

Defining Soilborne Pathogen Complexes Provides a New Foundation for the Effective Management of Faba Bean Root Diseases in Ethiopia
 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the Study Area
2.2. Assessment of Faba Bean Root Rot
2.3. Collection of Soil Samples from Farmer Fields
2.4. Extraction, Identification, and Quantification of DNA from Soils
2.5. High-Throughput Sequencing (HTS) Metabarcoding Studies of Samples
2.6. Bioinformatics and Statistical Analysis
3. Results
3.1. Soil Typology Across the Geographical Zones
3.2. Soilborne Pathogens Detected Using qPCR
3.3. Distribution of Soilborne Pathogens in Relation to Geographical Zones
3.4. Distribution of Soilborne Pathogens and Percentage Severity Index (PSI) Scores in Relation to Soil Type
3.5. Effect of Soil Drainage on Distribution of Some Selected Soilborne Pathogens
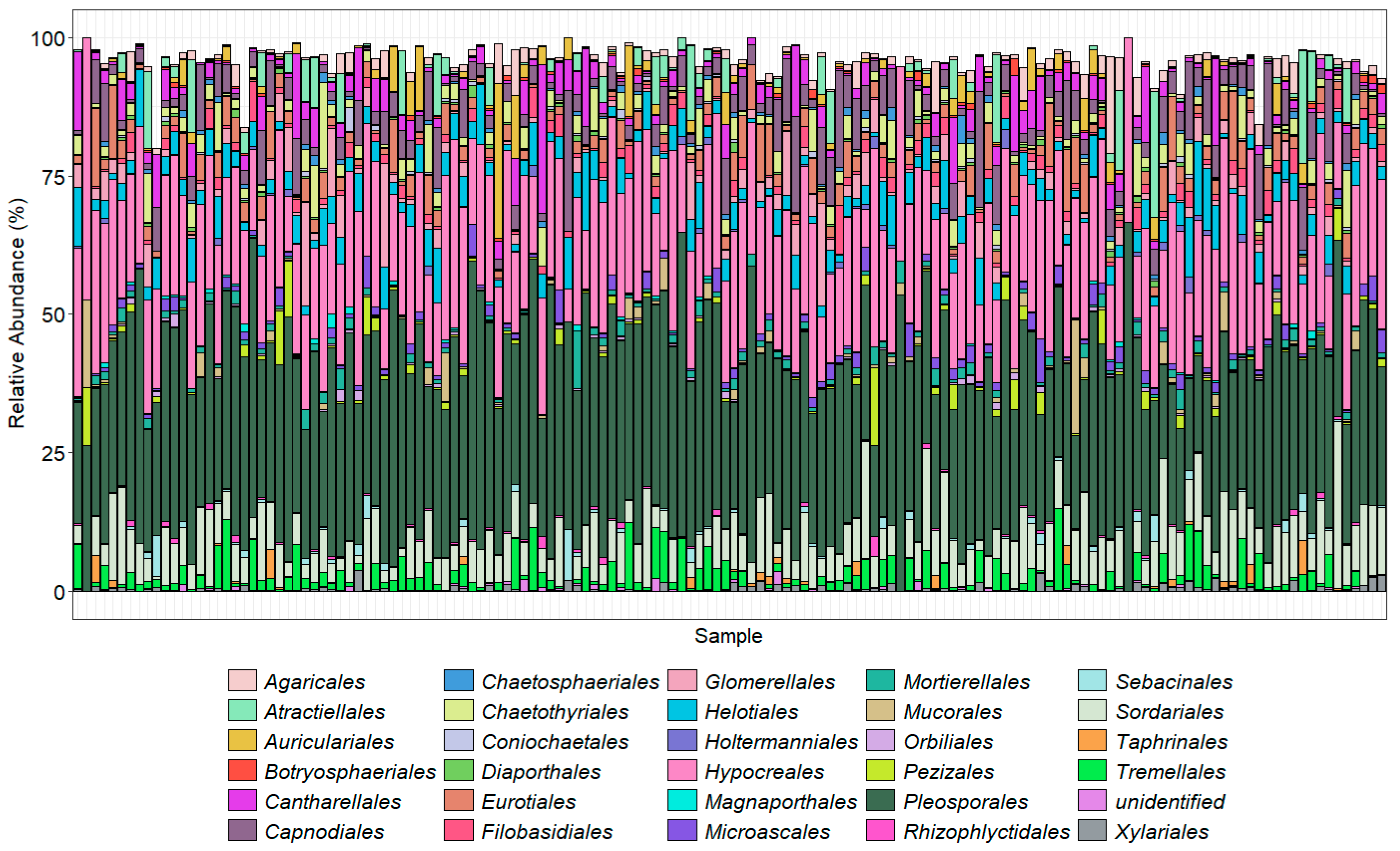
3.6. Metabarcoding of Soilborne Pathogens—Taxonomic Overview
4. Discussion
4.1. Soilborne Pathogens from Survey qPCR Study
4.2. Soilborne Pathogens from Metabarcoding Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PSI | Percentage Severity Index |
| AG | Anastomosis group |
| HTS | High-throughput sequencing |
| qPCR | Quantitative PCR |
| IDM | Integrated disease management |
| ASVs | Amplicon sequence variants |
| SARDI | South Australian Research Development Institute |
| NRRL | ARS culture collection number |
| ARS | Agricultural Research Service |
| NCBI | National Center for Biotechnology Information |
References
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World, 3rd ed.; Oxford University Press: New-York, NY, USA, 2001; p. 316. [Google Scholar] [CrossRef]
- Jensen, E.S.; Peoples, M.B.; Hauggaard-Nielsen, H. Faba bean in cropping systems. Field Crops Res. 2010, 115, 203–216. [Google Scholar] [CrossRef]
- Asfaw, T.; Tesfaye, G.; Beyene, D. Genetics and breeding of faba bean. In Cool Season Food Legumes in Ethiopia, Proceedings of the First National Cool Season Food Legume Review Conference, Aleppo, Syria, 16–20 December 1993; Asfaw, T., Ed.; EIAR, ICARDA: Addis Ababa, Ethiopia, 1994; pp. 122–137. [Google Scholar]
- CSA. Agricultural Sample Survey Area and Production of Major Crops; (Private peasant holdings, ‘Meher’ season); EIAR: Addis Ababa, Ethiopia, 2021; Volume I, pp. 135–143. [Google Scholar]
- Crepona, K.; Marget, P.; Peyronnet, C.; Carrouéea, B.; Arese, P.; Duc, G. Nutritional value of faba bean (Vicia faba L.) seeds for feed and food. Field Crops Res. 2010, 115, 329–339. [Google Scholar] [CrossRef]
- Mussa, J.; Dereje, G.; Gemechu, K. Procedures of Faba Bean Improvement Through Hybridization; Technical Manual, No. 21; Ethiopian Institute of Agricultural Research (EIAR): Addis Ababa, Ethiopia, 2008; p. 48. [Google Scholar]
- Marzano, S.Y.L. Assessment of Disease Suppression in Organic Transitional Cropping Systems. Ph.D. Thesis, The University of Illinois, Champaign, IL, USA, 2012; pp. 151–165. [Google Scholar]
- Stewart, R.B.; Dagnachew, Y. Index of Plant Disease in Ethiopia; Experimental Station Bulletin, No. 30; College of Agriculture, Haile Sellassie I University: Addis Ababa, Ethiopia, 1967; p. 35. [Google Scholar]
- Abraham, T. Increasing Crop Production Through Improved Plant Protection—Volume I; Plant Protection Society of Ethiopia (PPSE): Addis Ababa, Ethiopia, 2008; p. 598. [Google Scholar]
- Berhanu, B.; Getachew, M.; Teshome, G.; Temesgen, B. Faba bean and field pea diseases research. In Food and Forage Legumes: Progress and Prospects, Proceedings of the Progress of the Workshop on Food and Forage Legumes, 22–26 September 2003, Aleppo, Syria; EIAR, ICARDA: Addis Ababa, Ethiopia, 2003; pp. 221–227. [Google Scholar]
