
High Prevalence of Virulence and blaOXA Genes Encoding Carbapenemases Among Acinetobacter baumannii Isolates from Hospitalised Patients in Three Regions of Poland

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Detection of Antibiotic Resistance
2.3. DNA Isolation
2.4. Detection of Resistance and Virulence Genes
2.5. Biofilm Formation Assay
2.6. Statistical Analysis
3. Results
3.1. Resistance to Antibiotics
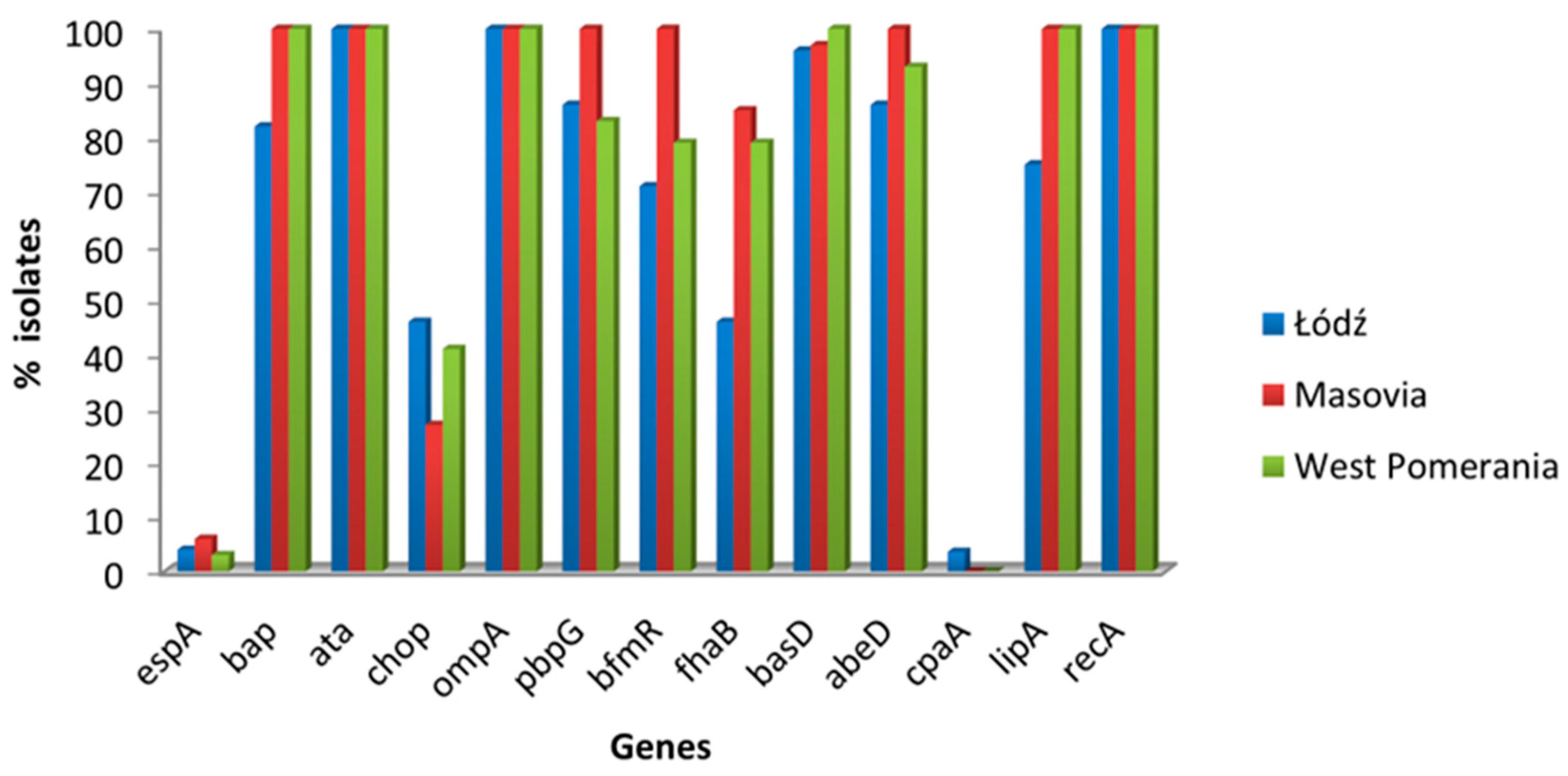
3.2. Virulence-Associated Genes in A. baumannii Isolates
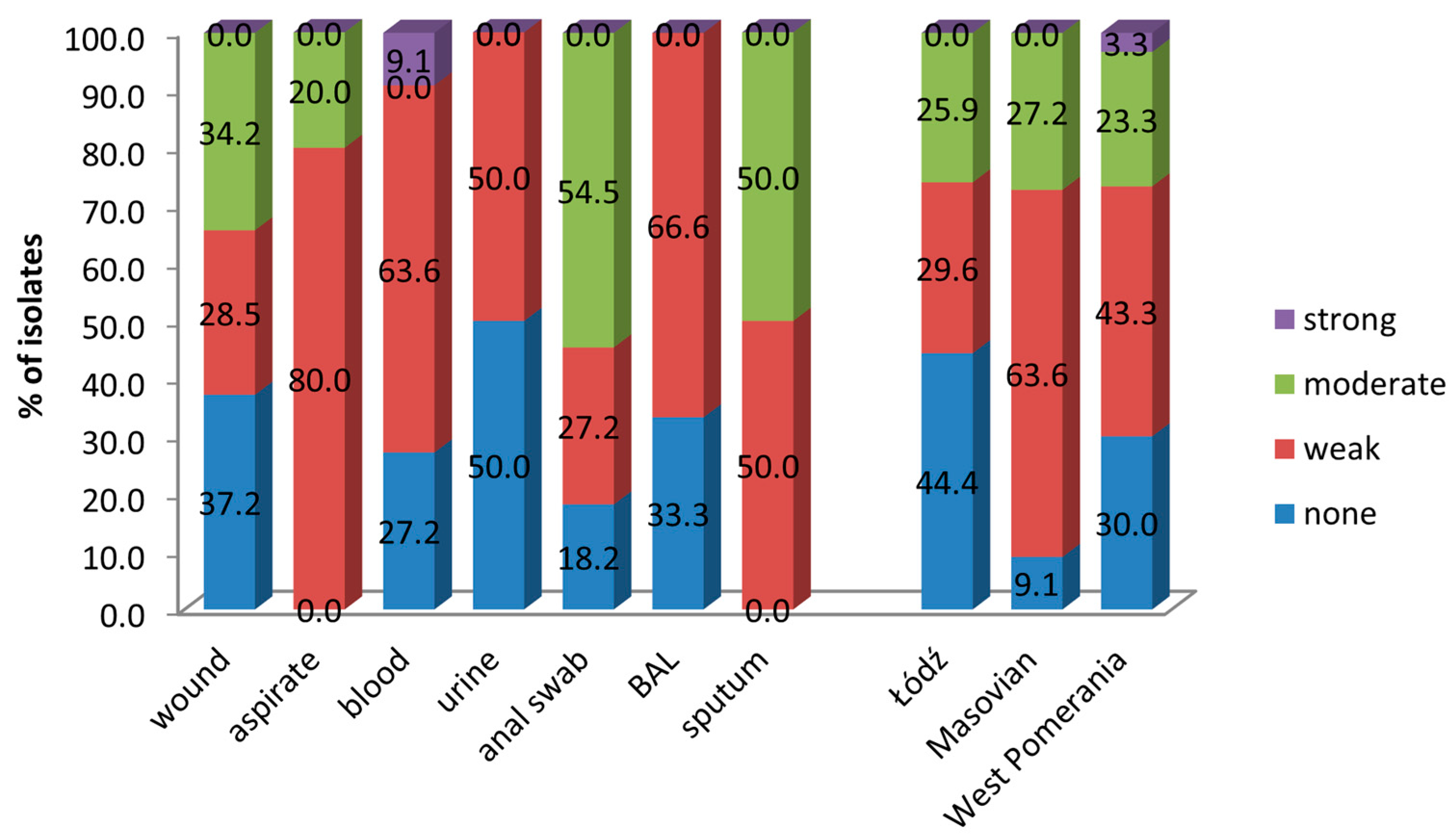
3.3. Biofilm Formation by A. baumannii
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klotz, P.; Higgins, P.G.; Schaubmar, A.R.; Failing, K.; Leidner, U.; Seifert, H.; Scheufen, S.; Semmler, T.; Ewers, C. Seasonal occurrence and carbapenem susceptibility of bovine Acinetobacter baumannii in Germany. Front. Microbiol. 2019, 10, 272. [Google Scholar] [CrossRef] [PubMed]
- La Scola, B.; Raoult, D. Acinetobacter baumannii in human body louse. Emerg. Infect. Dis. 2004, 10, 1671. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; McClean, S. Mapping Global Prevalence of Acinetobacter baumannii and Recent Vaccine Development to Tackle It. Vaccines 2021, 9, 570. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Antimicrobial Resistance in the EU/EEA (EARS-NET) Annual Epidemiological Report for 2023. 2024. Available online: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-eueea-ears-net-annual-epidemiological-report-2023 (accessed on 18 November 2024).
- European Centre for Disease Prevention and Control. Antimicrobial Resistance Surveillance in Europe 2022–2020. 2022. Available online: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-surveillance-europe-2022-2020-data (accessed on 26 January 2022).
