
Species Diversity and Molecular Screening of Pyrethroid Resistance Mediated by the Voltage-Gated Sodium Channel in Ixodid Ticks from Puducherry, India


 , , , and
, , , and 
Abstract
1. Introduction
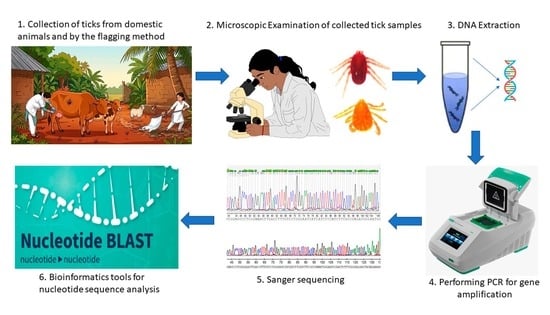
2. Materials and Methods
2.1. Study Area and Sample Collection
2.2. Morphological Identification
2.3. Calculation of Tick Infestation Indices
2.4. Genomic DNA Extraction
2.5. PCR Amplification of VGSC Gene
2.6. Sequencing and Data Analysis
3. Results
3.1. Tick Collection and Host Infestation Rate
3.2. Species Diversity and Proportional Representation
3.3. Mean Abundance and Mean Infestation Intensity
3.4. Acaricide Usage Patterns
3.5. Molecular Screening for VGSC Mutations
4. Discussion
4.1. Tick Species Diversity and Host Associations
4.2. Absence of Detectable kdr Mutations in Tick Populations from Puducherry
4.3. Indian and Global Perspectives on Acaricide Resistance
4.4. Ecological and Management Implications
4.5. Limitations and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PCR | Polymerase Chain Reaction |
| VGSC | Voltage-gated Sodium Channel |
| kdr | Knock Down Resistance |
| CI | Confidence Interval |
| SNP | Single-nucleotide polymorphisms |
| NCBI | National Center for Biotechnology Information |
| IVRI | Indian Veterinary Research Institute |
| LPT | larval packet test |
| LTT | larval tarsal test |
| SPs | synthetic pyrethroids |
References
- Raju, H.K.; Elango, A.; Krishnamoorthi, R.; Rahi, M. Prevalence and Abundance of Ixodid Ticks in Domestic Mammals in Villages at the Forest Fringes of the Western Ghats, India. Animals 2025, 15, 2005. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Azhahianambi, P.; de la Fuente, J. Control of Ticks of Ruminants, with Special Emphasis on Livestock Farming Systems in India: Present and Future Possibilities for Integrated Control—A Review. Exp. Appl. Acarol. 2006, 40, 49–66. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Nagar, G. Problem of Ticks and Tick-Borne Diseases in India with Special Emphasis on Progress in Tick Control Research: A Review. J. Vector Borne Dis. 2014, 51, 259. [Google Scholar] [CrossRef]
- Jongejan, F.; Uilenberg, G. The Global Importance of Ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Babu, N.N.; Kadni, T.S.; Rao, S.R.; Arunkumar, V.S.; Nepali, I.; Saha, A.; Varamballi, P.; Mudgal, P.P. Challenges in Combating Tick-Borne Disease Transmissions in India: Emphasizing the Importance of a One-Health Approach. One Health 2025, 21, 101199. [Google Scholar] [CrossRef]
- de Castro, J.J. Sustainable Tick and Tickborne Disease Control in Livestock Improvement in Developing Countries. Vet. Parasitol. 1997, 71, 77–97. [Google Scholar] [CrossRef]
- Minjauw, B.; Mcleod, B.A. Tick-Borne Diseases and Poverty: The Impact of Ticks and Tick-Borne Diseases on the Livelihood of Small-Scale and Marginal Livestock Owners in India and Eastern and Southern Africa; Research Report; DFID Animal Health Programme: Edinburgh, UK, 2003. [Google Scholar]
