
Serum Amyloid A3 Expression Is Enhanced by Gram-Negative Bacterial Stimuli in Bovine Endometrial Epithelial Cells

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells
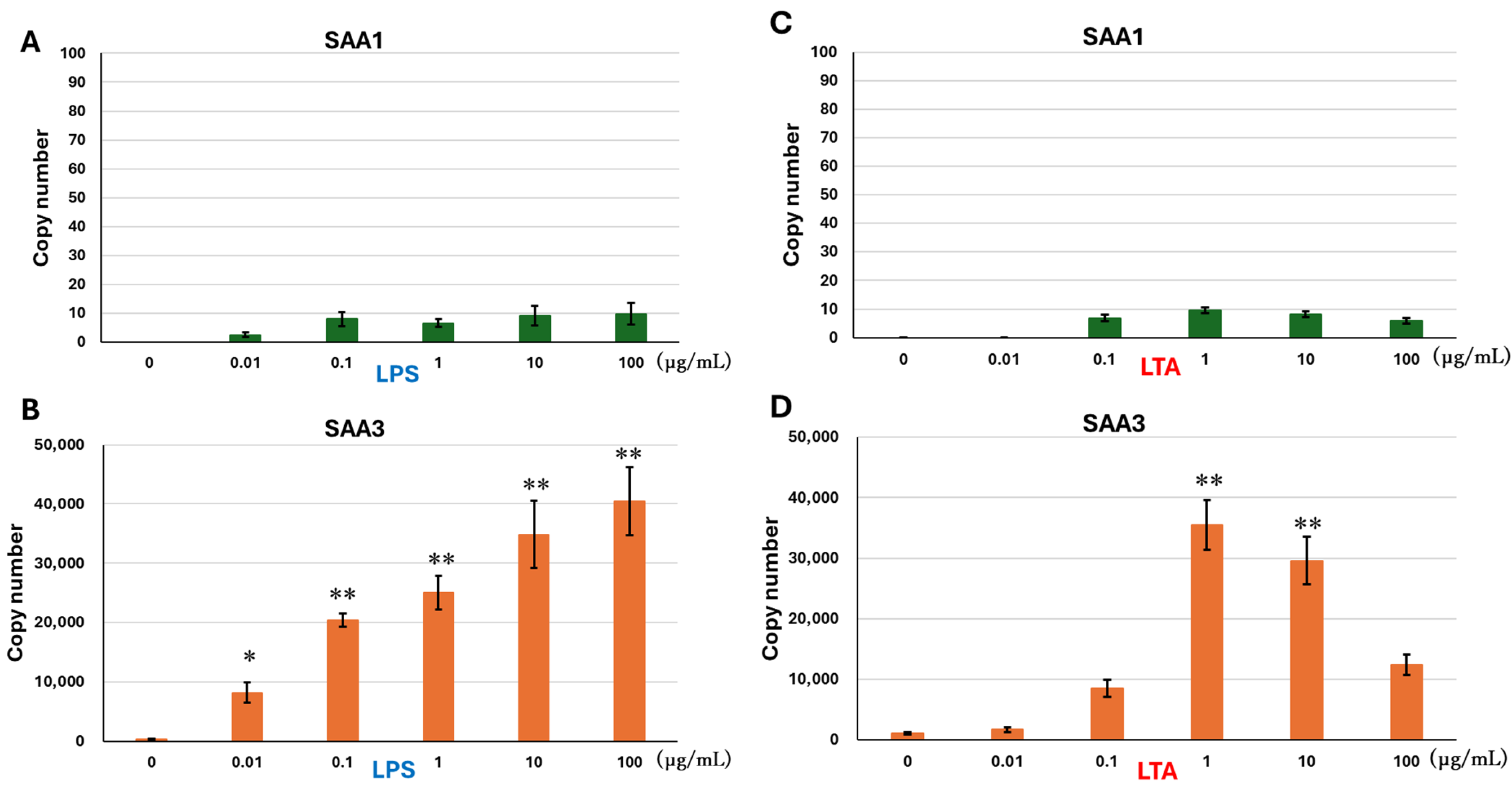
2.2. Treatment with LPS and LTA for mRNA Expression Analysis
2.3. RNA Extraction and cDNA Synthesis
2.4. Cloning and Sequencing
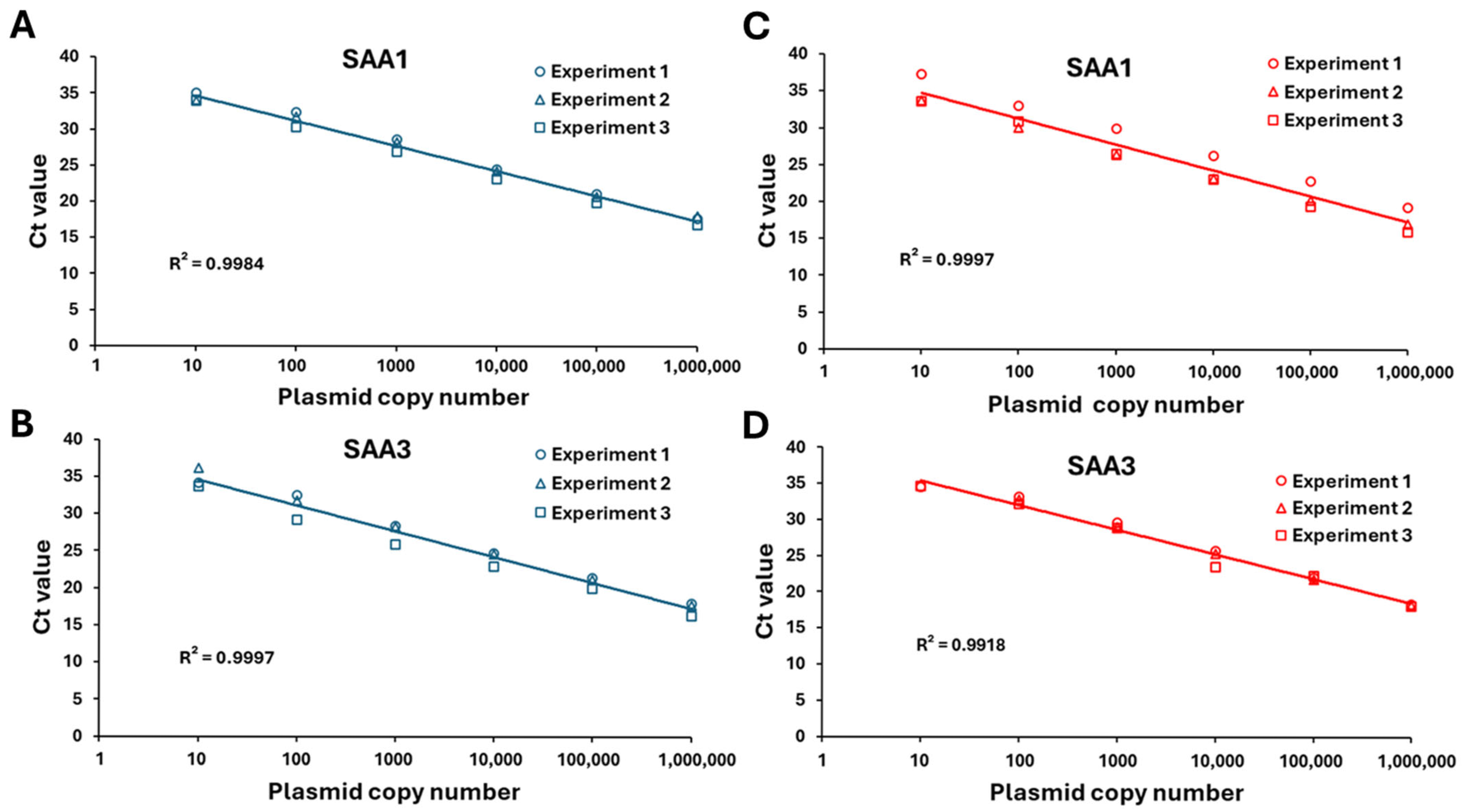
2.5. Determination of Linear Range Using Plasmid-Derived Gene Standards
2.6. Absolute Quantification in qPCR
2.7. Standard Curves for Measuring SAA1 and SAA3 mRNA Copy Numbers Through qPCR
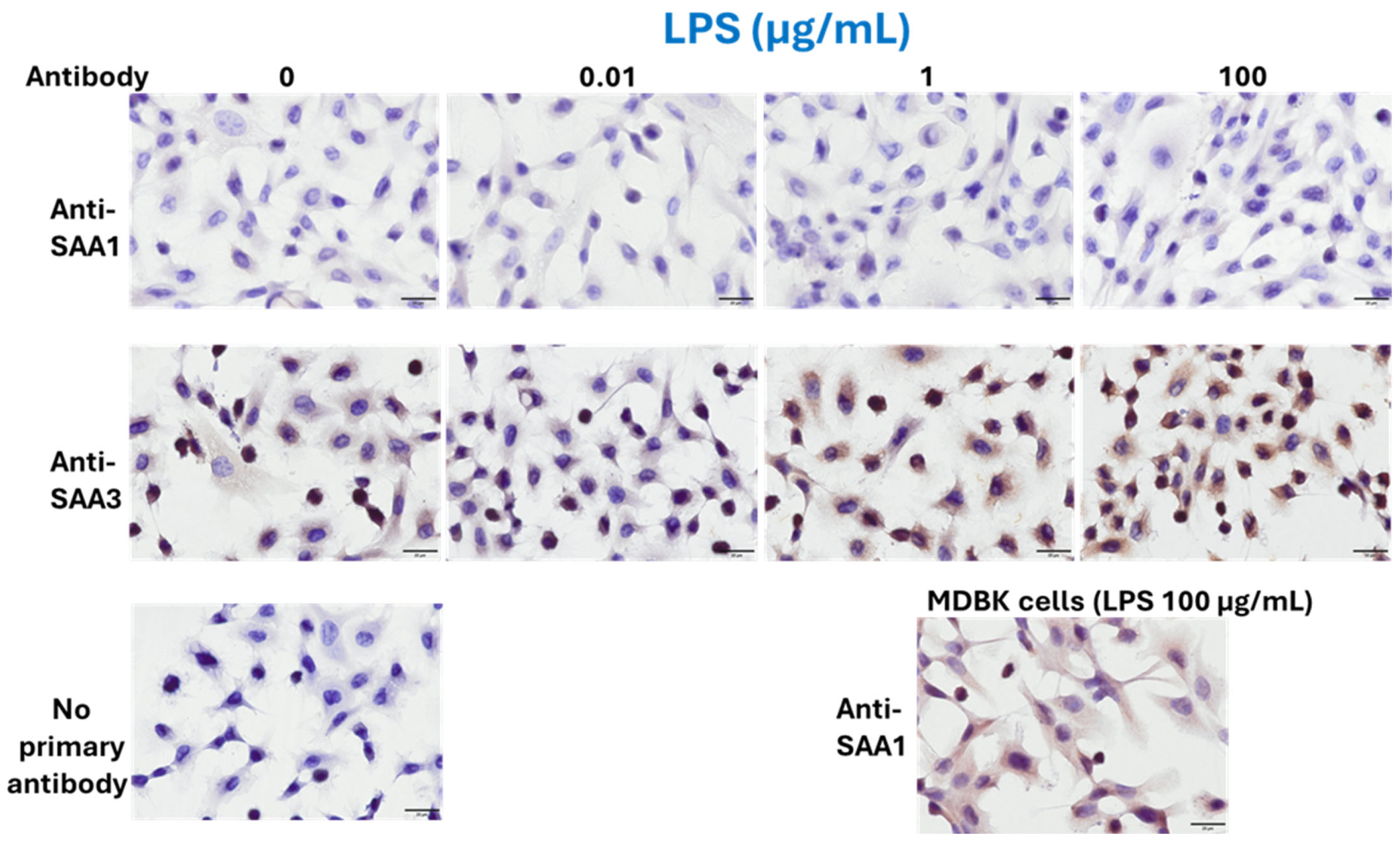
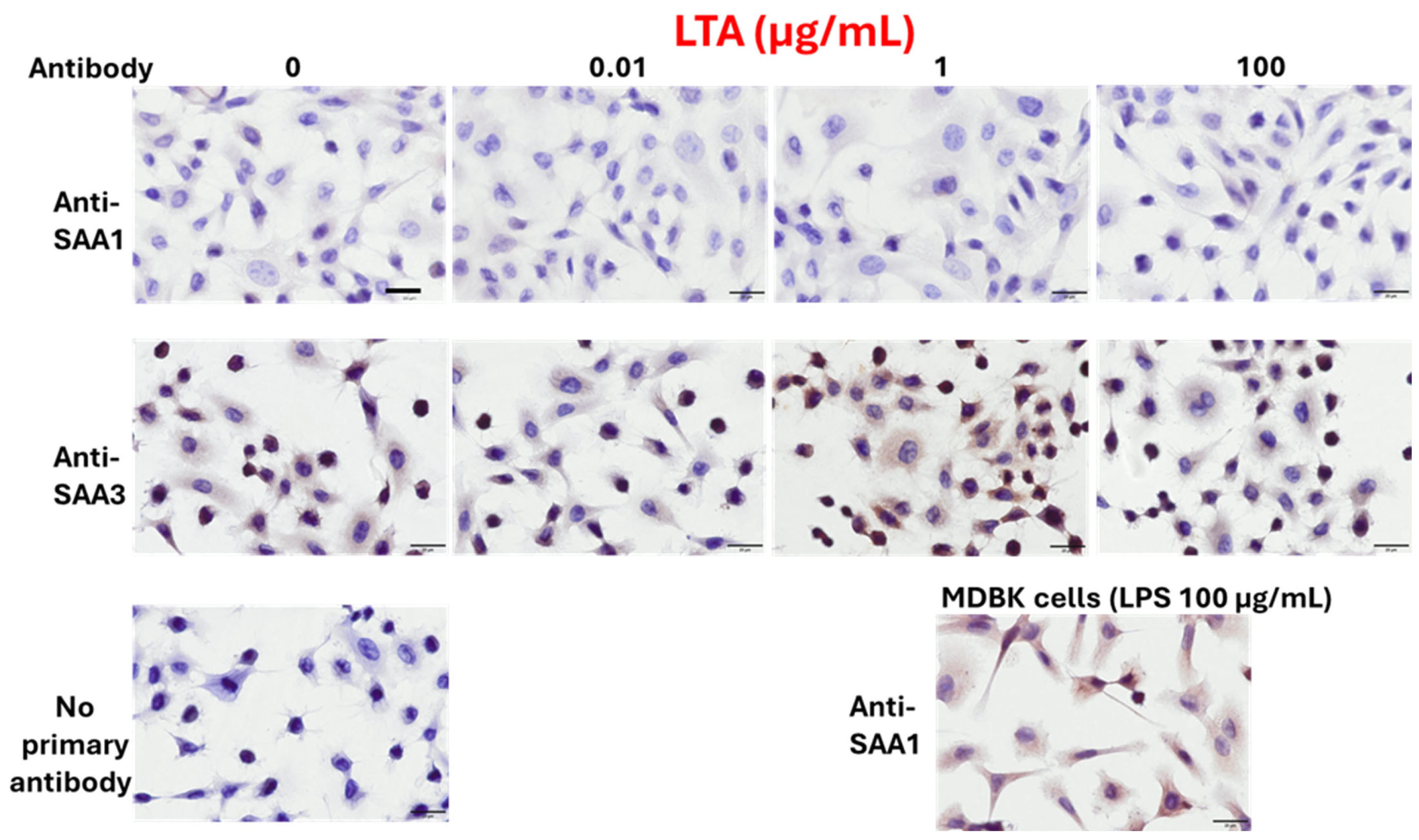
2.8. Treatment with LPS and LTA for ICC
2.9. ICC
2.10. Statistical Analysis
3. Results
3.1. mRNA Expressions of SAA1 and SAA3 in BEnEpCs Treated with LPS and LTA
3.2. Protein Expressions of SAA1 and SAA3 in BEnEpCs Treated with LPS and LTA
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Amyloid A |
| BEnEpC | Bovine endometrial epithelial cell |
| BME | Bovine mammary epithelial |
| BSA | Bovine serum albumin |
| ICC | Immunocytochemistry |
| IL | Interleukin |
| LPS | Lipopolysaccharide |
| LTA | Lipoteichoic acid |
| MDBK | Madin–Darby bovine kidney |
| MUC2 | Mucin2 |
| PBS | Phosphate-buffered saline |
| PCR | Polymerase chain reaction |
| qPCR | Quantitative real-time polymerase chain reaction |
| SAA | Serum amyloid A |
Appendix A
