Jin, H.; Kemble, G.; Subbarao, K. Evaluation of replication and cross-reactive antibody responses of h2 subtype influenza viruses in mice and ferrets. J. Virol. 2010, 84, 7695–7702. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.; Lee, D.H.; Park, J.K.; Yuk, S.S.; Kwon, J.H.; Nahm, S.S.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Experimental infection and natural contact exposure of ferrets with canine influenza virus (H3N2). J. Gen. Virol. 2013, 94, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Govorkova, E.A.; Rehg, J.E.; Krauss, S.; Yen, H.L.; Guan, Y.; Peiris, M.; Nguyen, T.D.; Hanh, T.H.; Puthavathana, P.; Long, H.T.; et al. Lethality to ferrets of h5n1 influenza viruses isolated from humans and poultry in 2004. J. Virol. 2005, 79, 2191–2198. [Google Scholar] [PubMed]
- Xu, Q.; Chen, Z.; Cheng, X.; Xu, L.; Jin, H. Evaluation of live attenuated H7N3 and H7N7 vaccine viruses for their receptor binding preferences, immunogenicity in ferrets and cross reactivity to the novel H7N9 virus. PLoS One 2013, 8, e76884. [Google Scholar] [CrossRef] [PubMed]
- Kimble, J.B.; Sorrell, E.; Shao, H.; Martin, P.L.; Perez, D.R. Compatibility of H9N2 avian influenza surface genes and 2009 pandemic H1N1 internal genes for transmission in the ferret model. Proc. Natl. Acad. Sci. USA 2011, 108, 12084–12088. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Gustin, K.M.; Pearce, M.B.; Maines, T.R.; Zeng, H.; Pappas, C.; Sun, X.; Carney, P.J.; Villanueva, J.M.; Stevens, J.; et al. Pathogenesis and transmission of avian influenza a (H7N9) virus in ferrets and mice. Nature 2013, 501, 556–559. [Google Scholar] [CrossRef] [PubMed]
- Richard, M.; Schrauwen, E.J.; de Graaf, M.; Bestebroer, T.M.; Spronken, M.I.; van Boheemen, S.; de Meulder, D.; Lexmond, P.; Linster, M.; Herfst, S.; et al. Limited airborne transmission of H7N9 influenza a virus between ferrets. Nature 2013, 501, 560–563. [Google Scholar] [PubMed]
- Van Riel, D.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.; Osterhaus, A.D.; Kuiken, T. Human and avian influenza viruses target different cells in the lower respiratory tract of humans and other mammals. Am. J. Pathol. 2007, 171, 1215–1223. [Google Scholar]
- Stark, G.V.; Long, J.P.; Ortiz, D.I.; Gainey, M.; Carper, B.A.; Feng, J.; Miller, S.M.; Bigger, J.E.; Vela, E.M. Clinical profiles associated with influenza disease in the ferret model. PLoS One 2013, 8, e58337. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Banner, D.; Degousee, N.; Leon, A.J.; Xu, L.; Paquette, S.G.; Kanagasabai, T.; Fang, Y.; Rubino, S.; Rubin, B.; et al. Differential pathological and immune responses in newly weaned ferrets are associated with a mild clinical outcome of pandemic 2009 H1N1 infection. J. Virol. 2012, 86, 13187–13201. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Kreijtz, J.H.; van Amerongen, G.; Fouchier, R.A.; Osterhaus, A.D.; Rimmelzwaan, G.F.; Kuiken, T. Pathogenesis of influenza a/H5N1 virus infection in ferrets differs between intranasal and intratracheal routes of inoculation. Am. J. Pathol. 2011, 179, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Rimmelzwaan, G.F.; Osterhaus, A.D. Animal models for the preclinical evaluation of candidate influenza vaccines. Expert Rev. Vaccines 2010, 9, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Zitzow, L.A.; Rowe, T.; Morken, T.; Shieh, W.J.; Zaki, S.; Katz, J.M. Pathogenesis of avian influenza a (H5N1) viruses in ferrets. J. Virol. 2002, 76, 4420–4429. [Google Scholar] [CrossRef] [PubMed]
- Van Riel, D.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.; Osterhaus, A.D.; Kuiken, T. H5N1 virus attachment to lower respiratory tract. Science 2006, 312, 399. [Google Scholar]
- Herlocher, M.L.; Truscon, R.; Elias, S.; Yen, H.L.; Roberts, N.A.; Ohmit, S.E.; Monto, A.S. Influenza viruses resistant to the antiviral drug oseltamivir: Transmission studies in ferrets. J. Infect. Dis. 2004, 190, 1627–1630. [Google Scholar] [CrossRef] [PubMed]
- Maines, T.R.; Jayaraman, A.; Belser, J.A.; Wadford, D.A.; Pappas, C.; Zeng, H.; Gustin, K.M.; Pearce, M.B.; Viswanathan, K.; Shriver, Z.H.; et al. Transmission and pathogenesis of swine-origin 2009 a (H1N1) influenza viruses in ferrets and mice. Science 2009, 325, 484–487. [Google Scholar]
- Staeheli, P.; Grob, R.; Meier, E.; Sutcliffe, J.G.; Haller, O. Influenza virus-susceptible mice carry mx genes with a large deletion or a nonsense mutation. Mol. Cell. Biol. 1988, 8, 4518–4523. [Google Scholar] [PubMed]
- Grimm, D.; Staeheli, P.; Hufbauer, M.; Koerner, I.; Martinez-Sobrido, L.; Solorzano, A.; Garcia-Sastre, A.; Haller, O.; Kochs, G. Replication fitness determines high virulence of influenza a virus in mice carrying functional mx1 resistance gene. Proc. Natl. Acad. Sci. USA 2007, 104, 6806–6811. [Google Scholar] [CrossRef] [PubMed]