- Hafeez, A.; Munir, T.; Najeeb, S.; Rehman, S.; Gilani, M.; Ansari, M.; Saad, N. ICU pathogens: A continuous challenge. J. Coll. Physicians Surg. Pak. 2016, 26, 577–580. [Google Scholar] [PubMed]
- Vincent, J.L.; Rello, J.; Marshall, J.; Silva, E.; Anzueto, A.; Martin, C.D.; Moreno, R.; Lipman, J.; Gomersall, C.; Sakr, Y.; et al. EPIC II Group of Investigators. International study of the prevalence and outcomes of infection in intensive care units. JAMA 2009, 30, 2323–2329. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.; Visca, P.; Towner, K.J. Acinetobacter baumannii: Evolution of a global pathogen. Pathog. Dis. 2014, 71, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Mohd Sazlly Lim, S.; Zainal Abidin, A.; Liew, S.M.; Roberts, J.A.; Sime, F.B. The global prevalence of multidrug-resistance among Acinetobacter baumannii causing hospital-acquired and ventilator-associated pneumonia and its associated mortality: A systematic review and meta-analysis. J. Infect. 2019, 79, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Gedefie, A.; Demsis, W.; Ashagrie, M.; Kassa, Y.; Tesfaye, M.; Tilahun, M.; Bisetegn, H.; Sahle, Z. Acinetobacter baumannii Biofilm Formation and Its Role in Disease Pathogenesis: A Review. Infect. Drug Resist. 2021, 10, 3711–3719. [Google Scholar] [CrossRef] [PubMed]
- Badave, G.K.; Dhananjay, K. Biofilm Producing Multidrug Resistant Acinetobacter baumannii: An Emerging Challenge. J. Clin. Diagn. Res. 2015, 9, DC08–DC10. [Google Scholar] [CrossRef] [PubMed]
- Gaddy, J.A.; Actis, L.A. Regulation of Acinetobacter baumannii biofilm formation. Future Microbiol. 2009, 4, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad Bugs, No Drugs: No ESKAPE! An Update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Brotfain, E.; Borer, A.; Koyfman, L.; Saidel-Odes, L.; Frenkel, A.; Gruenbaum, S.E.; Rosenzweig, V.; Zlotnik, A.; Klein, M. Multidrug resistance acinetobacter bacteremia secondary to ventilator-associated pneumonia: Risk factors and outcome. J. Intensive Care Med. 2017, 32, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Kempf, M.; Rolain, J.M. Emergence of resistance to carbapenems in Acinetobacter baumannii in Europe: Clinical impact and therapeutic options. Int. J. Antimicrob. Agents 2012, 39, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Smolyakov, R.; Borer, A.; Riesenberg, K.; Schlaeffer, F.; Alkan, M.; Porath, A.; Rimar, D.; Almog, Y.; Gilad, J. Nosocomial multi-drug resistant Acinetobacter baumannii bloodstream infection: Risk factors and outcome with ampicillin-sulbactam treatment. J. Hosp. Infect. 2003, 54, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Shao, N.; Zheng, J. Integrated co-functional network analysis on the resistance and virulence features in Acinetobacter baumannii. Front. Microbiol. 2020, 11, 2677. [Google Scholar] [CrossRef] [PubMed]
- WHO. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug Resistant Bacterial Infections, Including Tuberculosis. 2017. Available online: https://www.who.int/publications/i/item/WHO-EMP-IAU-2017.12 (accessed on 4 September 2017).
- Wu, H.J.; Xiao, Z.G.; Lv, X.J.; Huang, H.T.; Liao, C.; Hui, C.Y.; Xu, Y.; Li, H.F. Drug-resistant Acinetobacter baumannii: From molecular mechanisms to potential therapeutics (Review). Exp. Ther. Med. 2023, 25, 209. [Google Scholar] [CrossRef] [PubMed]
- Ambler, R.P. The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1980, 289, 321–331. [Google Scholar]
- Ramirez, M.S.; Bonomo, R.A.; Tolmasky, M.E. Carbapenemases: Transforming Acinetobacter baumannii into a Yet More Dangerous Menace. Biomolecules 2020, 6, 720. [Google Scholar] [CrossRef] [PubMed]
- EUCAST Expected Resistance Phenotypes; Version 15.0; European Committee on Antimicrobial Susceptibility Testing: Växjö, Sweden, 2025; Available online: www.eucast.org (accessed on 1 January 2025).
- CLSI M100; CLSI Performance Standards for Antimicrobial Susceptibility Testing, 34th ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2024.