- Balakrishnan, N. Emerging and Re-Emerging Tickborne Diseases of Public Health Importance in India. J. Commun. Dis. 2020, 52, 27–32. [Google Scholar]
- Soderlund, D.M.; Knipple, D.C. The Molecular Biology of Knockdown Resistance to Pyrethroid Insecticides. Insect Biochem. Mol. Biol. 2003, 33, 563–577. [Google Scholar] [CrossRef]
- Abbas, R.Z.; Zaman, M.A.; Colwell, D.D.; Gilleard, J.; Iqbal, Z. Acaricide Resistance in Cattle Ticks and Approaches to Its Management: The State of Play. Vet. Parasitol. 2014, 203, 6–20. [Google Scholar] [CrossRef]
- Guerrero, F.D.; Lovis, L.; Martins, J.R. Acaricide Resistance Mechanisms in Rhipicephalus (Boophilus) microplus. Rev. Bras. Parasitol. Vet. 2012, 21, 1–6. [Google Scholar] [CrossRef]
- Ghosh, S.; Gupta, S.; Ajith Kumar, K.G.; Sharma, A.K.; Kumar, S.; Nagar, G.; Kumar, R.; Paul, S.; Fular, A.; Chigure, G.; et al. Characterization and Establishment of a Reference Deltamethrin and Cypermethrin Resistant Tick Line (IVRI-IV) of Rhipicephalus (Boophilus) microplus. Pestic. Biochem. Physiol. 2017, 138, 66–70. [Google Scholar] [CrossRef]
- Rosario-Cruz, R.; Guerrero, F.D.; Miller, R.J.; Rodriguez-Vivas, R.I.; Tijerina, M.; Dominguez-Garcia, D.I.; Hernandez-Ortiz, R.; Cornel, A.J.; McAbee, R.D.; Alonso-Diaz, M.A. Molecular Survey of Pyrethroid Resistance Mechanisms in Mexican Field Populations of Rhipicephalus (Boophilus) microplus. Parasitol. Res. 2009, 105, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Klafke, G.M.; Miller, R.J.; Tidwell, J.P.; Thomas, D.B.; Sanchez, D.; Feria Arroyo, T.P.; Pérez de León, A.A. High-Resolution Melt (HRM) Analysis for Detection of SNPs Associated with Pyrethroid Resistance in the Southern Cattle Fever Tick, Rhipicephalus (Boophilus) Microplus (Acari: Ixodidae). Int. J. Parasitol. Drugs Drug Resist. 2019, 9, 100–111. [Google Scholar] [CrossRef]
- Stone, B.F.; Webber, L.G. Cattle Ticks, Boophilus microplus, Resistant to DDT, BHC, and Dieldrin. Aust. J. Agric. Res. 1960, 11, 106–119. [Google Scholar] [CrossRef]
- Li, A.Y.; Davey, R.B.; Miller, R.J.; George, J.E. Resistance to Coumaphos and Diazinon in Boophilus microplus (Acari: Ixodidae) and Evidence for the Involvement of an Oxidative Detoxification Mechanism. J. Med. Entomol. 2003, 40, 482–490. [Google Scholar] [CrossRef]
- Miller, R.J.; Davey, R.B.; George, J.E. Modification of the Food and Agriculture Organization Larval Packet Test to Measure Amitraz-Susceptibility against Ixodidae. J. Med. Entomol. 2002, 39, 645–651. [Google Scholar] [CrossRef]
- Perez-Cogollo, L.C.; Rodriguez-Vivas, R.I.; Ramirez-Cruz, G.T.; Miller, R.J. First Report of the Cattle Tick Rhipicephalus microplus Resistant to Ivermectin in Mexico. Vet. Parasitol. 2010, 168, 165–169. [Google Scholar] [CrossRef]
