Exploring PFGE for Detecting Large Plasmids in Campylobacter jejuni and Campylobacter coli Isolated from Various Retail Meats


Abstract
:1. Introduction
2. Results and Discussion
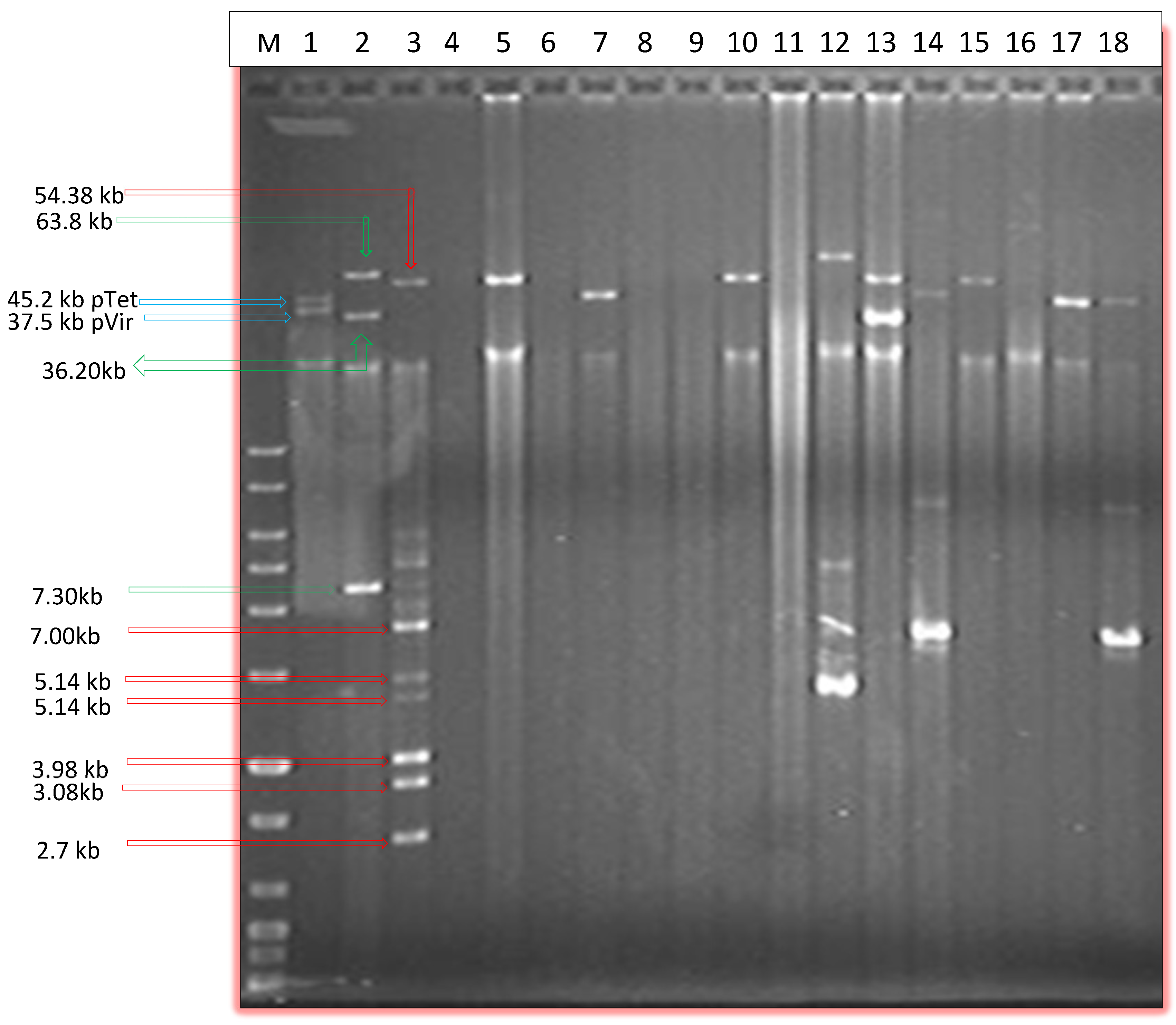
2.1. Prevalence of Plasmids by Alkaline Lysis and PFGE

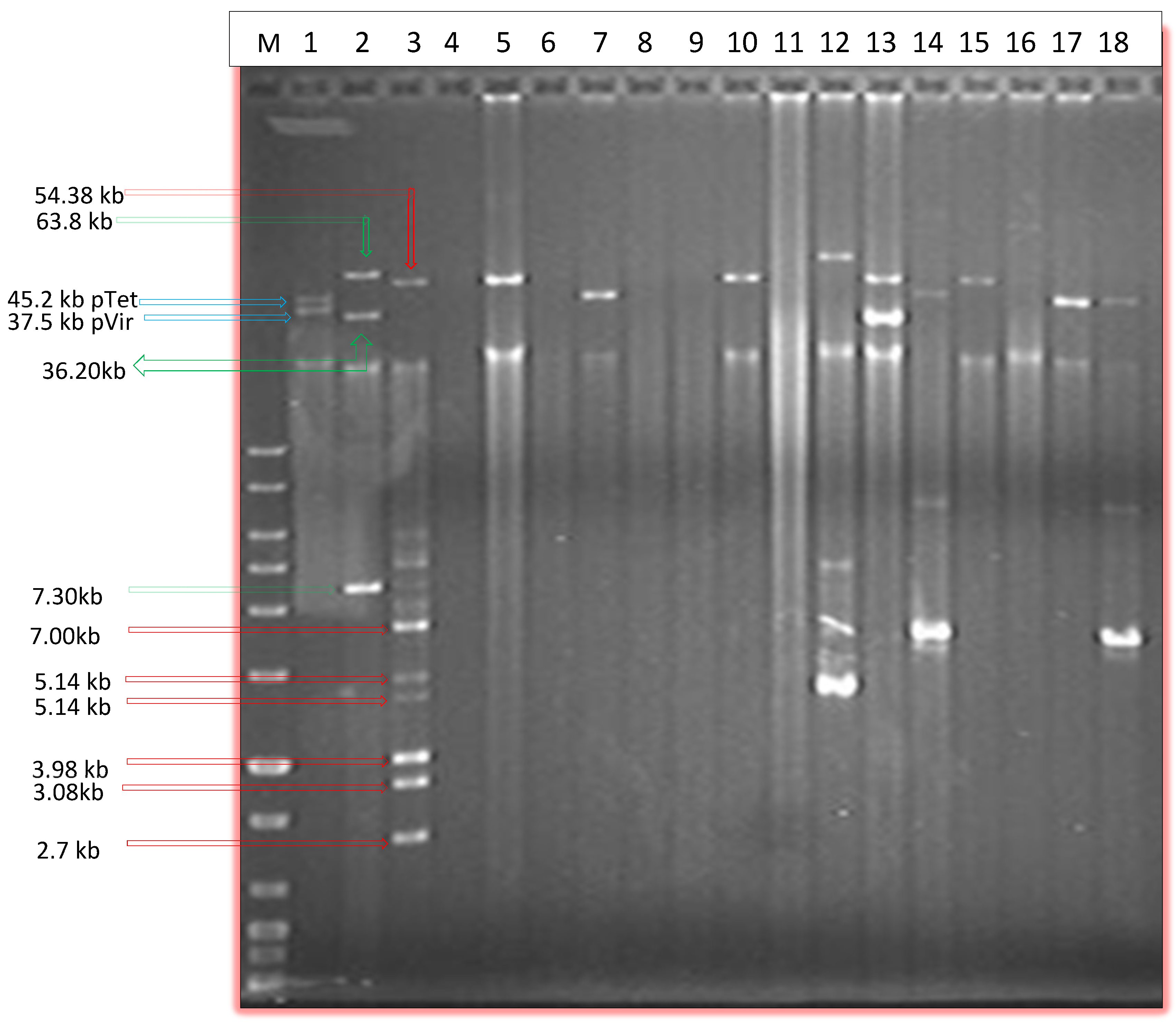{kind=link}
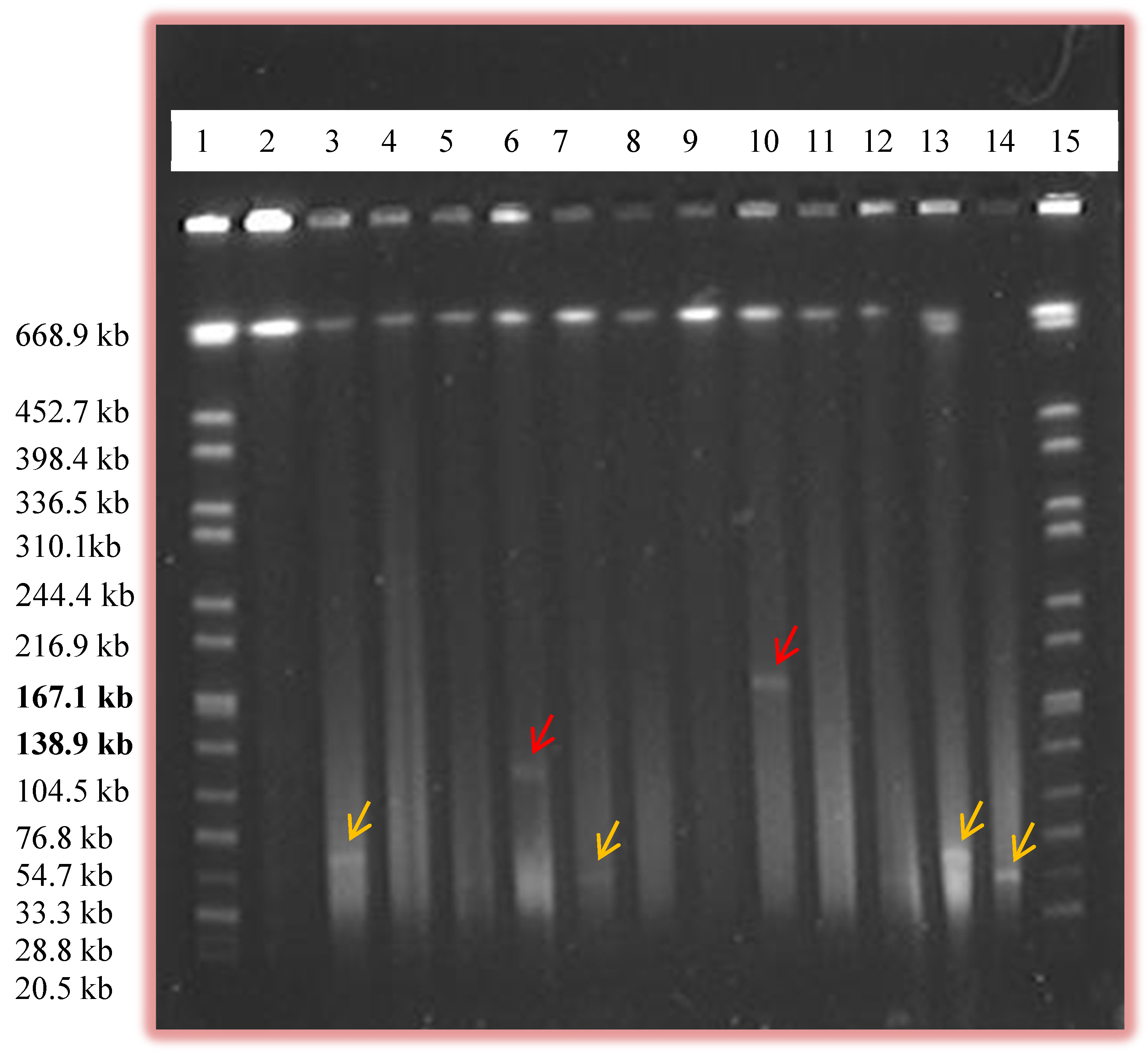{kind=link}
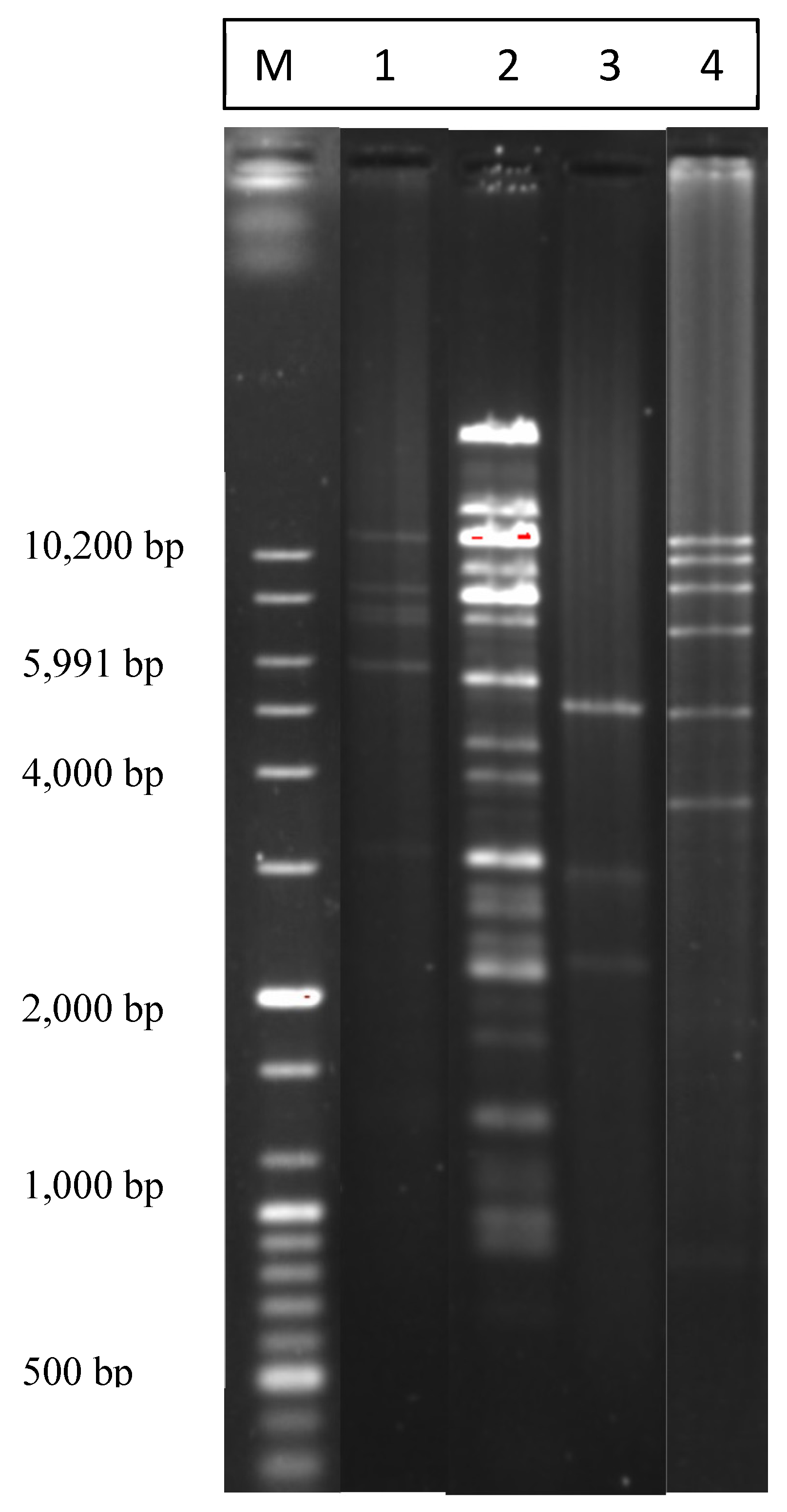{kind=link}
| Meat Sources | No of Strains in which Plasmids Were Detected n/N (%) | |||||
|---|---|---|---|---|---|---|
| Alkaline Lysis | Overall (Alkaline Lysis + PFGE) | |||||
| C. coli | C. jejuni | Total | C. coli | C. jejuni | Total | |
| Chicken | 8/9 (89%) | 11/30 (37%) | 19/39 (49%) | 9/9 (100%) | 16/30 (53%) | 25/39 (64%) |
| Chicken Livers | 35/60 (58%) | 20/38 (53%) | 55/98 (56%) | 41/60 (68%) | 23/38 (60.5%) | 64/98 (65%) |
| Chicken Gizzards | 3/3 (100%) | 3/10 (30%) | 6/13 (46%) | 3/3 (100%) | 3/10 (30%) | 6/13 (46%) |
| Beef Livers | 12/19 (63%) | 4/11 (36%) | 16/30 (53%) | 14/19 (74%) | 4/11 (36%) | 18/30 (60%) |
| Turkey | 1/2 (50%) | 3/5 (60%) | 4/7 (57%) | 1/2 (50%) | 3/5 (60%) | 4/7 (57%) |
| Pork | 2/2 (100%) | 0 | 2/2 (100%) | 2/2 (100%) | 0 | 2/2 (100%) |
| Total | 61/95 (64%) | 41/94 (44%) | 102/189 (54%) | 70/95 (73.5%) | 49/94 (52%) | 119/189 (63%) |