- Cerezales, M.; Biniossek, L.; Gerson, S.; Xanthopoulou, K.; Wille, J.; Wohlfarth, E.; Kaase, M.; Seifert, H.; Higgins, P.G. Novel multiplex PCRs for detection of the most prevalent carbapenemase genes in Gram-negative bacteria within Germany. J. Med. Microbiol. 2021, 70, 3. [Google Scholar] [CrossRef] [PubMed]
- Park, S.M.; Suh, J.W.; Ju, Y.K.; Kim, J.Y.; Kim, S.B.; Sohn, J.W.; Yoon, Y.K. Molecular and virulence characteristics of carbapenem-resistant Acinetobacter baumannii isolates: A prospective cohort study. Sci. Rep. 2023, 13, 19536. [Google Scholar] [CrossRef] [PubMed]
- Piechota, M.; Kot, B.; Frankowska-Maciejewska, A.; Grużewska, A.; Woźniak-Kosek, A. Biofilm formation by methicillin-resistant and methicillin-sensitive Staphylococcus aureus strains from hospitalized patients in Poland. Biomed. Res. Int. 2018, 27, 4657396. [Google Scholar]
- Russo, A.; Bassetti, M.; Ceccarelli, G.; Carannante, N.; Losito, A.R.; Bartoletti, M.; Corcione, S.; Granata, G.; Santoro, A.; Giacobbe, D.R.; et al. Bloodstream infections caused by carbapenem-resistant Acinetobacter baumannii: Clinical features, therapy and outcome from a multicenter study. J. Infect. 2019, 79, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Ayobami, O.; Willrich, N.; Suwono, B.; Eckmanns, T.; Markwart, R. The epidemiology of carbapenem-non-susceptible Acinetobacter species in Europe: Analysis of EARS-Net data from 2013 to 2017. Antimicrob. Resist Infect. Control. 2020, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Kasperski, T.; Romaniszyn, D.; Jachowicz-Matczak, E.; Pomorska-Wesołowska, M.; Wójkowska-Mach, J.; Chmielarczyk, A. Extensive Drug Resistance of Strong Biofilm-Producing Acinetobacter baumannii Strains Isolated from Infections and Colonization Hospitalized Patients in Southern Poland. Pathogens 2023, 12, 975. [Google Scholar] [CrossRef] [PubMed]
- Betrosian, A.P.; Frantzeskaki, F.; Xanthaki, A.; Georgiadis, G. High-dose ampicillin-sulbactam as an alternative treatment of late-onset VAP from multidrug-resistant Acinetobacter baumannii. Scand. J. Infect. Dis. 2007, 39, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.Y.; Lee, S.H.; Lee, S.Y.; Yang, S.; Noh, H.; Chung, E.K.; Lee, J.I. Antimicrobials for the treatment of drug-re sistant Acinetobacter baumannii pneumonia in criti cally ill patients: A systemic review and Bayesian network meta-analysis. Crit. Care Crit. Care 2017, 21, 319. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shu, Y.; Zhu, F.; Feng, B.; Zhang, Z.; Liu, L.; Wang, G. Comparative efficacy and safety of combination therapy with high-dose sulbactam or colistin with additional antibacterial agents for multiple drug-resistant and extensively drug-resistant Acinetobacter baumannii infections: A systematic review and network. J. Glob. Antimicrob. Resist. 2021, 24, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Lenhard, J.R.; Smith, N.M.; Bulman, Z.P.; Tao, X.; Thamlikitkul, V.; Shin, B.S.; Nation, R.L.; Li, J.; Bulitta, J.B.; Tsuji, B.T. High-dose ampicillin-sulbactam combinations combat polymyxin-resistant Acinetobacter baumannii in a hollow-fiber infection model. Antimicrob. Agents Chemother. 2017, 61, e01268–e1316. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.; Al-Saryi, N.; Al-Kadmy, I.M.S. Multidrug-resistant Acinetobacter baumannii as an emerging concern in hospitals. Mol. Biol. Rep. 2021, 48, 6987–6998. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.A.; Amyes, S.G. Oxa β-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [PubMed]