- Kumar, R.; Nagar, G.; Sharma, A.K.; Kumar, S.; Ray, D.D.; Chaudhuri, P.; Ghosh, S. Survey of Pyrethroids Resistance in Indian Isolates of Rhipicephalus (Boophilus) microplus: Identification of C190A Mutation in the Domain II of the Para-Sodium Channel Gene. Acta Trop. 2013, 125, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.K.; Gelot, I.S.; Jyoti; Bhat, S.A.; Singh, H.; Singh, V. Detection of Acaricidal Resistance in Hyalomma anatolicum anatolicum from Banaskantha District, Gujarat. J. Parasit. Dis. 2013, 39, 563. [Google Scholar] [CrossRef]
- Reck, J.; Klafke, G.M.; Webster, A.; Dall’Agnol, B.; Scheffer, R.; Souza, U.A.; Corassini, V.B.; Vargas, R.; dos Santos, J.S.; Martins, J.R.d.S. First Report of Fluazuron Resistance in Rhipicephalus microplus: A Field Tick Population Resistant to Six Classes of Acaricides. Vet. Parasitol. 2014, 201, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Siriporn, B.; Juasook, A.; Neelapaijit, N.; Kaewta, P.; Wu, Z. Detection of Ivermectin and Fipronil Resistance in Rhipicephalus sanguineus Sensu Lato in Maha Sarakham, Thailand. Vet. World 2023, 16, 1661. [Google Scholar] [CrossRef] [PubMed]
- Kleinschmidt, H.; Stone, N.E.; Ballard, R.; Thornton, N.B.; Pemberton, G.L.; Becker, S.; Klafke, G.M.; Wagner, D.M.; Busch, J.D. First Report of VGSC Mutations for Resistance to Synthetic Pyrethroids in Brown Dog Ticks (Rhipicephalus sanguineus Sensu Stricto) from Brazil. Parasites Vectors 2025, 18, 474. [Google Scholar] [CrossRef]
- He, H.; Chen, A.C.; Davey, R.B.; Ivie, G.W.; George, J.E. Identification of a Point Mutation in the Para-Type Sodium Channel Gene from a Pyrethroid-Resistant Cattle Tick. Biochem. Biophys. Res. Commun. 1999, 261, 558–561. [Google Scholar] [CrossRef]
- Morgan, J.A.T.; Corley, S.W.; Jackson, L.A.; Lew-Tabor, A.E.; Moolhuijzen, P.M.; Jonsson, N.N. Identification of a Mutation in the Para-Sodium Channel Gene of the Cattle Tick Rhipicephalus (Boophilus) microplus Associated with Resistance to Synthetic Pyrethroid Acaricides. Int. J. Parasitol. 2009, 39, 775–779. [Google Scholar] [CrossRef]
- Jonsson, N.N.; Cutullè, C.; Corley, S.W.; Seddon, J.M. Identification of a Mutation in the Para-Sodium Channel Gene of the Cattle Tick Rhipicephalus microplus Associated with Resistance to Flumethrin but Not to Cypermethrin. Int. J. Parasitol. 2010, 40, 1659–1664. [Google Scholar] [CrossRef]
- Kumar, S.; Sharma, A.K.; Nagar, G.; Rawat, S.S.; Tiwari, S.S.; Kumar, R.; Dhakad, M.L.; Sharma, R.K.; Saxana, R.K.; Mehraniya, R.S.; et al. Characterization of Acaricide Resistance in Tick Isolates Collected from Rajasthan, India. Indian J. Anim. Sci. 2016, 86, 14–23. [Google Scholar] [CrossRef]
- Sharif, M. A Revision of the Indian Ixodidae with Special Reference to the Collection in the Indian Museum. Rec. Zool. Surv. India 1928, 30, 217–344. [Google Scholar] [CrossRef]
- Geevarghese, G. Haemaphysalis Ticks of India, 1st ed.; Elsevier Insights; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Trapido, H.; Varma, M.G.R.; Rajagopalan, P.K.; Singh, K.R.P.; Rebello, M.J.; Centre, V.R. A Guide to the Identification of All Stages of the Haemaphysalis Ticks of South India. Bull. Entomol. Res. 1964, 55, 249–270. [Google Scholar] [CrossRef]