| Size of Plasmids (Kb) | # of Plasmids Detected by Alkaline Lysis | # of Plasmids Detected by PFGE |
|---|---|---|
| 0–45 | 89 | 20 |
| 45–90 | 56 | 81 |
| 90–135 | 3 | 10 |
| >135 | 0 | 1 |
| Total # of Plasmids | 148 | 111 |
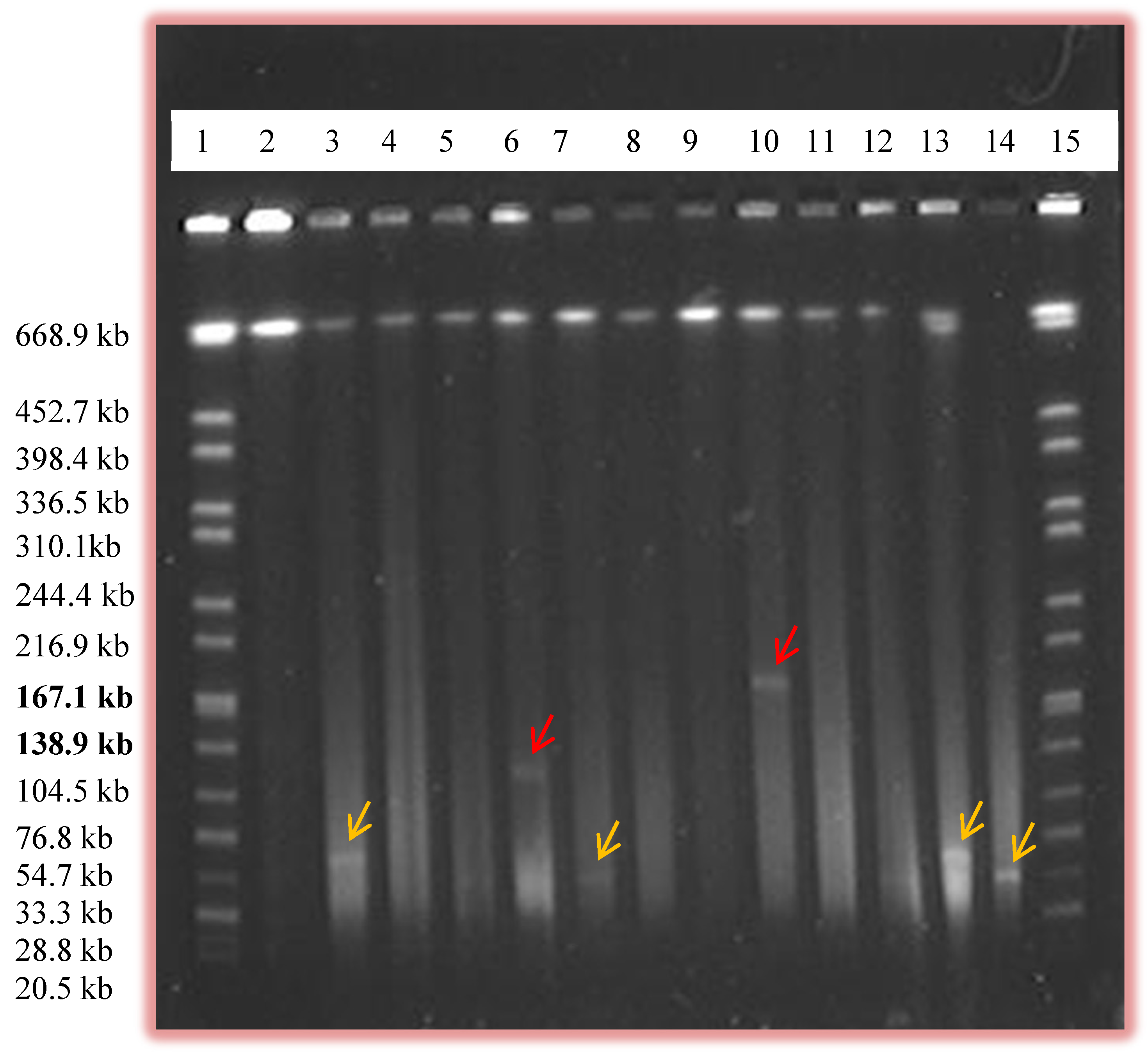
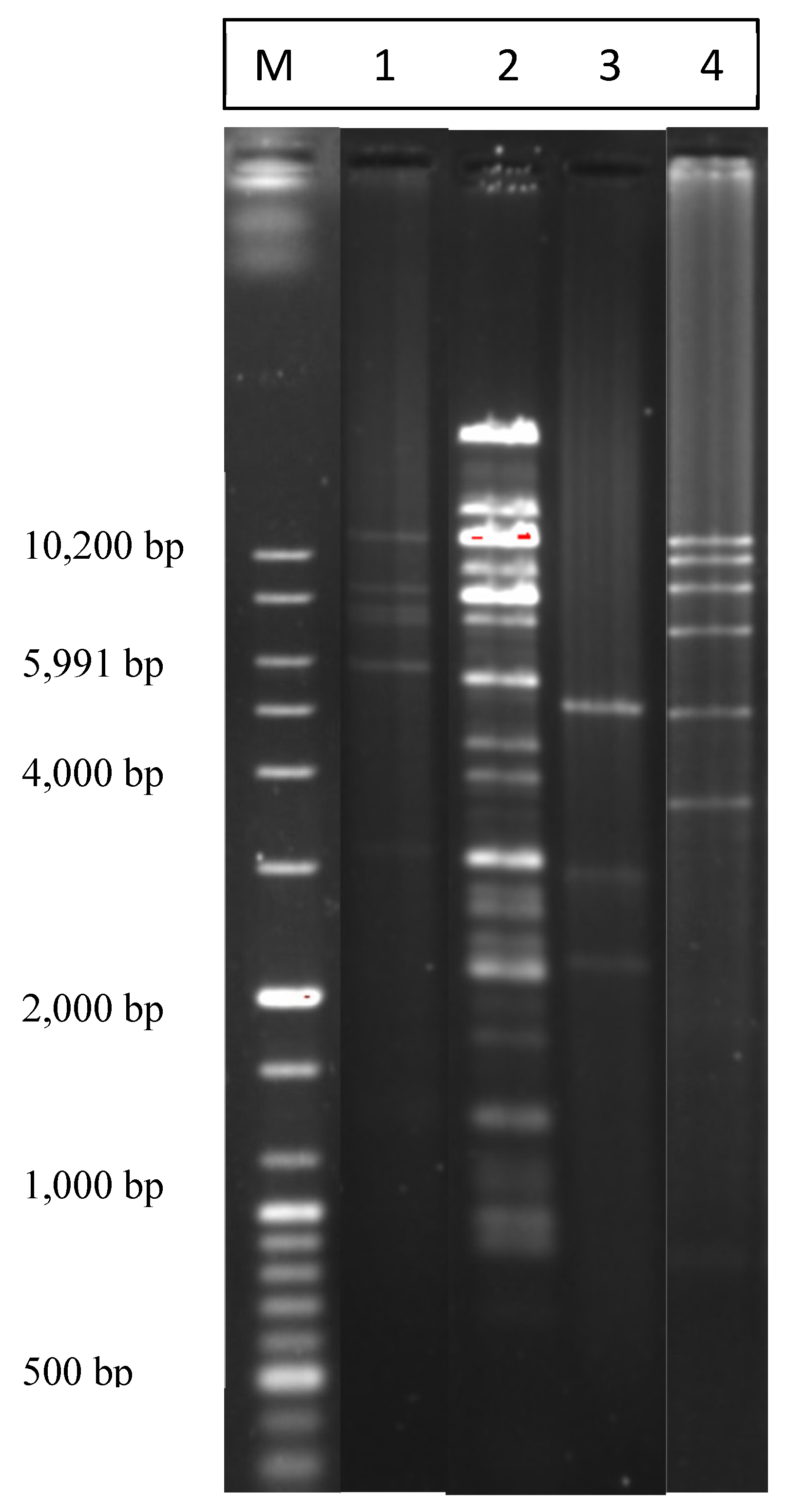
2.2. Restriction Analysis of the Isolated Plasmids
| Pattern | No. of Bands | No of Isolates | Species | Meat Source |
|---|---|---|---|---|
| A | 6 | 21 | C. coli | Chicken liver |
| B | 5 | 5 | C. coli | Beef liver |
| C | 3 | 4 + 14 | C. jejuni | Chicken + chicken liver |
| D | 5 | 3 | C. coli | Chicken liver |
| E | 4 | 2 | C. jejuni | Chicken liver |
| F | 5 | 2 + 2 | C. coli + C. jejuni | Beef liver + chicken liver |
| G | 6 | 3 | C. jejuni | Chicken liver |
| H | 6 | 2 | C. coli + C. jejuni | Beef liver |
| I | 3 | 3 | C. coli | Chicken liver |
| J | 6 | 2 | C. jejuni | Turkey |
| K | 2 | 5 | C. jejuni | Chicken |
3. Experimental Section
3.1. Bacterial Isolates
3.2. Plasmid Isolation and Screening by Alkaline Lysis and PFGE
3.3. Restriction Analysis of the Isolated Plasmids