- Eshetu, B.; Amare, A.; Seid, A. Evaluation of local isolates of Trichoderma spp. against black root rot (Fusarium solani) on faba bean. Plant Pathol. Microbiol. 2015, 6, 6279. [Google Scholar]
- Rubiales, D.; Khazaei, H. Advances in disease and pest resistance in faba bean. Theor. Appl. Genet. 2022, 135, 3735–3756. [Google Scholar] [CrossRef] [PubMed]
- Mukankusi, C.M.; Melis, R.J.; Derera, J.R.; Buruchara, R.A.; Mark, D. A screening technique for resistance to Fusarium root rot of common bean. Afr. J. Plant Sci. 2011, 5, 152–161. [Google Scholar]
- Bogale, M.; Steenkamp, E.T.; Wingfield, M.J.; Wingfield, B.D. Diverse Fusarium solani isolates colonies agricultural environments in Ethiopia. Eur. J. Plant Pathol. 2009, 124, 369–378. [Google Scholar] [CrossRef]
- Belay, H.; Anteneh, B. Integrated management of faba bean black root rot (Fusarium solani) through varietal resistance, drainage and adjustment of planting time. J. Plant Pathol. Microbiol. 2016, 7, 7. [Google Scholar] [CrossRef]
- Pandey, A.K.; Barbetti, M.J.; Kumar, A.; Gaulin, E.; Le May, C.; Pilet-Nayel, M.L.; You, M.P.; Lamichhane, J.R. Root disease complexes of arable crops: Where do we stand and where should we go? Crit. Rev. Plant Sci. 2025, 44, 1–29. [Google Scholar] [CrossRef]
- You, M.P.; Barbetti, M.J. Manipulating ecosystem environment enables management of soilborne pathogen complexes in annual legume forage systems. Plant Pathol. 2019, 68, 454–469. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; You, M.P.; Laudinot, V.; Byamukama, E.; Barbetti, M.J.; Aubertot, J.N. Revisiting sustainability of fungicide seed treatments for field crops with a focus on Franco-Australian-North American context. Plant Dis. 2020, 104, 610–623. [Google Scholar] [CrossRef]
- You, M.P.; Lamichhane, J.R.; Aubertot, J.N.; Barbetti, M.J. Understanding why effective fungicides against individual soilborne pathogens are ineffective with soilborne pathogen complexes. Plant Dis. 2020, 104, 904–920. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; You, M.P.; Barbetti, M.J.; Aubertot, J.N. Crop Establishment SIMulator: A qualitative aggregative model to predict the role of phytobiomes on field crop establishment. Phytobiomes J. 2020, 4, 327–339. [Google Scholar] [CrossRef]
- Ophel-Keller, K.; McKay, A.; Hartley, D.; Herdina; Curran, J. Development of a routine DNA-based testing service for soilborne diseases in Australia. Australas. Plant Pathol. 2008, 37, 243–253. [Google Scholar] [CrossRef]
- Vetrovsky, T.; Morais, D.; Kohout, P.L. Global Fungi, a global database of fungal occurrences from high-throughput-sequencing metabarcoding studies. Sci. Data 2020, 7, 228. [Google Scholar] [CrossRef]
- Davis, D.W.; Fritz, V.A.; Pfleger, F.L.; Percich, J.A.; Malvick, D.K. Garden pea lines resistant to root rot caused by Aphanomyces euteiches Drechs. Hort Sci. 1995, 30, 639–640. [Google Scholar]
- Huising, E.; Jeroen; David, E.; Bignell; Fatima, M.S.M. Chapter Two—Sampling strategy and Desing to Evaluate Below-ground Biodiversity. In A Handbook of Tropical Soil Biology: Sampling and Characterization of Below-Ground Biodiversity; Springer: London, UK, 2008; pp. 30–31. [Google Scholar]
- Tenedero, R.A.; Surtida, M.B. Soil Sampling and Preparation for Laboratory Analysis; Aquaculture Technology Module (5) Tigbauan, Iloilo; SEAFDEC Aquaculture Department: Tigbauan, Philippines, 1986; p. 11. [Google Scholar]
- Simpson, R.J.; Richardson, A.E.; Rile, I.T.; McKay, A.C.; McKay, S.F.; Ballard, R.A.; Ophel-Keller, K.; Hartley, D.; O’Rourke, T.A.; Sivasithamparam, K.; et al. Damage to roots of Trifolium subterraneum L., failure of seedlings to establish and the presence of root pathogens during autumn–winter. Grass Forage Sci. 2011, 66, 585–605. [Google Scholar] [CrossRef]