- Nowak, P.; Paluchowska, P.M.; Budak, A. Co-occurrence of carbapenem and aminoglycoside resistance genes among multidrug-resistant clinical isolates of Acinetobacter baumannii from Cracow, Poland. Med. Sci. Monit. Basic Res. 2014, 20, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Djahmi, N.; Dunyach-Remy, C.; Pantel, A.; Dekhil, M.; Sotto, A.; Lavigne, J.P. Epidemiology of carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Mediterranean countries. Biomed. Res. Int. 2014, 2014, 305784. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.C.; Huang, W.C.; Huang, T.W.; Wang, H.Y.; Lai, J.F.; Chen, T.L.; Lauderdale, T.L. Molecular epidemiology of emerging bla (OXA-23-Like)- and bla (OXA-24-Like)-carrying Acinetobacter baumannii in Taiwan. Antimicrob. Agents Chemother. 2018, 62, e01215-17. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, E.A.; Paula, G.R.; Mondino, P.J.J.; Chagas, T.P.G.; Mondino, S.S.B.; Mendonça-Souza, C.R.V. High rate of detection of OXA-23-producing Acinetobacter from two general hospitals in Brazil. Rev. Soc. Bras. Med. Trop. 2019, 52, e20190243. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Naas, T.; Nordmann, P. Diversity, epidemiology, and genetics of class D beta-lactamases. Antimicrob. Agents Chemother. 2010, 54, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, K.; Thirunarayan, M.A.; Krishnan, P. Coexistence of blaOXA-23 with blaNDM-1 and armA in clinical isolates of Acinetobacter baumannii from India. J. Antimicrob. Chemother. 2010, 65, 2253–2254. [Google Scholar] [CrossRef] [PubMed]
- Girija, S.A.; Jayaseelan, V.P.; Arumugam, P. Prevalence of VIM- and GIM-producing Acinetobacter baumannii from patients with severe urinary tract infection. Acta. Microbiol. Immunol. Hung. 2018, 65, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Azizi, O.; Shakibaie, M.R.; Badmasti, F.; Modarresi, F.; Ramazanzadeh, R.; Mansouri, S.; Shahcheraghi, F. Class 1 integrons in non-clonal multidrug-resistant Acinetobacter baumannii from Iran, description of the new blaIMP-55 allele in In1243. J. Med. Microbiol. 2016, 65, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, P.C.; Monteiro, A.S.; Marques, S.G.; Monteiro, S.G.; Monteiro-Neto, V.; Coqueiro, M.M.; Marques, A.C.; de Jesus Gomes Turri, R.; Santos, S.G.; Bomfim, M.R. Phenotypic and molecular detection of the blaKPC gene in clinical isolates from inpatients at hospitals in Sao Luis, MA, Brazil. BMC Infect. Dis. 2016, 16, 737. [Google Scholar] [CrossRef] [PubMed]
- Celik, B. Evaluation of the correlation between biofilm formation and drug resistance in clinical isolates of Acinetobacter baumannii. Int. J. Pathog. Res. 2020, 5, 16–27. [Google Scholar] [CrossRef]
- Asaad, A.M.; Soma, S.A.; Ajlan, E.; Awad, S.M. Epidemiology of biofilm producing Acinetobacter baumannii nosocomial isolates from a Tertiary Care Hospital in Egypt: A cross-sectional study. Infect. Drug Resist. 2021, 14, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Li, H.; Zhang, C.; Liang, B.; Li, J.; Wang, L.; Du, X.; Liu, X.; Qiu, S.; Song, H. Relationship between antibiotic resistance, biofilm formation, and biofilm-specific resistance in Acinetobacter baumannii. Front. Microbiol. 2016, 7, 483. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Baño, J.; Martí, S.; Soto, S.; Fernández-Cuenca, F.; Cisneros, J.M.; Pachón, J.; Pascual, A.; Martínez-Martínez, L.; McQueary, C.; Actis, L.A.; et al. Biofilm formation in Acinetobacter baumannii: Associated features and clinical implications. Clin. Microbiol. Infect. 2008, 14, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.H.; Hyun, S.H.; Lee, J.Y.; Lee, J.S.; Lee, Y.S.; Kim, S.A.; Chae, J.P.; Yoo, S.M.; Lee, J.C. Acinetobacter baumannii outer membrane protein A targets the nucleus and induces cytotoxicity. Cell. Microbiol. 2008, 10, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.H.; Lee, E.Y.; Lee, Y.C.; Park, T.I.; Kim, H.J.; Hyun, S.H.; Kim, S.A.; Lee, S.-K.; Lee, J.C. Outer membrane protein 38 of Acinetobacter baumannii localizes to the mitochondria and induces apoptosis of epithelial cells. Cell. Microbiol. 2005, 7, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Encinales, V.; Álvarez-Marín, R.; Pachón-Ibáñez, M.E.; Fernández-Cuenca, F.; Pascual, A.; Garnacho-Montero, J.; Martínez-Martínez, L.; Vila, J.; Tomás, M.M.; Miguel Cisneros, J.; et al. Overproduction of outer membrane protein a by Acinetobacter baumannii as a risk factor for nosocomial pneumonia, bacteremia, and mortality rate increase. J. Infect. Dis. 2017, 215, 966–974. [Google Scholar] [PubMed]
