Exploring Dangerous Connections between Klebsiella pneumoniae Biofilms and Healthcare-Associated Infections


Abstract
:1. Introduction
2. Results and Discussion
2.1. Bacteria Characterization

{kind=link}
{kind=link}
| Strain | Source | Year | Serologic Group | Fimbriae | β-lactamases | |
|---|---|---|---|---|---|---|
| Kp45 | Neck swab | 1980 | K:2 | fimH | mrkD | TEM-1 |
| Kp26 | Rectal swab | K:2 | n.a. | n.a. | TEM-1 | |
| Kp703 | Urine | O:1 | n.a. | mrkD | TEM-1 | |
| Kp3921 | 2010 | n.a. | fimH | mrkD | CTX-M-15 | |
| Kp2948 | Wound | K:2 | fimH | mrkD | KPC-3; TEM-1 | |
| Kp3421 | Urine | 2011 | n.a. | fimH | mrkD | CTX-M-15 |
| Kp3407 | n.a. | fimH | mrkD | KPC-3 | ||
| Kp3466 | n.a. | n.a. | n.a. | TEM-163 | ||
| Kp3385 | n.a. | fimH | mrkD | KPC-3 | ||
2.2. Biofilm Assembly
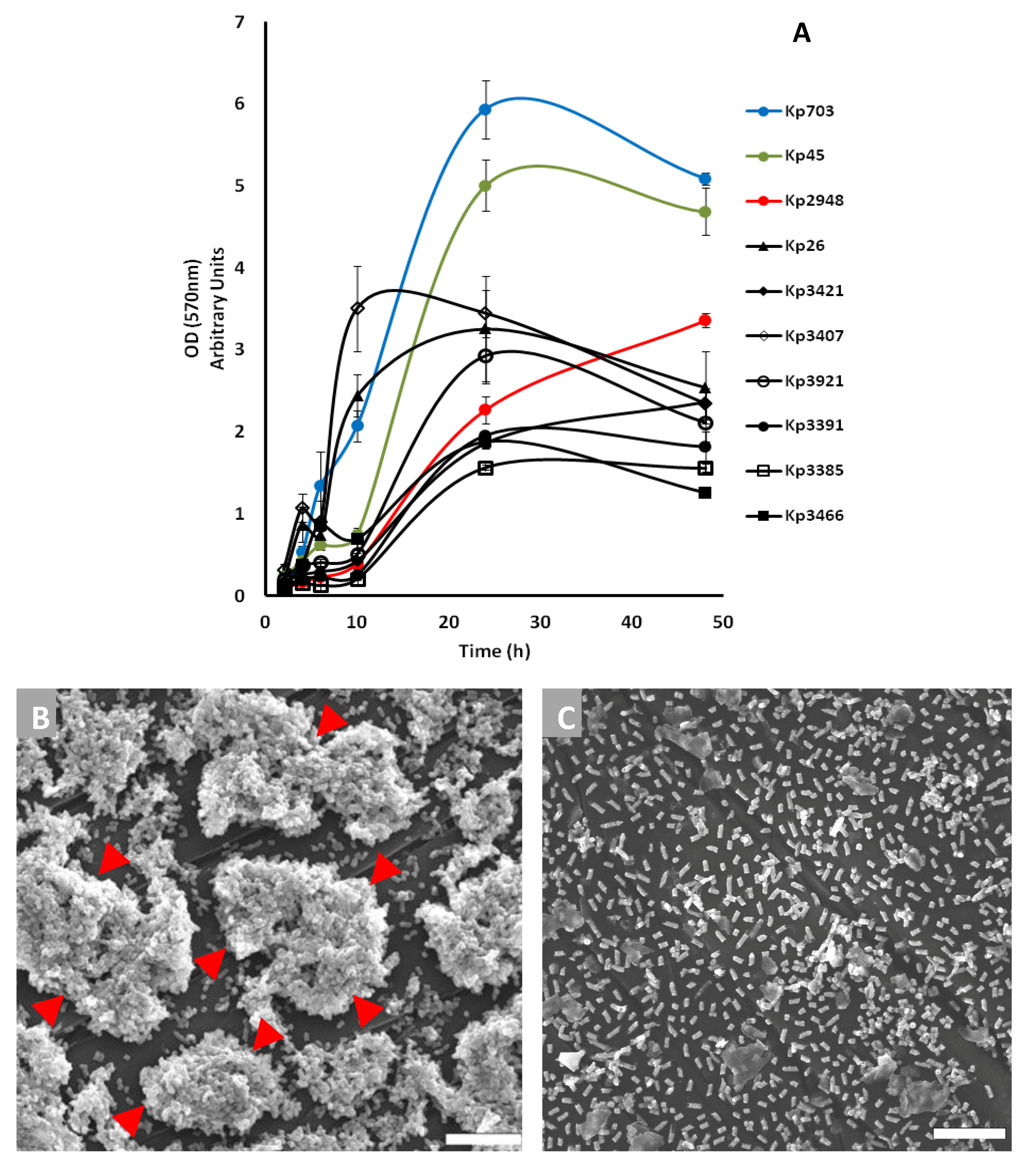
2.3. Scanning Electron Microscopy
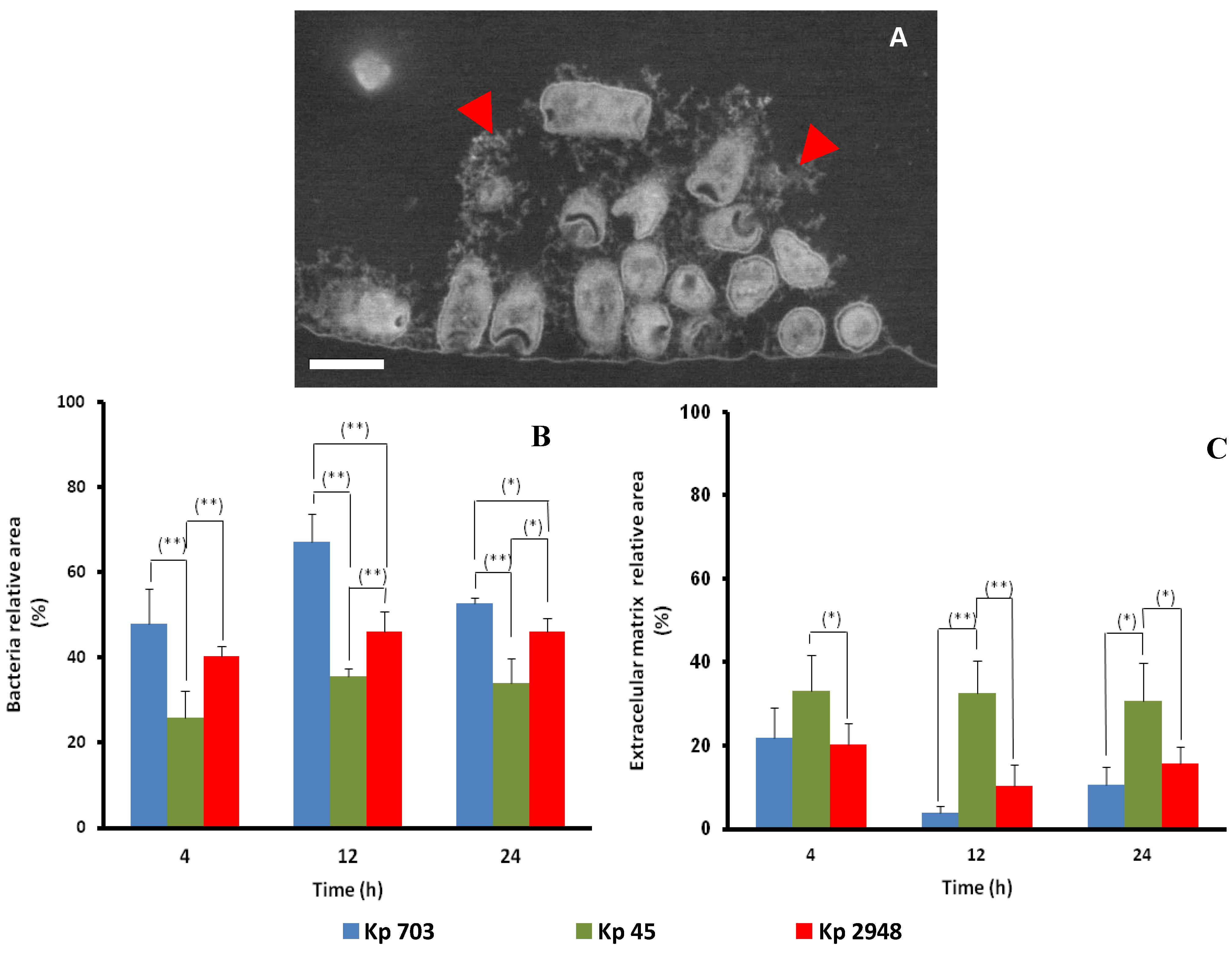
2.4. Antimicrobial Activity of Planktonic and Biofilm-Embedded K. pneumoniae Strains
| A: MIC (Planktonic) | ||||
| Drug | Amoxicillin (μg/mL) | Fosfomycin (μg/mL) | Gentamicin (μg/mL) | |
| Strain | ||||
| Kp45 | 250 | 0.781 | 3.05 | |
| Kp703 | 250 | <0.488 | 0.760 | |
| Kp2948 | >500 | 0.781 | 1.52 | |
| B: MIC (Biofilm) | ||||
| Drug | Amoxicillin (μg/mL) | Fosfomycin (μg/mL) | Gentamicin (μg/mL) | |
| Strain | ||||
| Kp45 | >2,500 | 0.781 | 24.4 | |
| Kp703 | >2,500 | 500 | 195 | |
| Kp2948 | 2,500 | 0.781 | 3.05 | |
3. Experimental Section
3.1. Bacterial Strains
3.2. Capsular Type
3.3. Biofilm Assay
3.4. Scanning Electron Microscopy (SEM)
3.5. Antibiotic Susceptibility Test