- Vancov, T.; Keen, B. Amplification of soil fungal community DNA using the ITS86F and ITS4 primers. FEMS Microbiol. Lett. 2009, 296, 91–96. [Google Scholar] [CrossRef]
- White, T.J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Bachofer, M. Molecular Biological Population Studies on Plasmopara halstedii, the Downy Mildew of Sunflowers. Ph.D. Thesis, University of Hohenheim, Stuttgart, Germany, 2004; pp. 1–140. [Google Scholar]
- Riit, T.; Tedersoo, L.; Drenkhan, R.; Runno-Paurson, E.; Kokko, H.; Anslan, S. Oomycete-specific ITS primers for identification and metabarcoding. Myco Keys. 2016, 14, 17–30. [Google Scholar] [CrossRef]
- Giblot-Ducray, D. New Capability to Survey Pulse and Cereal Crops for Root Pathogens; Report, No.: GRDC UOA1907-004BLX; South Australian Research and Development Institute: Urrbrae, Australia, 2020. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O. Scikit-learn. machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. Peer J. 2016, 4, 2584. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.M.; Proctor, D.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. bioRxiv 2017. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Hadley, W. Ggplot2; Springer Science & Business Media, LLC: New York, NY, USA, 2016; p. 213. [Google Scholar]
- Torres-Cruz, T.J.; Whitaker, B.K.; Proctor, R.H.; Broders, K.; Laraba, I.; Kim, H.-S.; Brown, D.W.; O’Donnell, K.; Estrada-Rodríguez, T.L.; Lee, W.-H.; et al. FUSARIUM-ID v.3.0: An Updated, Downloadable Resource for Fusarium Species Identification. Plant Dis. 2022, 106, 1610–1616. [Google Scholar] [CrossRef]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Põlme, S.; Riit, T.; Liiv, I. Shotgun metagenomes and multiple primer pair-barcode combinations of amplicons reveal biases in metabarcoding analyses of fungi. MycoKeys 2015, 10, 1–43. [Google Scholar] [CrossRef]
- Eshetu, B.; Amare, A.; Seid, A. Associations of biophysical factors with faba bean black root rot (Fusarium solani) epidemics in the northeastern highlands of Ethiopia. Crop Prot. 2013, 52, 39–46. [Google Scholar] [CrossRef]
- Lievens, B.; Brouwer, M.; Vanachter, A.C.R.C.; Cammue, B.P.A.; Thomma, B.P.H.J. Real-time PCR for detection and quantification of fungal and oomycete tomato pathogens in plant and soil samples. Plant Sci. 2006, 171, 155–165. [Google Scholar] [CrossRef]
- Carling, D.E.; Kuninaga, S.; Brainard, K.A. Hyphal anastomosis reactions, rDNA-internal transcribed spacer sequences, and virulence levels among subsets of Rhizoctonia solani anastomosis group-2 (AG-2) and AG-BI. Phytopathology 2002, 92, 4350. [Google Scholar] [CrossRef]
- Hane, J.K.; Anderson, J.P.; Williams, A.H.; Sperschneider, J.; Singh, K.B. Genome sequencing and comparative genomics of the broad host-range pathogen Rhizoctonia solani AG8. PLoS Genet. 2014, 10, e1004281. [Google Scholar] [CrossRef]
- Samuel, A.W.; Rama, D.; Narla-Eunice, W.M.; James, W.M. Prevalence and Characterization of Bean Root Rot Pathogens in Different Agro Ecological Zones. Ph.D. Thesis, Western Kenya University of Nairobi, Nairobi, Kenya, 2019. [Google Scholar] [CrossRef]
- Agnieszka, P.; Adam, O.; Gabriel, F.N.; Andrzej, K.; Marcin, K.; Grzegorz, D. Effect of weather conditions on yield and health status of faba bean seeds in Poland. Agronomy 2020, 10, 48. [Google Scholar] [CrossRef]
- Mwang’ombe, A.W.; Thiongo, G.; Olubayo, F.M.; Kiprop, E.K. Occurrence of root rot disease of common bean (Phaseolus vulgaris L.). Association with bean stem maggot Ophyiomia sp. in Embu District, Kenya. Plant Pathol. J. 2007, 6, 141–146. [Google Scholar] [CrossRef]