- Bentancor, L.V.; Camacho-Peiro, A.; Bozkurt-Guzel, C.; Pier, G.B.; Maira-Litrán, T. Identification of Ata, a multifunctional trimeric autotransporter of Acinetobacter baumannii. J. Bacteriol. 2012, 194, 3950–3960. [Google Scholar] [CrossRef] [PubMed]
- Weidensdorfer, M.; Ishikawa, M.; Hori, K.; Linke, D.; Djahanschiri, B.; Iruegas, R.; Ebersberger, I.; Riedel-Christ, S.; Enders, G.; Leukert, L.; et al. The Acinetobacter trimeric autotransporter adhesin Ata controls key virulence traits of Acinetobacter baumannii. Virulence 2019, 1, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Longo, F.; Vuotto, C.; Donelli, G. Biofilm formation in Acinetobacter baumannii. New Microbiol. 2014, 37, 119–127. [Google Scholar] [PubMed]
- Thummeepak, R.; Kongthai, P.; Leungtongkam, U.; Sitthisak, S. Distribution of Virulence Genes Involved in Biofilm Formation in Multi-Drug Resistant Acinetobacter baumannii Clinical Isolates. Int. Microbiol. 2016, 19, 121–129. [Google Scholar] [PubMed]
- Ghasemi, E.; Ghalavand, Z.; Goudarzi, H.; Yeganeh, F.; Hashemi, A.; Dabiri, H.; Mirsamadi, E.S.; Foroumand, M. Phenotypic and Genotypic Investigation of Biofilm Formation in Clinical and Environmental Isolates of Acinetobacter baumannii. Arch. Clin. Infect. Dis. 2018, 13, 12914. [Google Scholar] [CrossRef]
- Depka, D.; Bogiel, T.; Rzepka, M.; Gospodarek-Komkowska, E. The Prevalence of virulence factor genes among carbapenem-non-susceptible Acinetobacter baumannii clinical strains and their usefulness as potential molecular biomarkers of infection. Diagnostics 2023, 13, 1036. [Google Scholar] [CrossRef] [PubMed]
- Palethorpe, S.; Farrow III, J.M.; Wells, G.; Milton, M.E.; Actis, L.A.; Cavanagh, J.; Pesci, E.C. Acinetobacter baumannii regulates its stress responses via the BfmRS two-component regulatory system. J. Bacteriol. 2022, 204, e00494-21. [Google Scholar] [CrossRef] [PubMed]
- Tomaras, A.P.; Flagler, M.J.; Dorsey, C.W.; Gaddy, J.A.; Actis, L.A. Characterization of a two-component regulatory system from Acinetobacter baumannii that controls biofilm formation and cellular morphology. Microbiology 2008, 154, 3398–3409. [Google Scholar] [CrossRef] [PubMed]
- Marr, C.M.; MacDonald, U.; Trivedi, G.; Chakravorty, S.; Russo, T.A. An evaluation of BfmR-regulated antimicrobial resistance in the extensively drug resistant (XDR) Acinetobacter baumannii strain HUMC1. Front. Microbiol. 2020, 11, 2688. [Google Scholar] [CrossRef] [PubMed]
- Geisinger, E.; Isberg, R.R. Antibiotic modulation of capsular exopolysaccharide and virulence in Acinetobacter baumannii. PLoS Pathog. 2015, 11, e1004691. [Google Scholar] [CrossRef] [PubMed]
- Khamari, B.; Lama, M.; Pulusu, C.P.; Biswal, A.P.; Lingamallu, S.M.; Mukkirla, B.S.; Sahoo, A.K.; Dash, H.S.N.; Sharda, R.; Kumar, P.; et al. Molecular analyses of biofilm-producing clinical Acinetobacter baumannii isolates from a south Indian Tertiary Care Hospital. Med. Princ. Pract. 2020, 29, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Mozafari, H.; Mirkalantari, S.; Kalani, B.S.; Amirmozafari, N. Prevalence Determination of Virulence Related and Biofilm Formation Genes in Acinetobacter baumannii Isolates from Clinical Respiratory Samples in Imam Khomeini Hospital, Tehran, Iran in 2018. Iran. J. Med. Microbiol. 2021, 15, 266–280. [Google Scholar] [CrossRef]
- Pérez, A.; Merino, M.; Rumbo-Feal, S.; Álvarez-Fraga, L.; Vallejo, J.A.; Beceiro, A.; Ohneck, E.J.; Mateos, J.; Fernández-Puente, P.; Actis, L.A.; et al. The FhaB/FhaC two-partner secretion system is involved in adhesion of Acinetobacter baumannii AbH12O-A2 strain. Virulence 2016, 8, 959–974. [Google Scholar] [CrossRef] [PubMed]