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Allos, B.M. Campylobacter jejuni infections: Update on emerging issues and trends. Clin. Infect. Dis. 2001, 32, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control (CDC). CDC Estimates of Foodborne Illness in the United States. 2011. Available online: http://www.cdc.gov/foodborneburden/2011-foodborne-estimates.html (accessed on 16 May 2013). [Google Scholar]
- Altekruse, S.F.; Stern, N.J.; Fields, P.I.; Swerdlow, D.L. Campylobacter jejuni—An emerging foodborne pathogen. Emerg. Infect. Dis. 1999, 5, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lynn, A.M.; David, D.E.; Tang, H.; Xu, J.; Nayak, R.; Kaldhone, P.; Logue, C.M.; Foley, S.L. DNA sequence analysis of plasmids from multidrug resistant Salmonella enterica serotype Heidelberg isolates. PLoS One 2012, 7, e51160. [Google Scholar] [CrossRef]
- Johnson, T.J.; Nolan, L.K. Pathogenomics of the virulence plasmids of Escherichia coli. Microbiol. Mol. Biol. Rev. 2009, 73, 750–774. [Google Scholar] [CrossRef] [PubMed]
- Bacon, D.J.; Alm, R.A.; Burr, D.H.; Hu, L.; Kopecko, D.L.; Ewing, C.P.; Trust, T.J.; Guerry, P. Involvement of plasmid in virulence of Campylobacter jejuni 81–176. Infect. Immun. 2000, 68, 4384–4390. [Google Scholar] [CrossRef] [PubMed]