- Naseri, B. Bean production and Fusarium root rot in diverse soil environments in Iran. J. Soil Sc. Plant Nutrition. 2014, 14, 177–188. [Google Scholar] [CrossRef]
- Sun, R.Y.; Liu, Z.C.; Fu, K.; Fan, L.; Chen, J. Trichoderma biodiversity in China. J. Appl. Genet. 2012, 53, 343–354. [Google Scholar] [CrossRef]
- Mukhongo, R.W.; Kavoo-Mwangi, M.A.; Kahangi, M.E.; Ateka, E.M.; Were, A.B.; Okalebo, J.R.; Mutegi, M.E.; Mwangi, K.E.; Tepeni, T.T.; Njuguini, K.S.; et al. Occurrence of arbuscular mycorrhizal fungi and Fusarium in TC banana rhizosphere inoculated with microbiological products in different soils in Kenya. Int. J. Soil Sci. 2015, 10, 45–62. [Google Scholar] [CrossRef]
- Coyne, M. Soil Microbiology: An Explanatory Approach; Delmar Publishers: Albany, NY, USA, 1999. [Google Scholar]
- Hillel, D. Environmental Soil Physics; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Barnes, J.G.; Niall, P.M.; Rebecca, R.; Gary, D. Bending Extreme rainfall affects assembly of the root-associated fungal community. J. Phytologist. 2018, 220, 1172–1184. [Google Scholar] [CrossRef]
- Gregorich, E.G.; Beare, M.H.; McKim, U.F.; Skjemstad, J.D. Chemical and biological characteristics of physically uncomplexed organic matter. Soil Sci. Soc. Am. J. 2006, 70, 975–985. [Google Scholar] [CrossRef]
- Siles, J.A.; Margesin, R. Abundance and diversity of bacterial, archaeal, and fungal communities along an altitudinal gradient in alpine forest soils: What are the driving factors? Microb. Ecol. 2016, 72, 207–220. [Google Scholar] [CrossRef]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Mazzola, M.; Hewavitharana, S.S.; Strauss, S.L. Brassica seed meal soil amendments transform the rhizosphere microbiome and improve apple production through resistance to pathogen reinfestation. Ecol. Epidemiol. 2015, 105, 460–469. [Google Scholar] [CrossRef]
- Gill, J.S.; Sivasithamparam, K.; Smettem, K.R.J. Soil types with different textures affect development of Rhizoctonia root rot of wheat seedlings. Plant Soil. 2000, 221, 113–120. [Google Scholar] [CrossRef]
- Yaquelin, N.; Sarah, V.B.; Soraya, C.F.; Alexander, J.; Rene, C.; Lidcay, H.; Monica, H. Influence of soil type and indigenous pathogenic fungi on bean hypocotyl rot caused by Rhizoctonia solani AG4 HGI in Cuba. Soil Biol. Biochem. 2010, 42, 797–803. [Google Scholar] [CrossRef]
- Bue, M.; Reich, M.; Murat, C.; Morin, E.; Nilsson, R.H.; Uroz, S. Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol. 2009, 184, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.K.; Jensen, J.D.; Nielsen, G.C.; Jensen, J.E.; Spliid, N.H.; Thomsen, I.K. Fusarium head blight of cereals in Denmark: Species complex and related mycotoxins. Phytopathology 2011, 101, 960–969. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Tacke, B.K.; Casper, H.H. Gene genealogies reveal global phylogeographic structure and reproductive isolation among lineages of Fusarium graminearum, the fungus causing wheat scab. Proc. Natl. Acad. Sci. USA 2000, 97, 7905–7910. [Google Scholar] [CrossRef] [PubMed]
- Geiser, D.M.; Gasco, J.; Kang, M.M.; Mkalowska, S.; Veeraraghavan, I.; Ward, N. Fusarium: A DNA sequence database for identifying Fusarium. Eur. J. Plant Pathol. 2004, 110, 473–479. [Google Scholar] [CrossRef]
- Alford, B.A.; Bekele, D.; Yimer, S.M.; Fayyaz, A.; Carrasquilla-Garcia, N.; Chang, P.L.; Badger, C.; Surendrarao, A.; von Wettberg, E.J.B.; Munis, F.H.; et al. Microbial community analysis offers insight into the complex origins of plant disease in a smallholder farm context. Phytobiomes J. 2024, 8, 456–468. [Google Scholar] [CrossRef]