- Narasaraju, T.; Sim, M.K.; Ng, H.H.; Phoon, M.C.; Shanker, N.; Lal, S.K.; Chow, V.T. Adaptation of human influenza H3N2 virus in a mouse pneumonitis model: Insights into viral virulence, tissue tropism and host pathogenesis. Microbes Infect./Inst. Pasteur 2009, 11, 2–11. [Google Scholar] [CrossRef]
- Sakabe, S.; Ozawa, M.; Takano, R.; Iwastuki-Horimoto, K.; Kawaoka, Y. Mutations in pa, np, and ha of a pandemic (H1N1) 2009 influenza virus contribute to its adaptation to mice. Virus Res. 2011, 158, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Chen, H.; Huang, J.; Chen, Y.; Gu, M.; Wang, X.; Hu, S.; Liu, X.; Liu, X. A nonpathogenic duck-origin H9N2 influenza a virus adapts to high pathogenicity in mice. Arch. Virol. 2014. [Google Scholar] [CrossRef]
- Medina, R.A.; Manicassamy, B.; Stertz, S.; Seibert, C.W.; Hai, R.; Belshe, R.B.; Frey, S.E.; Basler, C.F.; Palese, P.; Garcia-Sastre, A. Pandemic 2009 H1N1 vaccine protects against 1918 spanish influenza virus. Nat. Commun. 2010, 1, 28. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Watanabe, S.; Ito, T.; Goto, H.; Wells, K.; McGregor, M.; Cooley, A.J.; Kawaoka, Y. Biological heterogeneity, including systemic replication in mice, of H5N1 influenza a virus isolates from humans in hong kong. J. Virol. 1999, 73, 3184–3189. [Google Scholar] [PubMed]
- Lu, X.; Tumpey, T.M.; Morken, T.; Zaki, S.R.; Cox, N.J.; Katz, J.M. A mouse model for the evaluation of pathogenesis and immunity to influenza a (H5N1) viruses isolated from humans. J. Virol. 1999, 73, 5903–5911. [Google Scholar] [PubMed]
- Xu, L.; Bao, L.; Deng, W.; Zhu, H.; Chen, T.; Lv, Q.; Li, F.; Yuan, J.; Xiang, Z.; Gao, K.; et al. The mouse and ferret models for studying the novel avian-origin human influenza a (H7N9) virus. Virol. J. 2013, 10, 253. [Google Scholar] [CrossRef] [PubMed]
- Ibricevic, A.; Pekosz, A.; Walter, M.J.; Newby, C.; Battaile, J.T.; Brown, E.G.; Holtzman, M.J.; Brody, S.L. Influenza virus receptor specificity and cell tropism in mouse and human airway epithelial cells. J. Virol. 2006, 80, 7469–7480. [Google Scholar] [CrossRef] [PubMed]
- Carey, M.A.; Bradbury, J.A.; Rebolloso, Y.D.; Graves, J.P.; Zeldin, D.C.; Germolec, D.R. Pharmacologic inhibition of cox-1 and cox-2 in influenza a viral infection in mice. PLoS One 2010, 5, e11610. [Google Scholar] [CrossRef] [PubMed]
- DiLillo, D.J.; Tan, G.S.; Palese, P.; Ravetch, J.V. Broadly neutralizing hemagglutinin stalk-specific antibodies require fcgammar interactions for protection against influenza virus in vivo. Nat. Med. 2014, 20, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Lamirande, E.W.; Subbarao, K. The mouse model for influenza. Curr. Protoc. Microbiol. 2009. Chapter 15, Unit 15G 13. [Google Scholar] [CrossRef]
- Ma, W.; Lager, K.M.; Vincent, A.L.; Janke, B.H.; Gramer, M.R.; Richt, J.A. The role of swine in the generation of novel influenza viruses. Zoonoses Public Health 2009, 56, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Couceiro, J.N.; Kelm, S.; Baum, L.G.; Krauss, S.; Castrucci, M.R.; Donatelli, I.; Kida, H.; Paulson, J.C.; Webster, R.G.; et al. Molecular basis for the generation in pigs of influenza a viruses with pandemic potential. J. Virol. 1998, 72, 7367–7373. [Google Scholar] [PubMed]
- Barnard, D.L. Animal models for the study of influenza pathogenesis and therapy. Antivir. Res. 2009, 82, A110–A122. [Google Scholar] [CrossRef] [PubMed]
- Lipatov, A.S.; Kwon, Y.K.; Sarmento, L.V.; Lager, K.M.; Spackman, E.; Suarez, D.L.; Swayne, D.E. Domestic pigs have low susceptibility to H5N1 highly pathogenic avian influenza viruses. PLoS Pathog. 2008, 4, e1000102. [Google Scholar] [CrossRef] [PubMed]
- Dea, S.; Bilodeau, R.; Sauvageau, R.; Montpetit, C.; Martineau, G.P. Antigenic variant of swine influenza virus causing proliferative and necrotizing pneumonia in pigs. J. Vet. Diagn. Invest.: Off. Publ. Am. Assoc. Vet. Lab. Diagnosticians, Inc 1992, 4, 380–392. [Google Scholar] [CrossRef]
- Chen, Q.; Madson, D.; Miller, C.L.; Harris, D.L. Vaccine development for protecting swine against influenza virus. Anim. Health Res. Rev./Conf. Res. Workers Anim. Dis. 2012, 13, 181–195. [Google Scholar] [CrossRef]
- Karlsson, E.A.; Engel, G.A.; Feeroz, M.M.; San, S.; Rompis, A.; Lee, B.P.; Shaw, E.; Oh, G.; Schillaci, M.A.; Grant, R.; et al. Influenza virus infection in nonhuman primates. Emerg. Infect. Dis. 2012, 18, 1672–1675. [Google Scholar] [CrossRef] [PubMed]
- Carroll, T.D.; Matzinger, S.R.; Genesca, M.; Fritts, L.; Colon, R.; McChesney, M.B.; Miller, C.J. Interferon-induced expression of mxa in the respiratory tract of rhesus macaques is suppressed by influenza virus replication. J. Immunol. 2008, 180, 2385–2395. [Google Scholar] [CrossRef] [PubMed]