References
- Sheldon, I.M.; Lewis, G.S.; LeBlanc, S.; Gilbert, R.O. Defining postpartum uterine disease in cattle. Theriogenology 2006, 65, 1516–1530. [Google Scholar] [CrossRef]
- LeBlanc, S.J. Postpartum uterine disease and dairy herd reproductive performance: A review. Vet. J. 2008, 176, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Aitken, S.L.; Corl, C.M.; Sordillo, L.M. Immunopathology of the postpartum cow: Insights into endometrial and mammary inflammation. J. Dairy Sci. 2016, 99, 2930–2940. [Google Scholar]
- Bicalho, M.L.S.; Machado, V.S.; Higgins, C.H.; Lima, F.S.; Bicalho, R.C. Genetic and functional analysis of the bovine uterine microbiota. Vet. Microbiol. 2010, 142, 379–385. [Google Scholar]
- Gilbert, R.O.; Shin, S.T.; Guard, C.L.; Erb, H.N.; Frajblat, M. Prevalence of endometritis and its effects on reproductive performance of dairy cows. Theriogenology 2005, 64, 1879–1888. [Google Scholar] [CrossRef]
- Kasimanickam, R.; Cornwell, J.M.; Nebel, R.L. Fertility following postpartum uterine infection in beef cattle. Theriogenology 2004, 61, 1285–1299. [Google Scholar]
- Patterson, D.J.; Perry, R.C.; Kiracofe, G.H.; Bellows, R.A.; Staigmiller, R.B.; Corah, L.R. Management considerations in beef heifer development and puberty. J. Anim. Sci. 2013, 70, 4018–4035. [Google Scholar] [CrossRef]