- Nicolaisen, M.; Suproniene, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Method. 2009, 76, 234–240. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Zone | Altitude (m) | Annual Rainfall (mm) | Average Annual Temperature Range (°C) | Root Zone Moisture (%) | Soil Surface Moisture (%) | Crop Type | ||
|---|---|---|---|---|---|---|---|---|
| Faba Bean | Field Pea | Lentil | ||||||
| South Wollo | 2605–2793 | 889–939 | 8.7–21.9 | 57–88 | 52–85 | 18 | - | - |
| North Shoa (AM) | 2462–3210 | 772–927 | 9.2–21.8 | 62–89 | 59–87 | 15 | 6 | - |
| North Shoa (OR) | 2510–3097 | 772–1038 | 7.9–19.0 | 64–91 | 60–88 | 20 | 1 | - |
| West Shoa | 2135–2699 | 847–1123 | 10.5–23.5 | 69–94 | 67–90 | 19 | 4 | - |
| Arsi | 2366–2976 | 587–799 | 8.1–21.2 | 52–77 | 50–73 | 20 | - | - |
| West Arsi | 2215–2840 | 770–851 | 9.6–21.5 | 55–85 | 53–82 | 18 | - | - |
| Bale | 2359–3334 | 666–858 | 8.78–21.4 | 50–80 | 49–76 | 16 | 7 | 6 |
| Pathogen | * Ethiopian New Record | Unit | Faba Bean (126 Sites) | Pea (18 Sites) | Lentil (6 Sites) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Incidence (No./126) | Ave DNA | Highest DNA | Incidence (No./18) | Ave DNA | Highest DNA | Incidence (No./6) | Ave DNA | Highest DNA | |||
| Root pathogens | |||||||||||
| Aphanomyces euteiches | * | pg DNA/g sample | 44 | 6 | 165 | 5 | 10 | 140 | 2 | 19 | 102 |
| Fusarium redolens | * | Ct value | 66 | 35 | 44 | 8 | 34 | 39 | 3 | 35 | 37 |
| Macrophomina phaseolina | * | kDNA copies/g sample | 123 | 47 | 316 | 16 | 39 | 112 | 6 | 70 | 224 |
| Phytophthora medicaginis | * | kDNA copies/g sample | 1 | 0.002 | 0.21 | 0 | 0 | 0 | 0 | 0 | 0 |
| Phytophthora megasperma | * | kDNA copies/g sample | 36 | 0.6 | 6.13 | 5 | 2 | 16 | 2 | 1 | 3 |
| Pythium clade F | pg DNA/g sample | 108 | 61 | 417 | 13 | 38 | 121 | 4 | 52 | 157 | |
| Pythium clade I | pg DNA/g sample | 124 | 135 | 1651 | 18 | 115 | 656 | 6 | 120 | 293 | |
| S. sclerotiorum/S. minor | kDNA copies/g sample | 32 | 83 | 2562 | 2 | 76 | 1360 | 1 | 3 | 21 | |
| Thielaviopsis basicola | * | kDNA copies/g sample | 22 | 2.3 | 72 | 1 | 1 | 13 | 0 | 0 | 0 |
| Verticillium dahliae | pg DNA/g sample | 27 | 0.9 | 41 | 2 | 0 | 1 | 2 | 1 | 3 | |
| Rhizoctonia solani AG2.1 | * | pg DNA/g sample | 63 | 32 | 1039 | 11 | 32 | 225 | 4 | 10 | 34 |
| Rhizoctonia solani AG2.2 | * | pg DNA/g sample | 44 | 240 | 18,390 | 5 | 227 | 2489 | 1 | 11 | 67 |
| Rhizoctonia solani AG3 | * | pg DNA/g sample | 16 | 18 | 728 | 3 | 2 | 31 | 1 | 0 | 2 |
| Rhizoctonia solani AG4 | * | pg DNA/g sample | 39 | 119 | 4221 | 7 | 1015 | 15,244 | 3 | 37 | 165 |
| Rhizoctonia solani AG5 | * | pg DNA/g sample | 6 | 18 | 1971 | 0 | 0 | 0 | 0 | 0 | 0 |
| Leaf pathogens | |||||||||||
| Ascochyta fabae | kDNA copies/g sample | 53 | 176 | 12,143 | 5 | 31 | 349 | 3 | 233 | 856 | |
| Ascochyta lentis | kDNA copies/g sample | 2 | 0.05 | 6.08 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Botrytis cinerea | kDNA copies/g sample | 100 | 14 | 440 | 13 | 2 | 13 | 4 | 4 | 20 | |
| Botrytis fabae | kDNA copies/g sample | 120 | 330 | 4500 | 18 | 206 | 1663 | 6 | 75 | 316 | |
| Didymella pinodes/Phoma pinodella | pg DNA/g sample | 110 | 1051 | 19,256 | 16 | 1151 | 15,162 | 5 | 370 | 1450 | |
| Phoma rabiei | kDNA copies/g sample | 2 | 0.01 | 0.61 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Nematodes | |||||||||||