- Rimmelzwaan, G.F.; Baars, M.; van Amerongen, G.; van Beek, R.; Osterhaus, A.D. A single dose of an iscom influenza vaccine induces long-lasting protective immunity against homologous challenge infection but fails to protect cynomolgus macaques against distant drift variants of influenza a (H3N2) viruses. Vaccine 2001, 20, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Safronetz, D.; Rockx, B.; Feldmann, F.; Belisle, S.E.; Palermo, R.E.; Brining, D.; Gardner, D.; Proll, S.C.; Marzi, A.; Tsuda, Y.; et al. Pandemic swine-origin H1N1 influenza a virus isolates show heterogeneous virulence in macaques. J. Virol. 2011, 85, 1214–1223. [Google Scholar] [CrossRef] [PubMed]
- Baskin, C.R.; Bielefeldt-Ohmann, H.; Tumpey, T.M.; Sabourin, P.J.; Long, J.P.; Garcia-Sastre, A.; Tolnay, A.E.; Albrecht, R.; Pyles, J.A.; Olson, P.H.; et al. Early and sustained innate immune response defines pathology and death in nonhuman primates infected by highly pathogenic influenza virus. Proc. Natl. Acad. Sci. USA 2009, 106, 3455–3460. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kiso, M.; Fukuyama, S.; Nakajima, N.; Imai, M.; Yamada, S.; Murakami, S.; Yamayoshi, S.; Iwatsuki-Horimoto, K.; Sakoda, Y.; et al. Characterization of H7N9 influenza a viruses isolated from humans. Nature 2013, 501, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Rimmelzwaan, G.F.; Baars, M.; van Beek, R.; van Amerongen, G.; Lovgren-Bengtsson, K.; Claas, E.C.; Osterhaus, A.D. Induction of protective immunity against influenza virus in a macaque model: Comparison of conventional and iscom vaccines. J. Gen. Virol. 1997, 78, 757–765. [Google Scholar]
- Cilloniz, C.; Shinya, K.; Peng, X.; Korth, M.J.; Proll, S.C.; Aicher, L.D.; Carter, V.S.; Chang, J.H.; Kobasa, D.; Feldmann, F.; et al. Lethal influenza virus infection in macaques is associated with early dysregulation of inflammatory related genes. PLoS Pathog. 2009, 5, e1000604. [Google Scholar] [CrossRef]
- Lowen, A.C.; Mubareka, S.; Tumpey, T.M.; García-Sastre, A.; Palese, P. The guinea pig as a transmission model for human influenza viruses. Proc. Natl. Acad. Sci. USA 2006, 103, 9988–9992. [Google Scholar] [CrossRef] [PubMed]
- Lowen, A.C.; Mubareka, S.; Steel, J.; Palese, P. Influenza virus transmission is dependent on relative humidity and temperature. PLoS Pathog. 2007, 3, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Mubareka, S.; Lowen, A.C.; Steel, J.; Coates, A.L.; Garcia-Sastre, A.; Palese, P. Transmission of influenza virus via aerosols and fomites in the guinea pig model. J. Infect. Dis. 2009, 199, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Pica, N.; Chou, Y.Y.; Bouvier, N.M.; Palese, P. Transmission of influenza b viruses in the guinea pig. J. Virol. 2012, 86, 4279–4287. [Google Scholar] [CrossRef] [PubMed]
- Lowen, A.C.; Steel, J.; Mubareka, S.; Carnero, E.; García-Sastre, A.; Palese, P. Blocking interhost transmission of influenza virus by vaccination in the guinea pig model. J. Virol. 2009, 83, 2803–2818. [Google Scholar] [CrossRef]
- Seibert, C.W.; Rahmat, S.; Krause, J.C.; Eggink, D.; Albrecht, R.A.; Goff, P.H.; Krammer, F.; Duty, J.A.; Bouvier, N.M.; García-Sastre, A.; et al. Recombinant iga is sufficient to prevent influenza virus transmission in guinea pigs. J. Virol. 2013, 87, 7793–7804. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.M.; Yates, R.J.; Browse, G.; Swann, Z.; Newton, J.R.; Jessett, D.; Davis-Poynter, N.; Mumford, J.A. Comparison of hamster and pony challenge models for evaluation of effect of antigenic drift on cross protection afforded by equine influenza vaccines. Equine Vet. J. 2003, 35, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Abou-Donia, H.; Jennings, R.; Potter, C.W. Growth of influenza a viruses in hamsters. Arch. Virol. 1980, 65, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Jennings, R.; Potter, C.W. The hamster as a model for the study of immunity to influenza virus infection. Fed. Proc. 1978, 37, 2072–2073. [Google Scholar] [PubMed]
- Potter, C.W.; Jennings, R. The hamster as a model system for the study of influenza vaccines. Postgrad. Med. J. 1976, 52, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Jennings, R.; Denton, M.D.; Potter, C.W. The hamster as an experimental animal for the study of influenza. I. The role of antibody in protection. Med. Microbiol. Immunol. 1976, 162, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Jennings, R.; Phair, J.P.; Denton, M.D.; Potter, C.W. The hamster as an experimental animal for the study of influenza. Ii. The role of spleen cells in protection. Med. Microbiol. Immunol. 1976, 162, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Shinya, K.; Makino, A.; Tanaka, H.; Hatta, M.; Watanabe, T.; Le, M.Q.; Imai, H.; Kawaoka, Y. Systemic dissemination of H5N1 influenza a viruses in ferrets and hamsters after direct intragastric inoculation. J. Virol. 2011, 85, 4673–4678. [Google Scholar] [CrossRef]