- Stone, N.E.; Olafson, P.U.; Davey, R.B.; Buckmeier, G.; Bodine, D.; Sidak-Loftis, L.C.; Giles, J.R.; Duhaime, R.; Miller, R.J.; Mosqueda, J.; et al. Multiple Mutations in the Para-Sodium Channel Gene Are Associated with Pyrethroid Resistance in Rhipicephalus microplus from the United States and Mexico. Parasites Vectors 2014, 7, 456. [Google Scholar] [CrossRef] [PubMed]
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying Parasites in Samples of Hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on Its Own Terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Reiczigel, J.; Marozzi, M.; Fábián, I.; Rózsa, L. Biostatistics for Parasitologists—A Primer to Quantitative Parasitology. Trends Parasitol. 2019, 35, 277–281. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2025. [Google Scholar]
- Nataraj, N.; Muthuraman, K.; Ayyanar, E.; Ashokkumar, M.; Kumar, A.; Srinivasan, L.; Devaraju, P. Ectoparasite Diversity in Pets and Livestock from Puducherry, India. Int. J. Acarol. 2021, 47, 628–632. [Google Scholar] [CrossRef]
- Latha, B.R.; Aiyasami, S.S.; Pattabiraman, G.; Sivaraman, T.; Rajavelu, G. Seasonal Activity of Ticks on Small Ruminants in Tamil Nadu State, India. Trop. Anim. Health Prod. 2004, 36, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Elango, A.; Shriram, A.N.; Raju, H.K.; Padmaja, S.; Kumar, A. Spatial and Temporal Prevalence, Abundance and Infestation Intensity of the Ixodid Tick Population on Small Domestic Ruminants (Goat and Sheep) in Different Agro-Climatic Regions of Tamil Nadu. Vet. Parasitol. Reg. Stud. Rep. 2025, 59, 101235. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F. Biology and Ecology of the Brown Dog Tick, Rhipicephalus sanguineus. Parasites Vectors 2010, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Lovis, L.; Mendes, M.C.; Perret, J.-L.; Martins, J.R.; Bouvier, J.; Betschart, B.; Sager, H. Use of the Larval Tarsal Test to Determine Acaricide Resistance in Rhipicephalus (Boophilus) microplus Brazilian Field Populations. Vet. Parasitol. 2013, 191, 323–331. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host | No. of Host Examined | No. of Host Infested | Infestation Rate (%) | Lower 95% CI | Upper 95% CI |
|---|---|---|---|---|---|
| Cattle | 413 | 169 | 40.92 | 36.28 | 45.72 |
| Dog | 15 | 9 | 60.00 | 35.75 | 80.18 |
| Goat | 124 | 24 | 19.35 | 13.37 | 27.19 |
| Total | 552 | 202 | 36.59 | 32.68 | 40.69 |
| Species | Cattle | Goat | Dog | Total | ||||
|---|---|---|---|---|---|---|---|---|
| No. of Ticks | 0% | No. of Ticks | % | No. of Ticks | % | No. of Ticks | % | |
| Haemaphysalis bispinosa | 1505 | 43.30 | 39 | 15.12 | 0 | 0.00 | 1544 | 40.86 |
| Haemaphysalis intermedia | 424 | 12.20 | 208 | 80.62 | 6 | 13.33 | 638 | 16.88 |
| Hyalomma kumari | 4 | 0.12 | 0 | 0.00 | 0 | 0.00 | 4 | 0.11 |
| Rhipicephalus (Bo.) annulatus | 1467 | 42.20 | 0 | 0.00 | 0 | 0.00 | 1467 | 38.82 |