- Urieli-Shoval, S.; Meek, R.L.; Hanson, R.H.; Eriksen, N.; Benditt, E.P. Human serum amyloid A genes are expressed in monocyte/macrophage cell lines. Am. J. Pathol. 2000, 157, 269–277. [Google Scholar]
- Wagener, K.; Prunner, I.; Pothmann, H. A review of the ongoing discussion about definition, diagnosis and pathomechanisms of bovine endometritis. Theriogenology 2015, 84, 1372–1383. [Google Scholar]
- Obici, L.; Merlini, G. AA amyloidosis: Basic knowledge, unmet needs and future treatments. Swiss Med. Wkly. 2012, 142, w13580. [Google Scholar] [CrossRef]
- Eckersall, P.D.; Bell, R. Acute phase proteins: Biomarkers of infection and inflammation in veterinary medicine. Vet. J. 2010, 185, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Uhlar, C.M.; Whitehead, A.S. Serum amyloid A, the major vertebrate acute-phase reactant. Eur. J. Biochem. 1999, 265, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Meek, R.L.; Benditt, E.P. Amyloid A gene family expression in different mouse tissues. J. Exp. Med. 1986, 164, 2006–2017. [Google Scholar] [CrossRef] [PubMed]
- McDonald, T.L.; Larso, M.A.; Mack, D.R.; Weber, A. Elevated extrahepatic expression and secretion of mammary-associated serum amyloid A 3 (M-SAA3) into colostrum. Vet. Immunol. Immunopathol. 2001, 83, 203–211. [Google Scholar] [CrossRef]
- Reigstad, C.S.; Lundén, G.O.; Felin, J.; Backhed, F. Regulation of serum amyloid A3 (SAA3) in mouse colonic epithelium and adipose tissue by the intestinal microbiota. PLoS ONE 2009, 4, e5842. [Google Scholar] [CrossRef]