- Ali, M.J.; Teh, C.Z.; Jennings, R.; Potter, C.W. Transmissibility of influenza viruses in hamsters. Arch. Virol. 1982, 72, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Eichelberger, M.C. The cotton rat as a model to study influenza pathogenesis and immunity. Viral. Immunol. 2007, 20, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Boukhvalova, M.S.; Prince, G.A.; Blanco, J.C. The cotton rat model of respiratory viral infections. Biologicals 2009, 37, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Yim, K.; Miles, B.; Zinsou, R.; Prince, G.; Boukhvalova, M. Efficacy of trivalent inactivated influenza vaccines in the cotton rat sigmodon hispidus model. Vaccine 2012, 30, 1291–1296. [Google Scholar] [CrossRef] [PubMed]
- Straight, T.M.; Ottolini, M.G.; Prince, G.A.; Eichelberger, M.C. Evidence of a cross-protective immune response to influenza a in the cotton rat model. Vaccine 2006, 24, 6264–6271. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.C.; Pletneva, L.M.; Wan, H.; Araya, Y.; Angel, M.; Oue, R.O.; Sutton, T.C.; Perez, D.R. Receptor characterization and susceptibility of cotton rats to avian and 2009 pandemic influenza virus strains. J. Virol. 2013, 87, 2036–2045. [Google Scholar] [CrossRef] [PubMed]
- Swayne, D.E.; Suarez, D.L.; Schultz-Cherry, S.; Tumpey, T.M.; King, D.J.; Nakaya, T.; Palese, P.; Garcia-Sastre, A. Recombinant paramyxovirus type 1-avian influenza-h7 virus as a vaccine for protection of chickens against influenza and newcastle disease. Avian Dis. 2003, 47, 1047–1050. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, L.A.; de Vries, R.P.; de Boer-Luijtze, E.A.; Rigter, A.; Rottier, P.J.; de Haan, C.A. A single immunization with soluble recombinant trimeric hemagglutinin protects chickens against highly pathogenic avian influenza virus H5N1. PLoS One 2010, 5, e10645. [Google Scholar] [CrossRef] [PubMed]
- Suarez, D.L.; Schultz-Cherry, S. Immunology of avian influenza virus: A review. Dev. Comp. Immunol. 2000, 24, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Pantin-Jackwood, M.J.; Suarez, D.L. Vaccination of domestic ducks against H5N1 hpai: A review. Virus Res. 2013, 178, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.M.; Quinlivan, M.; Sturgill, T.; Cullinane, A.; Horohov, D.W.; Zamarin, D.; Arkins, S.; Garcia-Sastre, A.; Palese, P. Influenza a viruses with truncated ns1 as modified live virus vaccines: Pilot studies of safety and efficacy in horses. Equine Vet. J. 2009, 41, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R.; Hannant, D.; Kydd, J.H.; Daly, J.M. Vaccination against equine influenza: Quid novi? Vaccine 2006, 24, 4047–4061. [Google Scholar] [CrossRef]
- Lee, D.H.; Bae, S.W.; Park, J.K.; Kwon, J.H.; Yuk, S.S.; Song, J.M.; Kang, S.M.; Kwon, Y.K.; Kim, H.Y.; Song, C.S. Virus-like particle vaccine protects against H3N2 canine influenza virus in dog. Vaccine 2013, 31, 3268–3273. [Google Scholar] [CrossRef] [PubMed]
- Van der Laan, J.W.; Herberts, C.; Lambkin-Williams, R.; Boyers, A.; Mann, A.J.; Oxford, J. Animal models in influenza vaccine testing. Expert Rev. Vaccines 2008, 7, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Eichelberger, M.C.; Green, M.D. Animal models to assess the toxicity, immunogenicity and effectiveness of candidate influenza vaccines. Expert Opin. Drug Metab. Toxicol. 2011, 7, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Thangavel, R.R.; Bouvier, N.M. Animal models for influenza virus pathogenesis, transmission, and immunology. J. Immunol. Methods 2014. [Google Scholar] [CrossRef]
- Watanabe, H.; Numata, K.; Ito, T.; Takagi, K.; Matsukawa, A. Innate immune response in th1- and th2-dominant mouse strains. Shock 2004, 22, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Pica, N.; Iyer, A.; Ramos, I.; Bouvier, N.M.; Fernandez-Sesma, A.; Garcia-Sastre, A.; Lowen, A.C.; Palese, P.; Steel, J. The dba.2 mouse is susceptible to disease following infection with a broad, but limited, range of influenza a and b viruses. J. Virol. 2011, 85, 12825–12829. [Google Scholar] [CrossRef] [PubMed]
- Okuno, Y.; Isegawa, Y.; Sasao, F.; Ueda, S. A common neutralizing epitope conserved between the hemagglutinins of influenza a virus h1 and h2 strains. J. Virol. 1993, 67, 2552–2558. [Google Scholar] [PubMed]
- Tan, G.S.; Krammer, F.; Eggink, D.; Kongchanagul, A.; Moran, T.M.; Palese, P. A pan-h1 anti-hemagglutinin monoclonal antibody with potent broad-spectrum efficacy in vivo. J. Virol. 2012, 86, 6179–6188. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Tan, G.S.; Hai, R.; Pica, N.; Petersen, E.; Moran, T.M.; Palese, P. Broadly protective monoclonal antibodies against h3 influenza viruses following sequential immunization with different hemagglutinins. PLoS Pathog. 2010, 6, e1000796. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.J.; Boyington, J.C.; McTamney, P.M.; Kong, W.P.; Pearce, M.B.; Xu, L.; Andersen, H.; Rao, S.; Tumpey, T.M.; Yang, Z.Y.; et al. Induction of broadly neutralizing H1N1 influenza antibodies by vaccination. Science 2010, 329, 1060–1064. [Google Scholar] [CrossRef] [PubMed]