- Astaneh, S.D.A.; Rasooli, I.; Gargari, S.L.M. The role of filamentous hemagglutinin adhesin in adherence and biofilm formation in Acinetobacter baumannii ATCC19606 (T). Microb. Pathog. 2014, 74, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.A.; Carlino-MacDonald, U.; Alvarado, C.L.; Davies, C.J.; Barnes, O.; Trivedi, G.; Mathur, P.; Hutson, A.; Adams, F.G.; Zang, M.; et al. Penicillin binding Pprotein 7/8 is a potential drug target in carbapenem-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2022, 67, e0103322. [Google Scholar] [CrossRef] [PubMed]
- Hasan, T.; Choi, C.H.; Oh, M.H. Genes involved in the biosynthesis and transport of acinetobactin in Acinetobacter baumannii. Genomics Inform. 2015, 13, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Porbaran, M.; Tahmasebi, H.; Arabestani, M.R. A comprehensive study of the relationship between the production of β-lactamase enzymes and iron/siderophore uptake regulatory genes in clinical isolates of Acinetobacter baumannii. Int. J. Microbiol. 2021, 17, 5565537. [Google Scholar] [CrossRef] [PubMed]
- Haurat, M.F.; Scott, N.E.; Venanzio, G.D.; Lopez, J.; Pluvinage, B.; Boraston, A.B.; Ferracane, M.J.; Feldman, M.F. The glycoprotease CpaA Secreted by medically relevant Acinetobacter species targets multiple O-linked host glycoproteins. mBio 2020, 11, e02033-20. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.A.; Luke, N.R.; Beanan, J.M.; Olson, R.; Sauberan, S.L.; MacDonald, U.; Schultz, L.W.; Umland, T.C.; Campagnari, A.A. The K1 Capsular polysaccharide of Acinetobacter baumannii strain 307-0294 is a major virulence factor. Infect. Immun. 2010, 78, 3993–4000. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.L.; Waack, U.; Smith, S.; Mobley, H.; Sandkvist, M. Acinetobacter baumannii is dependent on the type II secretion system and its substrate LipA for lipid utilization and in vivo fitness. J. Bacteriol. 2016, 198, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.W.; Park, S.M.; Ju, Y.K.; Yang, K.S.; Kim, J.Y.; Kim, S.B.; Sohn, J.W.; Yoon, Y.K. Clinical and molecular predictors of mortality in patients with carbapenem-resistant Acinetobacter baumannii bacteremia: A retrospective cohort study. J. Microbiol. Immunol. Infect. 2024, 57, 148–155. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Isolates | Number of Isolates from the Voivodeship: | |||
|---|---|---|---|---|
| Łódź | West Pomeranian | Masovian | Total | |
| Wound | 18 | 9 | 8 | 35 |
| Bronchial aspirate | 0 | 0 | 15 | 15 |
| Blood | 4 | 4 | 3 | 11 |
| Anal swab | 0 | 6 | 5 | 11 |
| Urine | 2 | 4 | 2 | 8 |
| BAL | 1 | 5 | 0 | 6 |
| Sputum | 2 | 2 | 0 | 4 |
| Total | 27 | 30 | 33 | 90 |
| Resistance Phenotypes (No. of Antibiotics) | Number (%) of Isolates from: | |||
|---|---|---|---|---|
| Masovia | West Pomerania | Łódź | Total | |
| (12) | ||||
| SAM, TZP, CAZ, FEP, IPM, MEM, GM, AK, TOB, CIP, LEV, SXT | 21 (63.6) | 18 (60.0) | 8 (29.6) | 47 (52.2) |
| (11) | ||||
| TZP, CAZ, FEP, IPM, MEM, GM, TOB, CIP, LEV, SXT | 9 (27.3) | 6 (20.0) | 3 (11.1) | 18 (20.0) |
| SAM, TZP, CAZ, FEP, IPM, MEM, GM, TOB, CIP, LEV, SXT | 0 (0.0) | 2 (6.7) | 0 (0.0) | 2 (2.2) |
| SAM, TZP, CAZ, IPM, MEM, GM, AK, TOB, CIP, LEV, SXT | 0 (0.0) | 1 (3.3) | 1 (3.7) | 2 (2.2) |
| (10) | ||||
| SAM, TZP, CAZ, FEP, IPM, MEM, GM, CIP, LEV, SXT | 2 (6.1) | 1 (3.3) | 0 (0.0) | 3 (3.3) |
| SAM, TZP, CAZ, FEP, GM, AK, TOB, CIP, LEV, SXT | 0 (0.0) | 0 (0.0) | 1 (3.7) | 1 (1.1) |
| TZP, CAZ, IPM, MEM, GM, AK, TOB, CIP, LEV, SXT | 0 (0.0) | 2 (6.7) | 2 (7.4) | 4 (4.4) |
| (9) | ||||
| TZP, CAZ, FEP, GM, AK, TOB, CIP, LEV, SXT | 0 (0.0) | 0 (0.0) | 2 (7.4) | 2 (2.2) |
| TZP, CAZ, FEP, IPM, MEM, GM, CIP, LEV, SXT | 1 (3.0) | 0 (0.0) | 0 (0.0) | 1 (1.1) |
| (6) | ||||
| CAZ, GM, AK, CIP, LEV, SXT | 0 (0.0) | 0 (0.0) | 1 (3.7) | 1 (1.1) |
| (5) | ||||
| SAM, TZP, CAZ, FEP, TOB | 0 (0.0) | 0 (0.0) | 1 (3.7) | 1 (1.1) |
| (1) | ||||
| TOB | 0 (0.0) | 0 (0.0) | 1 (3.7) | 1 (1.1) |
| Source | Virulence Genes, No. (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| espA | bap | ata | chop | ompA | pbpG | bfmR | fhaB | abeD | basD | cpaA | lipA | recA | |