- Bacon, D.J.; Alm, R.A.; Burr, D.H.; Hu, L.; Hickey, T.E.; Ewing, C.P.; Batchelor, R.A.; Trust, T.J.; Guerry, P. DNA sequence and mutational analysis of the pVir plasmid of Campylobacter jejuni 81–176. Infect. Immun. 2002, 70, 6242–6250. [Google Scholar] [CrossRef] [PubMed]
- Tracz, D.M.; Keelan, M.; Ahmed-Bentley, J.; Gibreel, A.; Kowalewska-Grochowska, K.; Taylor, D.E. pVir and bloody diarrhea in Campylobacter jejuni Enteritis. Emerg. Infect. Dis. 2005, 11, 838–843. [Google Scholar] [PubMed]
- Schmidt-Ott, R.; Pohl, S.; Burghard, S.; Weig, M.; Grob, U. Identification and characterization of a major subgroup of conjugative Campylobacter jejuni plasmids. J. Infect. 2005, 50, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Louwen, R.P.L.; vanBelkam, A.; Wagenaar, J.A.; Doorduyn, Y.; Achterberg, R.; Endtz, H. Lack of association between the presence of the pVir plasmid and bloody diarrhea in Campylobacter jejuni Enteritis. J. Clin. Microbiol. 2006, 44, 1867–1968. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; Williams, S.; Gordon, K.P.; Nolan, C.; Plorde, J.J. Survey of plasmids and resistance factors in Campylobacter jejuni and Campylobacter coli. Antimicrob. Agents Chemother. 1985, 27, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Tai, C.L.; Lin, S.C.; Chen, Y.T. Occurrence of plasmids and tetracycline resistance among Campylobacter jejuni and Campylobacter coli isolated from whole market chickens and clinical samples. Int. J. Food Microbiol. 