- Pica, N.; Hai, R.; Krammer, F.; Wang, T.T.; Maamary, J.; Eggink, D.; Tan, G.S.; Krause, J.C.; Moran, T.; Stein, C.R.; et al. Hemagglutinin stalk antibodies elicited by the 2009 pandemic influenza virus as a mechanism for the extinction of seasonal H1N1 viruses. Proc. Natl. Acad. Sci. USA 2012, 109, 2573–2578. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Pica, N.; Hai, R.; Tan, G.S.; Palese, P. Hemagglutinin stalk-reactive antibodies are boosted following sequential infection with seasonal and pandemic H1N1 influenza virus in mice. J. Virol. 2012, 86, 10302–10307. [Google Scholar] [CrossRef] [PubMed]
- Wrammert, J.; Koutsonanos, D.; Li, G.M.; Edupuganti, S.; Sui, J.; Morrissey, M.; McCausland, M.; Skountzou, I.; Hornig, M.; Lipkin, W.I.; et al. Broadly cross-reactive antibodies dominate the human b cell response against 2009 pandemic H1N1 influenza virus infection. J. Exp. Med. 2011, 208, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Margine, I.; Hai, R.; Albrecht, R.A.; Obermoser, G.; Harrod, A.C.; Banchereau, J.; Palucka, K.; Garcia-Sastre, A.; Palese, P.; Treanor, J.J.; et al. H3N2 influenza virus infection induces broadly reactive hemagglutinin stalk antibodies in humans and mice. J. Virol. 2013, 87, 4728–4737. [Google Scholar] [CrossRef] [PubMed]
- Steel, J.; Lowen, A.C.; Wang, T.T.; Yondola, M.; Gao, Q.; Haye, K.; Garcia-Sastre, A.; Palese, P. Influenza virus vaccine based on the conserved hemagglutinin stalk domain. mBio 2010. [Google Scholar] [CrossRef]
- Wei, C.J.; Yassine, H.M.; McTamney, P.M.; Gall, J.G.; Whittle, J.R.; Boyington, J.C.; Nabel, G.J. Elicitation of broadly neutralizing influenza antibodies in animals with previous influenza exposure. Sci. Transl. Med. 2012, 4, 147ra114. [Google Scholar] [CrossRef] [PubMed]
- Kanekiyo, M.; Wei, C.J.; Yassine, H.M.; McTamney, P.M.; Boyington, J.C.; Whittle, J.R.; Rao, S.S.; Kong, W.P.; Wang, L.; Nabel, G.J. Self-assembling influenza nanoparticle vaccines elicit broadly neutralizing H1N1 antibodies. Nature 2013, 499, 102–106. [Google Scholar] [PubMed]
- Krammer, F.; Margine, I.; Hai, R.; Flood, A.; Hirsh, A.; Tsvetnitsky, V.; Chen, D.; Palese, P. H3 stalk-based chimeric hemagglutinin influenza virus constructs protect mice from H7N9 challenge. J. Virol. 2014, 88, 2340–2343. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Pica, N.; Hai, R.; Margine, I.; Palese, P. Chimeric hemagglutinin influenza virus vaccine constructs elicit broadly protective stalk-specific antibodies. J. Virol. 2013, 87, 6542–6550. [Google Scholar] [CrossRef] [PubMed]
- Margine, I.; Krammer, F.; Hai, R.; Heaton, N.S.; Tan, G.S.; Andrews, S.A.; Runstadler, J.A.; Wilson, P.C.; Albrecht, R.A.; Garcia-Sastre, A.; et al. Hemagglutinin stalk-based universal vaccine constructs protect against group 2 influenza a viruses. J. Virol. 2013, 87, 10435–10446. [Google Scholar] [CrossRef] [PubMed]
- Ekiert, D.C.; Bhabha, G.; Elsliger, M.A.; Friesen, R.H.; Jongeneelen, M.; Throsby, M.; Goudsmit, J.; Wilson, I.A. Antibody recognition of a highly conserved influenza virus epitope. Science 2009, 324, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Terajima, M.; Cruz, J.; Co, M.D.; Lee, J.H.; Kaur, K.; Wrammert, J.; Wilson, P.C.; Ennis, F.A. Complement-dependent lysis of influenza a virus-infected cells by broadly cross-reactive human monoclonal antibodies. J. Virol. 2011, 85, 13463–13467. [Google Scholar] [CrossRef] [PubMed]
- Lawrenz, M.B.; Wooten, R.M.; Zachary, J.F.; Drouin, S.M.; Weis, J.J.; Wetsel, R.A.; Norris, S.J. Effect of complement component C3 deficiency on experimental lyme borreliosis in mice. Infect. Immun. 2003, 71, 4432–4440. [Google Scholar] [CrossRef] [PubMed]
- Francis, T.; Magill, T.P. Immunological studies with the virus of influenza. J. Exp. Med. 1935, 62, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Francis, T.; Shope, R.E. Neutralization tests with sera of convalescent or immunized animals and the viruses of swine and human influenza. J. Exp. Med. 1936, 63, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Cameron, C.M.; Cameron, M.J.; Bermejo-Martin, J.F.; Ran, L.; Xu, L.; Turner, P.V.; Ran, R.; Danesh, A.; Fang, Y.; Chan, P.K.; et al. Gene expression analysis of host innate immune responses during lethal H5N1 infection in ferrets. J. Virol. 2008, 82, 11308–11317. [Google Scholar] [CrossRef] [PubMed]
- Ochi, A.; Danesh, A.; Seneviratne, C.; Banner, D.; Devries, M.E.; Rowe, T.; Xu, L.; Ran, L.; Czub, M.; Bosinger, S.E.; et al. Cloning, expression and immunoassay detection of ferret ifn-gamma. Dev. Comp. Immunol. 2008, 32, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Nakata, M.; Itou, T.; Sakai, T. Molecular cloning and phylogenetic analysis of inflammatory cytokines of the ferret (mustela putorius furo). J. Vet. Med. Sci./ Jpn. Soc. Vet. Sci. 2008, 70, 543–550. [Google Scholar] [CrossRef]