- Lin, Y.; Rajala, M.W.; Berger, J.P.; Moller, D.E.; Barzilai, N.; Scherer, P.E. Hyperglycemia-induced production of acute phase reactants in adipose tissue. J. Biol. Chem. 2001, 276, 42077–42083. [Google Scholar] [CrossRef]
- Liu, J.; Wang, D.; Li, S.Q.; Ye, R.D. Suppression of LPS-induced tau heperphosphorylation by serum amyloid A. J. Neuroinflamm. 2016, 13, 28. [Google Scholar] [CrossRef]
- Larson, M.A.; Weber, A.; Weber, A.T.; McDonald, T.L. Differential expression and secretion of bovine serum amyloid A3 (SAA3) by mammary epithelial cells stimulated with prolactin or lipopolysaccharide. Vet. Immunol. Immunopathol. 2005, 107, 255–264. [Google Scholar] [CrossRef]
- Murata, E.; Kozaki, S.; Murakami, T.; Shimizu, K.; Okada, A.; Ishiguro, N.; Inoshima, Y. Differential expression of serum amyloid A1 and A3 in bovine epithelia. J. Vet. Med. Sci. 2020, 82, 1409–1412. [Google Scholar] [CrossRef]
- Saremi, B.; Mielenz, M.; Rahman, M.M.; Hosseini, A.; Kopp, C.; Dänicke, S.; Ceciliani, F. Hepatic and extrahepatic expression of serum amyloid A3 during lactation in dairy cow. J. Dairy Sci. 2013, 96, 6944–6954. [Google Scholar] [CrossRef]
- Green, B.B.; Kerr, D.E. Epigenetic contribution to individual variation in response to lipopolysaccharide in bovine dermal fibroblasts. Vet. Immunol. Immunopathol. 2014, 157, 49–58. [Google Scholar] [CrossRef]