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- European Centre for Disease Prevention and Control. Annual Epidemiological Report 2012, Reporting on 2010 Surveillance Data and 2011 Epidemic Intelligence Data; ECDC: Stockholm, Sweden, 2013. [Google Scholar]
- European Centre for Disease Prevention and Control. Surveillance of Surgical Site Infections in European Hospitals–HAISSI Protocol; Protocol Version 1.02; ECDC: Stockholm, Sweden, 2012. [Google Scholar]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. J. 2002, 8, 881–890. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Hooper, D.C. Hospital-Acquired Infections Due to Gram-Negative Bacteria. N. Engl. J. Med. 2010, 362, 1804–1813. [Google Scholar] [CrossRef]
- Bales, P.M.; Renke, E.M.; May, S.L.; Shen, Y.; Nelson, D.C. Purification and characterization of biofilm-associated EPS Exopolysaccharides from ESKAPE organisms and other pathogens. PLoS One 2013, 8, e67950. [Google Scholar]
- Pomakov, A.O.; Hsiao, C.-B.; Beanan, J.M.; Olson, R.; MacDonald, U.; Keynan, Y.; Russo, T.A. Clinical and phenotypic differences between classic and hypervirulent Klebsiella pneumonia: An emerging and under-recognized pathogenic variant. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 981–989. [Google Scholar] [CrossRef]
- Chen, P.; Seth, A.K.; Abercrombie, J.J.; Mustoe, T.A.; Leung, K.P. Activity of Imipenem against Klebsiella pneumoniae Biofilms In Vitro and In Vivo. Antimicrob. Agents Chemother. 2014, 58, 1208–1213. [Google Scholar] [CrossRef]