| Meloidogyne javanica/incognita/arenaria | pg DNA/g sample | 6 | 23 | 1791 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Pratylenchus neglectus | nematodes/g soil | 4 | 0.04 | 4.06 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Pratylenchus penetrans | nematodes/g soil | 4 | 0.06 | 3.34 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Pratylenchus quasitereoides | nematodes/g soil | 1 | 0 | 0.02 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Pratylenchus thornei | nematodes/g soil | 75 | 1 | 31 | 13 | 6 | 93 | 5 | 4 | 14 | |
| PSI | No. | A. euteiches | M. phaseolina | Ph. megasperma | Pythium clade F | Pythium clade I | T. basicola | V. dahliae | ||||||||||||||
| Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | ||
| 1–5 | 45 | 6 | 0 | 165 | 38 | 0 | 169 | 0.5 | 0 | 4 | 57 | 0 | 368 | 93 | 0 | 587 | 0.6 | 0 | 11 | 0 | 0 | 3 |
| 6–15 | 35 | 4 | 0 | 49 | 57 | 1 | 316 | 0.9 | 0 | 6 | 63 | 0 | 319 | 109 | 0 | 770 | 3.4 | 0 | 72 | 0 | 0 | 4 |
| 16–25 | 22 | 9 | 0 | 87 | 63 | 0 | 216 | 0.3 | 0 | 2 | 50 | 0 | 143 | 149 | 2 | 810 | 4.5 | 0 | 69 | 1 | 0 | 7 |
| 26–50 | 17 | 7 | 0 | 37 | 38 | 2 | 110 | 0.7 | 0 | 6 | 43 | 0 | 290 | 296 | 5 | 1651 | 0.6 | 0 | 8 | 1 | 0 | 10 |
| >50 | 6 | 10 | 0 | 37 | 25 | 2 | 100 | 0.3 | 0 | 1 | 173 | 0 | 417 | 90 | 10 | 201 | 5.9 | 0 | 33 | 5 | 0 | 41 |
| R. solani AG2-1 | R. solani AG2-2 | R. solani AG3 | R. solani AG4 | R. solani AG5 | A. fabae | B. cinerea | ||||||||||||||||
| PSI | No. | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max |
| 1–5 | 45 | 20 | 0 | 225 | 175 | 0 | 3095 | 6 | 0 | 226 | 118 | 0 | 4017 | 38 | 0 | 1971 | 69 | 0 | 881 | 7 | 0 | 104 |
| 6–15 | 35 | 22 | 0 | 143 | 101 | 0 | 1343 | 55 | 0 | 728 | 325 | 0 | 4221 | 3 | 0 | 78 | 10 | 0 | 225 | 41 | 0 | 440 |
| 16–25 | 22 | 50 | 0 | 1039 | 563 | 0 | 18,390 | 3 | 0 | 87 | 473 | 0 | 15,244 | 6 | 0 | 2226 | 358 | 0 | 12,143 | 4 | 0 | 40 |
| 26–50 | 17 | 38 | 0 | 697 | 9 | 0 | 143 | 9 | 0 | 101 | 3 | 0 | 39 | 0 | 0 | 0 | 289 | 0 | 4355 | 6 | 0 | 63 |
| >50 | 6 | 22 | 0 | 127 | 8 | 0 | 67 | 13 | 0 | 100 | 19 | 0 | 165 | 0 | 0 | 0 | 61 | 0 | 518 | 5 | 0 | 21 |
| P. rabiei | M. javanica++ | P. neglectus | P. thornei | B. fabae | D. pinodes/pinodella | P. penetrans | ||||||||||||||||
| PSI | No. | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max | Ave | Min | Max |
| 1–5 | 45 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 4 | 0 | 0 | 5 | 259 | 0 | 3831 | 3010 | 0 | 19,256 | 0 | 0 | 0 |
| 6–15 | 35 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 31 | 416 | 0 | 4500 | 1457 | 0 | 16,631 | 0 | 0 | 3 |
| 16–25 | 22 | 0 | 0 | 0 | 0 | 0 | 396 | 0 | 0 | 1 | 3 | 0 | 93 | 255 | 0 | 4116 | 1103 | 0 | 15,162 | 0 | 0 | 0 |
| 26–50 | 17 | 0 | 0 | 0 | 0 | 0 | 1791 | 0 | 0 | 0 | 1 | 0 | 14 | 145 | 0 | 754 | 744 | 0 | 6884 | 0 | 0 | 0 |
| >50 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 9 | 859 | 0 | 3739 | 223 | 0 | 675 | 0 | 0 | 3 |
| Source Variation | Significances in Relation to | |||||||
|---|---|---|---|---|---|---|---|---|
| pH | PSI | A. euteiches | F. redolens | M. phaseolina | Ph. megasperma | Pythium clade F | Pythium clade I. | |
| Soil type | *** | *** | *** | * | ns | ** | ns | * |
| Zone | *** | ** | ns | ns | ns | * | ns | ** |
| Soil type x Zone | *** | ns | ns | ns | ns | ns | ns | *** |
| S. sclerotiorum/minor. | T. basicola | V. dahliae | R. solani AG2.1 | B. fabae | R. solani AG2.2 | R. solani AG3 | R. solani AG4 | |
| Soil type | *** | ** | ns | ns | ns | ** | ns | ns |
| Zone | ns | ns | *** | ns | ** | ns | ** | ns |
| Soil type x Zone | ns | ** | ns | ns | ** | ns | ns | ns |
| R. solani AG5 | D. pinodes+ | P. rabiei | M. javanica++ | P. neglectus | P. penetrans | P. quasitereoides | P. thornei | |
| Soil type | ns | ** | ns | *** | ns | ns | ns | ns |
| Zone | ns | * | ns | ns | ns | ns | ns | * |
| Soil type x Zone | ns | ns | ns | ns | ns | ns | ns | ns |