- Rutigliano, J.A.; Doherty, P.C.; Franks, J.; Morris, M.Y.; Reynolds, C.; Thomas, P.G. Screening monoclonal antibodies for cross-reactivity in the ferret model of influenza infection. J. Immunol. Methods 2008, 336, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, E.H.; Pascua, P.N.; Kwon, H.I.; Lim, G.J.; Decano, A.; Kim, S.M.; Song, M.K.; Shin, E.C.; Choi, Y.K. Evaluation of heterosubtypic cross-protection against highly pathogenic H5N1 by active infection with human seasonal influenza a virus or trivalent inactivated vaccine immunization in ferret models. J. Gen. Virol. 2014, 95, 793–798. [Google Scholar] [CrossRef] [PubMed]
- O'Donnell, C.D.; Wright, A.; Vogel, L.; Boonnak, K.; Treanor, J.J.; Subbarao, K. Humans and ferrets with prior H1N1 influenza virus infections do not exhibit evidence of original antigenic sin after infection or vaccination with the 2009 pandemic H1N1 virus. Clin. Vaccine Immunol.: CVI 2014. [Google Scholar] [CrossRef]
- Houser, K.V.; Pearce, M.B.; Katz, J.M.; Tumpey, T.M. Impact of prior seasonal H3N2 influenza vaccination or infection on protection and transmission of emerging variants of influenza a(H3N2)v virus in ferrets. J. Virol. 2013, 87, 13480–13489. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Hai, R.; Yondola, M.; Tan, G.S.; Leyva-Grado, V.H.; Ryder, A.B.; Miller, M.S.; Rose, J.K.; Palese, P.; Garcia-Sastre, A.; et al. Assessment of influenza virus hemagglutinin stalk-based immunity in ferrets. J. Virol. 2014, 88, 3432–3442. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.R.; Yu, Y.H.; Tseng, Y.C.; Chiang, W.L.; Chiang, M.F.; Ko, Y.A.; Chiu, Y.K.; Ma, H.H.; Wu, C.Y.; Jan, J.T.; et al. Vaccination of monoglycosylated hemagglutinin induces cross-strain protection against influenza virus infections. Proc. Natl. Acad. Sci. USA 2014, 111, 2476–2481. [Google Scholar] [PubMed]
- Leroux-Roels, I.; Borkowski, A.; Vanwolleghem, T.; Drame, M.; Clement, F.; Hons, E.; Devaster, J.M.; Leroux-Roels, G. Antigen sparing and cross-reactive immunity with an adjuvanted rH5N1 prototype pandemic influenza vaccine: A randomised controlled trial. Lancet 2007, 370, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Pearce, M.B.; Belser, J.A.; Gustin, K.M.; Pappas, C.; Houser, K.V.; Sun, X.; Maines, T.R.; Pantin-Jackwood, M.J.; Katz, J.M.; Tumpey, T.M. Seasonal trivalent inactivated influenza vaccine protects against 1918 spanish influenza virus infection in ferrets. J. Virol. 2012, 86, 7118–7125. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Baz, M.; Lu, J.; Paskel, M.; Santos, C.; Subbarao, K.; Jin, H.; Matsuoka, Y. Development of a high yield live attenuated H7N9 influenza vaccine that provides protection against homologous and heterologous h7 wild-type viruses in ferrets. J. Virol. 2014. [Google Scholar] [CrossRef]
- Chen, G.L.; Lamirande, E.W.; Cheng, X.; Torres-Velez, F.; Orandle, M.; Jin, H.; Kemble, G.; Subbarao, K. Evaluation of three live attenuated h2 pandemic influenza vaccine candidates in mice and ferrets. J. Virol. 2014, 88, 2867–2876. [Google Scholar] [CrossRef] [PubMed]
- Suguitan, A.L., Jr.; McAuliffe, J.; Mills, K.L.; Jin, H.; Duke, G.; Lu, B.; Luke, C.J.; Murphy, B.; Swayne, D.E.; Kemble, G.; et al. Live, attenuated influenza a H5N1 candidate vaccines provide broad cross-protection in mice and ferrets. PLoS Med. 2006, 3, e360. [Google Scholar] [CrossRef] [PubMed]
- Talaat, K.R.; Karron, R.A.; Luke, C.J.; Thumar, B.; McMahon, B.A.; Chen, G.L.; Lamirande, E.W.; Jin, H.; Coelingh, K.L.; Kemble, G.; et al. An open label phase i trial of a live attenuated h6n1 influenza virus vaccine in healthy adults. Vaccine 2011, 29, 3144–3148. [Google Scholar] [CrossRef] [PubMed]
- Talaat, K.R.; Karron, R.A.; Liang, P.H.; McMahon, B.A.; Luke, C.J.; Thumar, B.; Chen, G.L.; Min, J.Y.; Lamirande, E.W.; Jin, H.; et al. An open-label phase i trial of a live attenuated H2N2 influenza virus vaccine in healthy adults. Influenza Other Respir Viruses 2013, 7, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Karron, R.A.; Talaat, K.; Luke, C.; Callahan, K.; Thumar, B.; Dilorenzo, S.; McAuliffe, J.; Schappell, E.; Suguitan, A.; Mills, K.; et al. Evaluation of two live attenuated cold-adapted H5N1 influenza virus vaccines in healthy adults. Vaccine 2009, 27, 4953–4960. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Suguitan, A.; Orandle, M.; Paskel, M.; Boonnak, K.; Gardner, D.J.; Feldmann, F.; Feldmann, H.; Marino, M.; Jin, H.; et al. African green monkeys recapitulate the clinical experience with replication of live attenuated pandemic influenza virus vaccine candidates. J. Virol. 2014, 88, 8139–8152. [Google Scholar] [CrossRef] [PubMed]
- Richt, J.A.; Lekcharoensuk, P.; Lager, K.M.; Vincent, A.L.; Loiacono, C.M.; Janke, B.H.; Wu, W.H.; Yoon, K.J.; Webby, R.J.; Solorzano, A.; et al. Vaccination of pigs against swine influenza viruses by using an ns1-truncated modified live-virus vaccine. J. Virol. 2006, 80, 11009–11018. [Google Scholar] [CrossRef] [PubMed]