1994, 24, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, W.C.; Marko, M.A.; Hennessy, J.N.; Penner, J.L. Occurrence of plasmid DNA in serologically defined strains of Campylobacter jejuni and Campylobacter coli. Infect. Immun. 1983, 40, 460–463. [Google Scholar]
- Tenover, F.C.; Elvrum, P.M. Detection of two different kanamycin resistance genes in naturally occurring isolates of Campylobacter jejuni and Campylobacter coli. Antimicrob. Agents Chemother. 1988, 32, 1170–1173. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, W.C.; Munroe, D.L.G. Occurrence of plasmids and antibiotic resistance among Campylobacter jejuni and Campylobacter coli isolated from healthy and diarrheic animals. J. Clin. Microbiol. 1985, 22, 339–346. [Google Scholar]
- Ge, B.; White, D.G.; McDermott, P.F.; Girard, W.; Zhao, S.; Hubert, S.; Meng, J. Antimicrobial-resistant Campylobacter species from retail raw meats. Appl. Environ. Microbiol. 2003, 69, 3005–3007. [Google Scholar] [CrossRef] [PubMed]
- Dasti, J.I.; Gross, U.; Pohl, S.; Lugert, R.; Weig, M.; Schmidt-Ott, R. Role of plasmid –encoded tet(O) gene in tetracycline –resistant clinical isolates of Campylobacter jejuni and Campylobacter coli. J. Med. Microbiol. 2007, 56, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Batchelor, R.A.; Pearson, B.M.; Friis, L.M.; Guerry, P.; Wells, J.M. Nucleotide sequences and comparison of two large conjugative plasmids from different Campylobacters species. Microbiology 2004, 150, 3507–3517. [Google Scholar] [CrossRef] [PubMed]
- Aquino, M.H.C.; Filgueiras, A.L.L.; Ferreira, M.C.S.; Oliveira, S.S.; Bastos, M.C.; Tibana, A. Antimicrobial resistance and plasmid profiles of Campylobacter jejuni and Campylobacter coli from human and animal sources. Lett. Appl. Microbiol. 2002, 34, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Baserisalehi, M.; Bahador, N. A study on relationship of plasmids with antibiotics resistance in thermophilic Campylobacter spp. isolates from environmental samples. Biotechnology 2008, 7, 813–817. [Google Scholar] [CrossRef]
- Barton, B.M.; Harding, G.P.; Zuccarelli, A.J. A general method for detecting and sizing the plasmids. Anal. Biochem. 1995, 226, 135–240. [Google Scholar] [CrossRef]
- Noormohamed, A.; Fakhr, M.K. Incidence and Antimicrobial resistance profiling of Campylobacter in retail chicken livers and gizzards. Foodborne Pathog. Dis. 2012, 9, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Noormohamed, A.; Fakhr, M.K. A higher prevalence rate of Campylobacter in retail beef livers compared to other beef and pork meat cuts. Int. J. Environ. Res. Public Health 2013, 10, 2058–2068. [Google Scholar] [CrossRef] [PubMed]
- Sagara, H.; Mochizuki, A.; Okamura, N.; Nakaya, R. Antimicrobial resistance of Campylobacter jejuni and Campylobacter coli with special reference to plasmid profiles of Japanese clinical isolates. Antimicrob. Agents Chemother. 1987, 31, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Pearson, B.M.; Rokney, A.; Crossman, L.C.; Miller, W.G.; Wain, J.; van Vliet, A.H.M. Complete genome sequence of the Campylobacter coli Clinical isolate. Genome Anounc. 2013. [Google Scholar] [CrossRef]
- Chen, Y.; Mukherjee, S.; Hoffman, M.; Kotewicz, M.L.; Young, S.; Abbott, J.; Luo, Y.; Davidson, M.K.; Allard, M.; McDermott, P.; et al. Whole-genome sequencing of gentamicin-resistant Campylobacter coli isolated from U.S. retail meats reveals novel plasmid-mediated aminoglycoside resistance genes. Antimicrob. Agents Chemother. 2013, 57, 5398–5405. [Google Scholar]
- Centers for Disease Control (CDC). Standard Operating Procedure for PulseNet PFGE of Campylobacter jejuni. 2011. Available online: http://www.cdc.gov/pulsenet/PDF/campylobacter-pfge-protocol-508c.pdf (accessed on 15 August 2011). [Google Scholar]