- Taira, Y.; Inoshima, Y.; Ishiguro, N.; Murakami, T.; Matsui, T. Isolation and characterization of monoclonal antibodies against bovine serum amyloid A1 protein. Amyloid 2009, 16, 215–220. [Google Scholar] [CrossRef]
- Kiku, Y.; Nagasawa, Y.; Tanabe, F.; Sugawara, K.; Watanabe, A.; Hata, E.; Ozawa, T.; Nakajima, K.I.; Arai, T.; Hayashi, T. The cell wall component lipoteichoic acid of Staphylococcus aureus induces chemokine gene expression in bovine mammary epithelial cells. J. Vet. Med. Sci. 2016, 78, 1505–1510. [Google Scholar] [CrossRef]
- Chapwanya, A.; Meade, K.G.; Doherty, L.M.; Callanan, J.J.; O’Farrelly, C. Endometrial epithelial cells are potent producers of tracheal antimicrobial peptide and serum amyloid A3 gene expression in response to E. coli stimulation. Vet. Immunol. Immunopathol. 2013, 151, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.W.; Shen, P.X.; Sun, Y.; Zhang, X.; Yan, C.B.; Yu, J.C.; Liu, F.; Yang, D.X.; Xu, E.B.; Wang, Y.Z.; et al. Transcriptome profiling of bovine endometrial epithelial cells induced by lipopolysaccharides in vitro. Anim. Biotechnol. 2023, 34, 4588–4599. [Google Scholar] [CrossRef] [PubMed]
- Parés, S.; Fàbregas, F.; Bach, A.; Garcia-Fruitós, E.; Prado, A.; Arís, A. Short communication: Recombinant mammary serum amyloid A3 as a potential strategy for preventing intramammary infections in dairy cows at dryoff. J. Dairy Sci. 2020, 103, 3615–3621. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. Bacteria within the uterus and therapy of bacterial uterine disease. J. Farm. Anim. Infect. Dis. 2012, 1, 97–103. [Google Scholar]