- Schroll, C.; Barken, K.B.; Karen, A.K.; Struve, C. Role of type1 and type 3 fimbriae in Klebsiella pneumoniae biofilm formation. BMC Microbiol. 2010, 10, 179–188. [Google Scholar] [CrossRef]
- Stahlhut, S.G.; Chattopadhyay, S.; Kisiela, D.I.; Hvidtfeldt, K.; Clegg, S.; Struve, C.; Sokurenko, E.V.; Krogfelta, K.A. Structural and Population Characterization of MrkD, the Adhesive Subunit of Type 3 Fimbriae. J. Bacteriol. 2013, 195, 5602–5613. [Google Scholar] [CrossRef]
- Watnick, P.; Kolter, R. Biofilm, city of microbes. J. Bacteriol. 2000, 182, 2675–2679. [Google Scholar] [CrossRef]
- Reichelt, M.; Joubert, L.; Perrino, J.; Koh, A.L.; Phanwar, I.; Arvin, A.M. 3D reconstruction of VZV infected cell nuclei and PML nuclear cages by serial section array scanning electron microscopy and electron tomography. PLoS Pathog. 2012, 8, e1002740. [Google Scholar] [CrossRef]
- Jefferson, K.K. What drives bacteria to produce a biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef]
- Lewis, K. Riddle of biofilm resistance. Antimicrob. Agents Chemother. 2001, 45, 999–1007. [Google Scholar] [CrossRef]
- Antonia, C.K.; Petros, I.R.; Vouloumanou, E.K.; Gkegkes, I.D.; Falagas, M.E. Synergy of fosfomycin with other antibiotics for Gram-positive and Gram-negative bacteria. Eur. J. Clin. Pharmacol. 2010, 66, 359–368. [Google Scholar] [CrossRef]
- Caneiras, C.; Calisto, F.; Narciso, A.; Arriaga, S.; Melo-Cristino, J.; Duarte, A. Resistance and Virulence Characterisation of CTX-M15 and KPC-3 Klebsiella pneumoniae Clinical Isolates. In Proceedings of the 21st European Congress of Clinical Microbiology and Infectious Diseases, Milan, Italy, 7–10 May 2011.