- Noormohamed, A.; Fakhr, M.K. Molecular Typing of Campylobacter jejuni and Campylobacter coli isolated from various retail meats by MLST and PFGE. Foods 2014, 3, 82–93. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marasini, D.; Fakhr, M.K. Exploring PFGE for Detecting Large Plasmids in Campylobacter jejuni and Campylobacter coli Isolated from Various Retail Meats. Pathogens 2014, 3, 833-844. https://doi.org/10.3390/pathogens3040833
Marasini D, Fakhr MK. Exploring PFGE for Detecting Large Plasmids in Campylobacter jejuni and Campylobacter coli Isolated from Various Retail Meats. Pathogens. 2014; 3(4):833-844. https://doi.org/10.3390/pathogens3040833
Chicago/Turabian StyleMarasini, Daya, and Mohamed K. Fakhr. 2014. "Exploring PFGE for Detecting Large Plasmids in Campylobacter jejuni and Campylobacter coli Isolated from Various Retail Meats" Pathogens 3, no. 4: 833-844. https://doi.org/10.3390/pathogens3040833
APA StyleMarasini, D., & Fakhr, M. K. (2014). Exploring PFGE for Detecting Large Plasmids in Campylobacter jejuni and Campylobacter coli Isolated from Various Retail Meats. Pathogens, 3(4), 833-844. https://doi.org/10.3390/pathogens3040833