| Source of Variation | pH | PSI | Pathogens | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Geographical Zone | A. eut | F. red. | M. ph | P. meg | Py. F | Py. I | S. scl | T. bas | V. dha | R.A2.1 | R.A2.2 | ||
| Arsi | 4.9 bc | 10.7 bc | 2.8 ab | 19.8 a | 82.5 a | 1.1 a | 48.8 ab | 93.1 bc | 71.5 a | 0.5 c | 0.1 b | 42.5 a | 83.9 a |
| Bale | 5.7 ab | 5.1 c | 0.6 b | 10.9 ab | 90.9 a | 0.6 ab | 37.9 ab | 64.6 bc | 0 a | 0.4 c | 0 b | 8.0 a | 180.3 a |
| North Shoa Amhara | 5.9 ab | 14.4 ab | 6.2 ab | 17.0 a | 35.5 ab | 0.5 ab | 51.9 ab | 66.0 bc | 32.9 a | 0.04 c | 0.8 b | 3.1 a | 42.3 a |
| North Shoa Oromia | 4.2 cd | 10.4 bc | 3.0 ab | 13.3 ab | 39.2 ab | 0.3 b | 40.5 ab | 88.5 bc | 118.2 a | 6.2 a | 0.2 b | 25.1 a | 119.7 a |
| South Wollo | 3.5 d | 5.7 c | 3.4 ab | 4.9 b | 19.9 b | 0.2 b | 22.5 b | 30.5 c | 95.7 a | 0.04 c | 0.4 b | 7.5 a | 167.1 a |
| West Arsi | 4.2 cd | 15.0 ab | 9.9 a | 15.4 ab | 15.0 b | 0.2 b | 62.2 a | 111.0 b | 16.6 a | 3.0 b | 0.6 b | 15.8 a | 119.0 a |
| West Shoa | 6.4 a | 20.5 a | 9.2 a | 19.8 a | 48.1 ab | 1.0 a | 46.2 ab | 207.0 a | 101.7 a | 2.0 bc | 3.7 a | 48.6 a | 16.6 a |
| LSD | 1.12 | 8.08 | 7.22 | 11.04 | 56.95 | 0.63 | 36.60 | 78.34 | 123.15 | 2.41 | 1.66 | 52.42 | 208.13 |
| Geographical Zone | R.AG3 | R.AG4 | R.AG5 | A. fab | B. cin | B. fab | D. pin | P. rab | M. jav | P. neg | P. pen | P. qua | P. tho |
| Arsi | 19.9 abc | 39.3 a | 0 a | 2.3 b | 2.6 b | 126.9 c | 277.6 c | 0 a | 0 b | 0.01 a | 0.09 a | 0 a | 2.3 a |
| Bale | 12.9 bcd | 47.3 a | 0 a | 6.7 b | 3.1 b | 118.1 c | 430.7 bc | 0 a | 0.1 b | 0 a | 0 a | 0 a | 0.11 b |
| North Shoa Amhara | 6.9 cd | 7.4 a | 0 a | 454.6 a | 9.7 ab | 108.3 c | 1142.3 ab | 0 a | 0.4 b | 0.02 a | 0 a | 0 a | 0.16 b |
| North Shoa Oromia | 38.1 a | 104.4 a | 8.5 a | 221.1 ab | 13.4 ab | 247.3 bc | 1166.9 ab | 0 a | 0 b | 0 a | 0.01 a | 0 a | 0.5 b |
| South Wollo | 0.1 d | 151.4 a | 0 a | 6.1 b | 4.9 ab | 405.4 ab | 392.9 bc | 0 a | 0 b | 0.15 a | 0 a | 0.1 a | 0.17 b |
| West Arsi | 30.7 ab | 164.2 a | 3.0 a | 8.8 b | 11.3 ab | 386.6 ab | 294.8 c | 0 a | 0.1 b | 0.01 a | 0 a | 0 a | 0.24 b |
| West Shoa | 17.5 bcd | 8.3 a | 0.5 a | 62.2 ab | 25.2 a | 459.3 a | 1307.3 a | 0.01 a | 165.0 a | 0 a | 0.13 a | 0 a | 1.3 ab |
| LSD | 19.69 | 193.12 | 9.02 | 402.92 | 22.10 | 202.87 | 845.12 | 0.01 | 61.93 | 0.16 | 0.16 | 0.00 | 1.44 |
| Soil Type | pH | PSI | A. eut | F. red | M. pha | P. meg | Py. F | Py. I | S. scl | T. bas | V. dha | R.AG2.1 | R.AG2.2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mixed | 5.1 b | 10.0 b | 2.8 b | 11.1 b | 32.9 a | 0.4 b | 28.4 b | 63.4 b | 18.7 b | 2.1 a | 0.2 b | 21.9 a | 33.9 b |
| Nitisol | 3.3 c | 6.5 b | 0.9 b | 11.8 b | 51.6 a | 0.3 b | 18.8 b | 83.8 b | 11.9 b | 0.1 b | 0.7 ab | 7.6 a | 45.9 b |
| Vertisol | 6.5 a | 18.5 a | 11.3 a | 20.5 a | 57.4 a | 0.9 a | 85.7 a | 135.9 a | 156.5 a | 3.0 a | 1.5 a | 35.1 a | 232.6 a |
| LSD | 0.7 | 5.3 | 4.7 | 7.2 | 37.3 | 0.4 | 24.0 | 51.3 | 80.6 | 1.6 | 1.1 | 34.3 | 136.2 |
| R.AG3 | R.AG4 | R.AG5 | A. fab | B. cin | B. fab | D. pin | P. rab | M. jav | P. neg | P. pen | P. qua | P. tho | |
| Mixed | 4.2 b | 9.0 b | 0 a | 11.4 a | 6.1 a | 181.0 b | 556.6 b | 0 a | 0 b | 0.01 a | 0.05 a | 0 a | 0.5 ab |
| Nitisol | 9.8 b | 22.7 b | 0.2 a | 50.0 a | 6.7 a | 143.5 b | 323.6 b | 0 a | 0 b | 0 a | 0 a | 0 a | 0.2 b |
| Vertisol | 40.1 a | 192.1 a | 4.9 a | 265.2 a | 17.2 a | 469.3 a | 1268.0 a | 0 a | 70.9 a | 0.07 a | 0.05 a | 0 a | 1.3 a |
| LSD | 12.9 | 126.4 | 5.9 | 263.8 | 14.5 | 132.8 | 553.3 | ns | 40.5 | 0.11 | 0.10 | 0.00 | 0.95 |
| Vertisol Soils | Other Soil Types | All | |||||
|---|---|---|---|---|---|---|---|