- Endimiani, A.; Patel, G.; Hujer, K.M.; Swaminathan, M.; Perez, F.; Rice, L.B.; Jacobs, M.R.; Bonomo, R.A. In vitro activity of fosfomycin against blaKPC-containing Klebsiella pneumoniae isolates, including those nonsusceptible to tigecycline and/or colistin. Antimicrob. Agents Chemother. 2010, 54, 526–529. [Google Scholar] [CrossRef]
- De Araujo, C.; Balestrino, D.; Roth, L.; Charbonnel, N.; Forestier, C. Quorum sensing affects biofilm formation through lipopolysaccharide synthesis in Klebsiella pneumoniae. Res. Microbiol. 2010, 161, 595–603. [Google Scholar] [CrossRef]
- Wu, K.M.; Li, L.H.; Yan, J.J.; Tsao, N.; Liao, T.L.; Tsai, H.C.; Fung, C.P.; Chen, H.J.; Liu, Y.M.; Wang, J.T.; et al. Genome sequencing and comparative analysis of Klebsiella pneumoniae NTUH-K2044, a strain causing liver abscess and meningitis. J. Bacteriol. 2009, 191, 4492–4501. [Google Scholar] [CrossRef]
- Arakawa, Y.; Wacharotayankun, R.; Nagatsuka, T.; Ito, H.; Kato, N.; Ohta, M. Genomic Organization of the Klebsiella pneumoniae cps Region Responsible for Serotype K2 Capsular Polysaccharide Synthesis in the Virulent Strain Chedid. J. Bacteriol. 1995, 177, 1788–1796. [Google Scholar]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Etienne, G.; Laval, F.; Villeneuve, C.; Dinadayala, P.; Abouwarda, A.; Zerbib, D.; Galamba, A.; Daffé, M. The cell envelope structure and properties of Mycobacterium smegmatis mc2 155: Is there a clue for the unique transformability of the strain? Microbiology 2005, 151, 2075–2086. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bandeira, M.; Carvalho, P.A.; Duarte, A.; Jordao, L. Exploring Dangerous Connections between Klebsiella pneumoniae Biofilms and Healthcare-Associated Infections. Pathogens 2014, 3, 720-731. https://doi.org/10.3390/pathogens3030720
Bandeira M, Carvalho PA, Duarte A, Jordao L. Exploring Dangerous Connections between Klebsiella pneumoniae Biofilms and Healthcare-Associated Infections. Pathogens. 2014; 3(3):720-731. https://doi.org/10.3390/pathogens3030720
Chicago/Turabian StyleBandeira, Maria, Patricia Almeida Carvalho, Aida Duarte, and Luisa Jordao. 2014. "Exploring Dangerous Connections between Klebsiella pneumoniae Biofilms and Healthcare-Associated Infections" Pathogens 3, no. 3: 720-731. https://doi.org/10.3390/pathogens3030720
APA StyleBandeira, M., Carvalho, P. A., Duarte, A., & Jordao, L. (2014). Exploring Dangerous Connections between Klebsiella pneumoniae Biofilms and Healthcare-Associated Infections. Pathogens, 3(3), 720-731. https://doi.org/10.3390/pathogens3030720