| Rhipicephalus (Bo.) microplus | 13 | 0.37 | 0 | 0.00 | 0 | 0.00 | 13 | 0.34 |
| Rhipicephalus sanguineus | 15 | 0.43 | 3 | 1.16 | 25 | 55.56 | 43 | 1.14 |
| Rhipicephalus simus | 8 | 0.23 | 4 | 1.55 | 14 | 31.11 | 26 | 0.69 |
| Rhipicephalus haemaphysaloides | 40 | 1.15 | 4 | 1.55 | 0 | 0.00 | 44 | 1.16 |
| Grand Total | 3476 | 91.98 | 258 | 6.83 | 45 | 1.19 | 3779 | 100.00 |
| Species | Mean Abundance | Mean Infestation Intensity |
|---|---|---|
| Haemaphysalis bispinosa | 2.80 | 7.64 |
| Haemaphysalis intermedia | 1.16 | 3.16 |
| Hyalomma kumari | 0.01 | 0.02 |
| Rhipicephalus (Bo.) annulatus | 2.66 | 7.26 |
| Rhipicephalus (Bo.) microplus | 0.02 | 0.06 |
| Rhipicephalus sanguineus | 0.08 | 0.21 |
| Rhipicephalus simus | 0.05 | 0.13 |
| Rhipicephalus haemaphysaloides | 0.08 | 0.22 |
| Grand Total | 6.85 | 18.71 |
| Species Name | Gene Name | Accession Number | Base Pair |
|---|---|---|---|
| Haemaphysalis bispinosa | Domain II of Voltage gated sodium channel | PX802710 | 148 bp |
| Haemaphysalis intermedia | Domain II of Voltage gated sodium channel | PX802712 | 167 bp |
| Hyalomma kumari | Domain II of Voltage gated sodium channel | PX802713 | 166 bp |
| Rhipicephalus (Bo.) annulatus | Domain II of Voltage gated sodium channel | PX530872 | 163 bp |
| Rhipicephalus (Bo.) microplus | Domain II of Voltage gated sodium channel | PX845415 | 167 bp |
| Rhipicephalus sanguineus | Domain II of Voltage gated sodium channel | PX845414 | 154 bp |
| Rhipicephalus simus | Domain II of Voltage gated sodium channel | PX845413 | 148 bp |
| Rhipicephalus haemaphysaloides | Domain II of Voltage gated sodium channel | PX802711 | 140 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Jasmiminal, K.H.; Ayyanar, E.; Madda, C.; Ramakrishnan, V.K.; Konuganti, H.K.R.; Rahi, M. Species Diversity and Molecular Screening of Pyrethroid Resistance Mediated by the Voltage-Gated Sodium Channel in Ixodid Ticks from Puducherry, India. Pathogens 2026, 15, 577. https://doi.org/10.3390/pathogens15060577
Jasmiminal KH, Ayyanar E, Madda C, Ramakrishnan VK, Konuganti HKR, Rahi M. Species Diversity and Molecular Screening of Pyrethroid Resistance Mediated by the Voltage-Gated Sodium Channel in Ixodid Ticks from Puducherry, India. Pathogens. 2026; 15(6):577. https://doi.org/10.3390/pathogens15060577
Chicago/Turabian StyleJasmiminal, Kaja Hussain, Elango Ayyanar, Chakravarthi Madda, Vignesh Kumar Ramakrishnan, Hari Kishan Raju Konuganti, and Manju Rahi. 2026. "Species Diversity and Molecular Screening of Pyrethroid Resistance Mediated by the Voltage-Gated Sodium Channel in Ixodid Ticks from Puducherry, India" Pathogens 15, no. 6: 577. https://doi.org/10.3390/pathogens15060577
APA StyleJasmiminal, K. H., Ayyanar, E., Madda, C., Ramakrishnan, V. K., Konuganti, H. K. R., & Rahi, M. (2026). Species Diversity and Molecular Screening of Pyrethroid Resistance Mediated by the Voltage-Gated Sodium Channel in Ixodid Ticks from Puducherry, India. Pathogens, 15(6), 577. https://doi.org/10.3390/pathogens15060577