| No Drainage | Drainage | Overall | No Drainage | Drainage | Overall | ||
| Number of sites | 22 | 24 | 46 | 58 | 21 | 79 | 125 |
| Root rot incidence (%) | 25.8 | 10.7 | 17.9 | 15.6 | 14.7 | 15.3 | 16.3 |
| Average readings | |||||||
| Aphanomyces euteiches | 4.4 | 6.1 | 5.3 | 8.6 | 2.5 | 7.0 | 6.4 |
| Macrophomina phaseolina | 32.8 | 46.2 | 39.8 | 53.7 | 44.8 | 51.4 | 47.1 |
| Pythium clade F | 90.5 | 50.8 | 69.8 | 57.5 | 51.6 | 56.0 | 61.0 |
| Pythium clade I | 166.7 | 69.3 | 115.9 | 171.0 | 75.8 | 145.7 | 134.7 |
| Rhizoctonia solani AG2.1 | 19.1 | 28.3 | 23.9 | 44.6 | 14.8 | 36.7 | 32.0 |
| Rhizoctonia solani AG2.2 | 162.3 | 44.0 | 100.6 | 405.2 | 86.5 | 320.5 | 239.6 |
| Rhizoctonia solani AG3 | 7.1 | 46.9 | 27.9 | 14.9 | 4.3 | 12.1 | 17.9 |
| Rhizoctonia solani AG4 | 254.6 | 118.1 | 183.4 | 91.6 | 56.4 | 82.3 | 119.5 |
| Rhizoctonia solani AG5 | 0.0 | 9.6 | 5.0 | 1.6 | 93.9 | 26.1 | 18.4 |
| Isolate ID | Species Complex | Species Name |
|---|---|---|
| NRRL_22278 | F. solani Species Complex (FSSC) 11a | F. vanettenii |
| NRRL_22436 | FSSC 8b | F. neocosmosporiellum |
| NRRL_32484/NRRL_32810/NRRL_32492 | FSSC 5j, m, and c | F. solani |
| NRRL_22640 | FSSC 2i | F. keratoplasticum |
| NRRL_25221 | FFSC Asian | Fusarium sp. (undescribed) |
| NRRL_22901 | F. redolens Species Complex (FRSC) | F. redolens |
| CBS_143611 | F. tricinctum species complex (FTSC) 6 | F. iranicum |
| F1529 | FTSC 4 | F. avenaceum |
| P454a | FTSC 2 | F. acuminatum |
| FRC_R8227 | F. sambucinum species complex (FSAMSC) | F. sambucinum |
| NRRL_28505/NRRL_29011/NRRL_26083 | FSAMSC | F. graminearum |
| NRRL_66931 | FSAMSC | F. brachygibbosum |
| NRRL_46743 | FSAMSC | Fusarium sp. (undescribed) |
| NRRL_13338/NRRL_28505 | F. chlamydosporum species complex (FCSC) 4a and b | F. nelsonii |
| NRRL_45997/NRRL_43623 | F. incarnatum-equiseti species complex (FIESC) 5 a,b,e, and f | F. clavum |
| NRRL_36269 | FIESC 12a, b, and c | F. flagelliforme |
| NRRL_43638 | FIESC 6a | F. brevicaudatum |
| NRRL_36321/NRRL43636 | FIESC 14a and c | F. equiseti |
| NRRL_22550/NRRL_22544/NRRL_36471/NRRL_26437/NRRL_20433/NRRL_36286/NRRL_26438/NRRL_28366/NRRL_22534/NRRL_26033 | F. oxysporum species complex (FOSC) | F. oxysporum |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yilma, S.; Bekele, B.; Van Leur, J.; You, M.P.; Kemal, S.-A.; Giblot-Ducray, D.; Hill, K.; Selvaraji, T.; Lencho, A.; Driba, L.; et al. Defining Soilborne Pathogen Complexes Provides a New Foundation for the Effective Management of Faba Bean Root Diseases in Ethiopia. Pathogens 2025, 14, 695. https://doi.org/10.3390/pathogens14070695
Yilma S, Bekele B, Van Leur J, You MP, Kemal S-A, Giblot-Ducray D, Hill K, Selvaraji T, Lencho A, Driba L, et al. Defining Soilborne Pathogen Complexes Provides a New Foundation for the Effective Management of Faba Bean Root Diseases in Ethiopia. Pathogens. 2025; 14(7):695. https://doi.org/10.3390/pathogens14070695
Chicago/Turabian StyleYilma, Solomon, Berhanu Bekele, Joop Van Leur, Ming Pei You, Seid-Ahmed Kemal, Danièle Giblot-Ducray, Kelly Hill, Thangavel Selvaraji, Alemu Lencho, Lemma Driba, and et al. 2025. "Defining Soilborne Pathogen Complexes Provides a New Foundation for the Effective Management of Faba Bean Root Diseases in Ethiopia" Pathogens 14, no. 7: 695. https://doi.org/10.3390/pathogens14070695
APA StyleYilma, S., Bekele, B., Van Leur, J., You, M. P., Kemal, S.-A., Giblot-Ducray, D., Hill, K., Selvaraji, T., Lencho, A., Driba, L., & Barbetti, M. J. (2025). Defining Soilborne Pathogen Complexes Provides a New Foundation for the Effective Management of Faba Bean Root Diseases in Ethiopia. Pathogens, 14(7), 695. https://doi.org/10.3390/pathogens14070695