- Wesley, R.D.; Tang, M.; Lager, K.M. Protection of weaned pigs by vaccination with human adenovirus 5 recombinant viruses expressing the hemagglutinin and the nucleoprotein of H3N2 swine influenza virus. Vaccine 2004, 22, 3427–3434. [Google Scholar] [CrossRef] [PubMed]
- Masic, A.; Pyo, H.M.; Babiuk, S.; Zhou, Y. An eight-segment swine influenza virus harboring h1 and h3 hemagglutinins is attenuated and protective against H1N1 and H3N2 subtypes in pigs. J. Virol. 2013, 87, 10114–10125. [Google Scholar] [CrossRef] [PubMed]
- Hancock, K.; Veguilla, V.; Lu, X.; Zhong, W.; Butler, E.N.; Sun, H.; Liu, F.; Dong, L.; DeVos, J.R.; Gargiullo, P.M.; et al. Cross-reactive antibody responses to the 2009 pandemic H1N1 influenza virus. N. Engl. J. Med. 2009, 361, 1945–1952. [Google Scholar] [CrossRef] [PubMed]
- Van den Brand, J.M.; Kreijtz, J.H.; Bodewes, R.; Stittelaar, K.J.; van Amerongen, G.; Kuiken, T.; Simon, J.; Fouchier, R.A.; Del Giudice, G.; Rappuoli, R.; et al. Efficacy of vaccination with different combinations of mf59-adjuvanted and nonadjuvanted seasonal and pandemic influenza vaccines against pandemic H1N1 (2009) influenza virus infection in ferrets. J. Virol. 2011, 85, 2851–2858. [Google Scholar] [CrossRef] [PubMed]
- Ellebedy, A.H.; Ducatez, M.F.; Duan, S.; Stigger-Rosser, E.; Rubrum, A.M.; Govorkova, E.A.; Webster, R.G.; Webby, R.J. Impact of prior seasonal influenza vaccination and infection on pandemic a (H1N1) influenza virus replication in ferrets. Vaccine 2011, 29, 3335–3339. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.L.; Min, J.Y.; Lamirande, E.W.; Santos, C.; Jin, H.; Kemble, G.; Subbarao, K. Comparison of a live attenuated 2009 h1n1 vaccine with seasonal influenza vaccines against 2009 pandemic H1N1 virus infection in mice and ferrets. J. Infect. Dis. 2011, 203, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.L.; Lager, K.M.; Janke, B.H.; Gramer, M.R.; Richt, J.A. Failure of protection and enhanced pneumonia with a us H1N2 swine influenza virus in pigs vaccinated with an inactivated classical swine H1N1 vaccine. Vet. Microbiol. 2008, 126, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Reeth, K.V.; Brown, I.; Essen, S.; Pensaert, M. Genetic relationships, serological cross-reaction and cross-protection between H1N2 and other influenza a virus subtypes endemic in european pigs. Virus Res. 2004, 103, 115–124. [Google Scholar] [CrossRef]
- Macklin, M.D.; McCabe, D.; McGregor, M.W.; Neumann, V.; Meyer, T.; Callan, R.; Hinshaw, V.S.; Swain, W.F. Immunization of pigs with a particle-mediated dna vaccine to influenza a virus protects against challenge with homologous virus. J. Virol. 1998, 72, 1491–1496. [Google Scholar] [PubMed]
- Gauger, P.C.; Vincent, A.L.; Loving, C.L.; Henningson, J.N.; Lager, K.M.; Janke, B.H.; Kehrli, M.E., Jr.; Roth, J.A. Kinetics of lung lesion development and pro-inflammatory cytokine response in pigs with vaccine-associated enhanced respiratory disease induced by challenge with pandemic (2009) a/H1N1 influenza virus. Vet. Pathol. 2012, 49, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Gauger, P.C.; Vincent, A.L.; Loving, C.L.; Lager, K.M.; Janke, B.H.; Kehrli, M.E., Jr.; Roth, J.A. Enhanced pneumonia and disease in pigs vaccinated with an inactivated human-like (delta-cluster) H1N2 vaccine and challenged with pandemic 2009 H1N1 influenza virus. Vaccine 2011, 29, 2712–2719. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, C.S.; Gramer, M.R.; Barbe, F.; van Doorsselaere, J.; van Reeth, K. Efficacy of commercial swine influenza vaccines against challenge with a recent european H1N1 field isolate. Vet. Microbiol. 2010, 144, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Van Reeth, K.; Labarque, G.; de Clercq, S.; Pensaert, M. Efficacy of vaccination of pigs with different H1N1 swine influenza viruses using a recent challenge strain and different parameters of protection. Vaccine 2001, 19, 4479–4486. [Google Scholar] [CrossRef]
- Van Reeth, K.; Van Gucht, S.; Pensaert, M. Investigations of the efficacy of european H1N1- and H3N2-based swine influenza vaccines against the novel H1N2 subtype. Vet. Rec. 2003, 153, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.L.; Ma, W.; Lager, K.M.; Richt, J.A.; Janke, B.H.; Sandbulte, M.R.; Gauger, P.C.; Loving, C.L.; Webby, R.J.; Garcia-Sastre, A. Live attenuated influenza vaccine provides superior protection from heterologous infection in pigs with maternal antibodies without inducing vaccine-associated enhanced respiratory disease. J. Virol. 2012, 86, 10597–10605. [Google Scholar] [CrossRef] [PubMed]
- Braucher, D.R.; Henningson, J.N.; Loving, C.L.; Vincent, A.L.; Kim, E.; Steitz, J.; Gambotto, A.A.; Kehrli, M.E., Jr. Intranasal vaccination with replication-defective adenovirus type 5 encoding influenza virus hemagglutinin elicits protective immunity to homologous challenge and partial protection to heterologous challenge in pigs. Clin. Vaccine Immunol.: CVI 2012, 19, 1722–1729. [Google Scholar] [CrossRef] [PubMed]