- Osawa, T. Predisposing factors, diagnostic and therapeutic aspects of persistent endometritis in postpartum cows. J. Reprod. Dev. 2021, 67, 291–299. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aoyagi, K.; Owaki, K.; Sakai, H.; Okada, A.; Inoshima, Y. Serum Amyloid A3 Expression Is Enhanced by Gram-Negative Bacterial Stimuli in Bovine Endometrial Epithelial Cells. Pathogens 2025, 14, 729. https://doi.org/10.3390/pathogens14080729
Aoyagi K, Owaki K, Sakai H, Okada A, Inoshima Y. Serum Amyloid A3 Expression Is Enhanced by Gram-Negative Bacterial Stimuli in Bovine Endometrial Epithelial Cells. Pathogens. 2025; 14(8):729. https://doi.org/10.3390/pathogens14080729
Chicago/Turabian StyleAoyagi, Kazuha, Keishi Owaki, Hiroki Sakai, Ayaka Okada, and Yasuo Inoshima. 2025. "Serum Amyloid A3 Expression Is Enhanced by Gram-Negative Bacterial Stimuli in Bovine Endometrial Epithelial Cells" Pathogens 14, no. 8: 729. https://doi.org/10.3390/pathogens14080729
APA StyleAoyagi, K., Owaki, K., Sakai, H., Okada, A., & Inoshima, Y. (2025). Serum Amyloid A3 Expression Is Enhanced by Gram-Negative Bacterial Stimuli in Bovine Endometrial Epithelial Cells. Pathogens, 14(8), 729. https://doi.org/10.3390/pathogens14080729