| Wound n = 35 | 1 (2.8) | 32 (91.4) | 35 (100) | 15 (42.8) | 35 (100) | 31 (88.5) | 27 (77.1) | 21 (60.0) | 30 (85.7) | 34(97.1) | 1 (4.1) | 32 (91.4) | 35 (100) |
| Bronchial aspirate n = 15 | 1 (6.6) | 15 (100) | 15 (100) | 3 (20.0) | 15 (100) | 15 (100) | 15 (100) | 12 (80.0) | 15 (100) | 14(93.3) | 0 (0.0) | 15 (100) | 15 (100) |
| Anal swab n = 11 | 0 (0.0) | 11 (100) | 11 (100) | 7 (63.6) | 11 (100) | 11 (100) | 11 (100) | 11 (100) | 11 (100) | 11 (100) | 0 (0.0) | 11 (100) | 11 (100) |
| Blood n = 11 | 0 (0.0) | 11 (100) | 11 (100) | 4 (36.3) | 11 (100) | 9 (81.8) | 10 (90.9) | 8 (72.7) | 11 (100) | 11 (100) | 0 (0.0) | 10 (90.9) | 11 (100) |
| Urine n = 8 | 1 (12.5) | 8 (100) | 8 (100) | 1 (12.5) | 8 (100) | 7 (87.5) | 6 (75.0) | 6 (75.0) | 7 (87.5) | 8 (100) | 0 (0.0) | 8 (100) | 8 (100) |
| BAL n = 6 | 1 (16.6) | 6 (100) | 6 (100) | 2 (33.3) | 6 (100) | 4 (66.6) | 3 (50.0) | 4 (66.6) | 6 (100) | 6 (100) | 0 (0.0) | 6 (100) | 6 (100) |
| Sputum n = 4 | 0 (0.0) | 2 (50.0) | 4 (100) | 2 (50.0) | 4 (100) | 4 (100) | 4 (100) | 2 (50.0) | 4 (100) | 4 (100) | 0 (0.0) | 2 (50.0) | 4 (100) |
| Total | 4 (4.4) | 85 (94.4) | 90 (100) | 34 (37.7) | 90 (100) | 81 (90.0) | 76 (84.4) | 64 (71.1) | 84 (93.3) | 88 (97.7) | 1 (1.1) | 83 (92.2) | 90 (100) |
| Source | No. (%) of Resistant Isolates to Antimicrobials | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SAM | TZP | CAZ | FEP | IPM | MEM | GM | AK | TOB | CIP | LEV | SXT | |
| Wound n = 35 | 17 (48.6) | 28 (80.0) | 29 (82.9) | 23 (65.7) | 25 (71.4) | 25 (71.4) | 29 (82.8) | 26 (74.3) | 27 (77.1) | 29 (82.8) | 29 (82.8) | 29 (82.8) |
| Bronchial aspirate n = 15 | 11 (73.3) | 15 (100) | 15 (100) | 15 (100) | 15 (100) | 15 (100) | 15 (100) | 14 (93.3) | 14 (93.3) | 15 (100) | 15 (100) | 15 (100) |
| Anal swab n = 11 | 9 (81.8) | 11 (100) | 11 (100) | 11 (100) | 11 (100) | 11 (100) | 11 (100) | 10 (90.9) | 11 (100) | 11 (100) | 11 (100) | 11 (100) |
| Blood n = 11 | 9 (81.8) | 11 (100) | 11 (100) | 11 (100) | 11 (100) | 11 (100) | 11 (100) | 10 (90.9) | 10 (90.9) | 11 (100) | 11 (100) | 11 (100) |
| Urine n = 8 | 5 (62.5) | 8 (100) | 8(100) | 8(100) | 8 (100) | 8 (100) | 8(100) | 8(100) | 8(100) | 8(100) | 8(100) | 8(100) |
| BAL n = 6 | 5 (83.3) | 6 (100) | 6(100) | 5 (83.3) | 5 (83.3) | 5(83.3) | 5 (83.3) | 5 (83.3) | 6(100) | 5 (83.3) | 5 (83.3) | 5 (83.3) |
| Sputum n = 4 | 0(0.0) | 2 (50.0) | 2 (50.0) | 2 (50.0) | 2 (50.0) | 2(50.0) | 2 (50.0) | 2 (50.0) | 2 (50.0) | 2 (50.0) | 2 (50.0) | 2 (50.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szemraj, M.; Piechota, M.; Olszowiec, K.; Wicha, J.; Pruss, A.; Sienkiewicz, M.; Witeska, M.; Szweda, P.; Kot, B. High Prevalence of Virulence and blaOXA Genes Encoding Carbapenemases Among Acinetobacter baumannii Isolates from Hospitalised Patients in Three Regions of Poland. Pathogens 2025, 14, 731. https://doi.org/10.3390/pathogens14080731
Szemraj M, Piechota M, Olszowiec K, Wicha J, Pruss A, Sienkiewicz M, Witeska M, Szweda P, Kot B. High Prevalence of Virulence and blaOXA Genes Encoding Carbapenemases Among Acinetobacter baumannii Isolates from Hospitalised Patients in Three Regions of Poland. Pathogens. 2025; 14(8):731. https://doi.org/10.3390/pathogens14080731
Chicago/Turabian StyleSzemraj, Magdalena, Małgorzata Piechota, Kamila Olszowiec, Jolanta Wicha, Agata Pruss, Monika Sienkiewicz, Małgorzata Witeska, Piotr Szweda, and Barbara Kot. 2025. "High Prevalence of Virulence and blaOXA Genes Encoding Carbapenemases Among Acinetobacter baumannii Isolates from Hospitalised Patients in Three Regions of Poland" Pathogens 14, no. 8: 731. https://doi.org/10.3390/pathogens14080731
APA StyleSzemraj, M., Piechota, M., Olszowiec, K., Wicha, J., Pruss, A., Sienkiewicz, M., Witeska, M., Szweda, P., & Kot, B. (2025). High Prevalence of Virulence and blaOXA Genes Encoding Carbapenemases Among Acinetobacter baumannii Isolates from Hospitalised Patients in Three Regions of Poland. Pathogens, 14(8), 731. https://doi.org/10.3390/pathogens14080731