- De Vleeschauwer, A.R.; Baras, B.; Kyriakis, C.S.; Jacob, V.; Planty, C.; Giannini, S.L.; Mossman, S.; van Reeth, K. Efficacy of an as03a-adjuvanted split H5N1 influenza vaccine against an antigenically distinct low pathogenic H5N1 virus in pigs. Vaccine 2012, 30, 5557–5563. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Loving, C.L.; Manischewitz, J.; King, L.R.; Gauger, P.C.; Henningson, J.; Vincent, A.L.; Golding, H. Vaccine-induced anti-ha2 antibodies promote virus fusion and enhance influenza virus respiratory disease. Sci. Transl. Med. 2013, 5, 200ra114. [Google Scholar] [CrossRef] [PubMed]
- Corti, D.; Voss, J.; Gamblin, S.J.; Codoni, G.; Macagno, A.; Jarrossay, D.; Vachieri, S.G.; Pinna, D.; Minola, A.; Vanzetta, F.; et al. A neutralizing antibody selected from plasma cells that binds to group 1 and group 2 influenza a hemagglutinins. Science 2011, 333, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Janjua, N.Z.; Skowronski, D.M.; Hottes, T.S.; Osei, W.; Adams, E.; Petric, M.; Sabaiduc, S.; Chan, T.; Mak, A.; Lem, M.; et al. Seasonal influenza vaccine and increased risk of pandemic a/h1n1-related illness: First detection of the association in british columbia, canada. Clin. Infect. Dis.: Off. Publ. Infect. Dis. Soc. Am. 2010, 51, 1017–1027. [Google Scholar] [CrossRef]
- Skowronski, D.M.; de Serres, G.; Crowcroft, N.S.; Janjua, N.Z.; Boulianne, N.; Hottes, T.S.; Rosella, L.C.; Dickinson, J.A.; Gilca, R.; Sethi, P.; et al. Association between the 2008–09 seasonal influenza vaccine and pandemic h1n1 illness during spring-summer 2009: Four observational studies from canada. PLoS Med. 2010, 7, e1000258. [Google Scholar] [CrossRef] [PubMed]
- Cowling, B.J.; Ng, S.; Ma, E.S.; Fang, V.J.; So, H.C.; Wai, W.; Cheng, C.K.; Wong, J.Y.; Chan, K.H.; Ip, D.K.; et al. Protective efficacy against pandemic influenza of seasonal influenza vaccination in children in Hong Kong: A randomized controlled trial. Clin. Infect. Dis.: Off. Publ. Infect. Dis. Soc. Am. 2012, 55, 695–702. [Google Scholar]
- Garcia-Garcia, L.; Valdespino-Gomez, J.L.; Lazcano-Ponce, E.; Jimenez-Corona, A.; Higuera-Iglesias, A.; Cruz-Hervert, P.; Cano-Arellano, B.; Garcia-Anaya, A.; Ferreira-Guerrero, E.; Baez-Saldana, R.; et al. Partial protection of seasonal trivalent inactivated vaccine against novel pandemic influenza a/H1N1 2009: Case-control study in mexico city. BMJ 2009, 339, b3928. [Google Scholar] [PubMed]
- Johns, M.C.; Eick, A.A.; Blazes, D.L.; Lee, S.E.; Perdue, C.L.; Lipnick, R.; Vest, K.G.; Russell, K.L.; DeFraites, R.F.; Sanchez, J.L. Seasonal influenza vaccine and protection against pandemic (H1N1) 2009-associated illness among us military personnel. PLoS One 2010, 5, e10722. [Google Scholar] [CrossRef] [PubMed]
- Heinen, P.P.; Rijsewijk, F.A.; de Boer-Luijtze, E.A.; Bianchi, A.T. Vaccination of pigs with a dna construct expressing an influenza virus m2-nucleoprotein fusion protein exacerbates disease after challenge with influenza a virus. J. Gen. Virol. 2002, 83, 1851–1859. [Google Scholar] [PubMed]
- Baskin, C.R.; Bielefeldt-Ohmann, H.; Garcia-Sastre, A.; Tumpey, T.M.; Van Hoeven, N.; Carter, V.S.; Thomas, M.J.; Proll, S.; Solorzano, A.; Billharz, R.; et al. Functional genomic and serological analysis of the protective immune response resulting from vaccination of macaques with an ns1-truncated influenza virus. J. Virol. 2007, 81, 11817–11827. [Google Scholar] [CrossRef] [PubMed]
- Rimmelzwaan, G.F.; Baars, M.; van Beek, R.; de Lijster, P.; de Jong, J.C.; Claas, E.C.; Osterhaus, A.D. Influenza virus subtype cross-reactivities of haemagglutination inhibiting and virus neutralising serum antibodies induced by infection or vaccination with an iscom-based vaccine. Vaccine 1999, 17, 2512–2516. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Gao, Y.; Shinya, K.; Li, C.K.; Li, Y.; Shi, J.; Jiang, Y.; Suo, Y.; Tong, T.; Zhong, G.; et al. Immunogenicity and protective efficacy of a live attenuated H5N1 vaccine in nonhuman primates. PLoS Pathog. 2009, 5, e1000409. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Margine, I.; Krammer, F. Animal Models for Influenza Viruses: Implications for Universal Vaccine Development. Pathogens 2014, 3, 845-874. https://doi.org/10.3390/pathogens3040845
Margine I, Krammer F. Animal Models for Influenza Viruses: Implications for Universal Vaccine Development. Pathogens. 2014; 3(4):845-874. https://doi.org/10.3390/pathogens3040845
Chicago/Turabian StyleMargine, Irina, and Florian Krammer. 2014. "Animal Models for Influenza Viruses: Implications for Universal Vaccine Development" Pathogens 3, no. 4: 845-874. https://doi.org/10.3390/pathogens3040845
APA StyleMargine, I., & Krammer, F. (2014). Animal Models for Influenza Viruses: Implications for Universal Vaccine Development. Pathogens, 3(4), 845-874. https://doi.org/10.3390/pathogens3040845