
Copper-Induced Stimulation of Ectophosphatase Activity of Candida albicans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms and Growth Conditions
2.2. Materials
2.3. Ectophosphatase Activity Measurements
2.4. Statistical Analysis
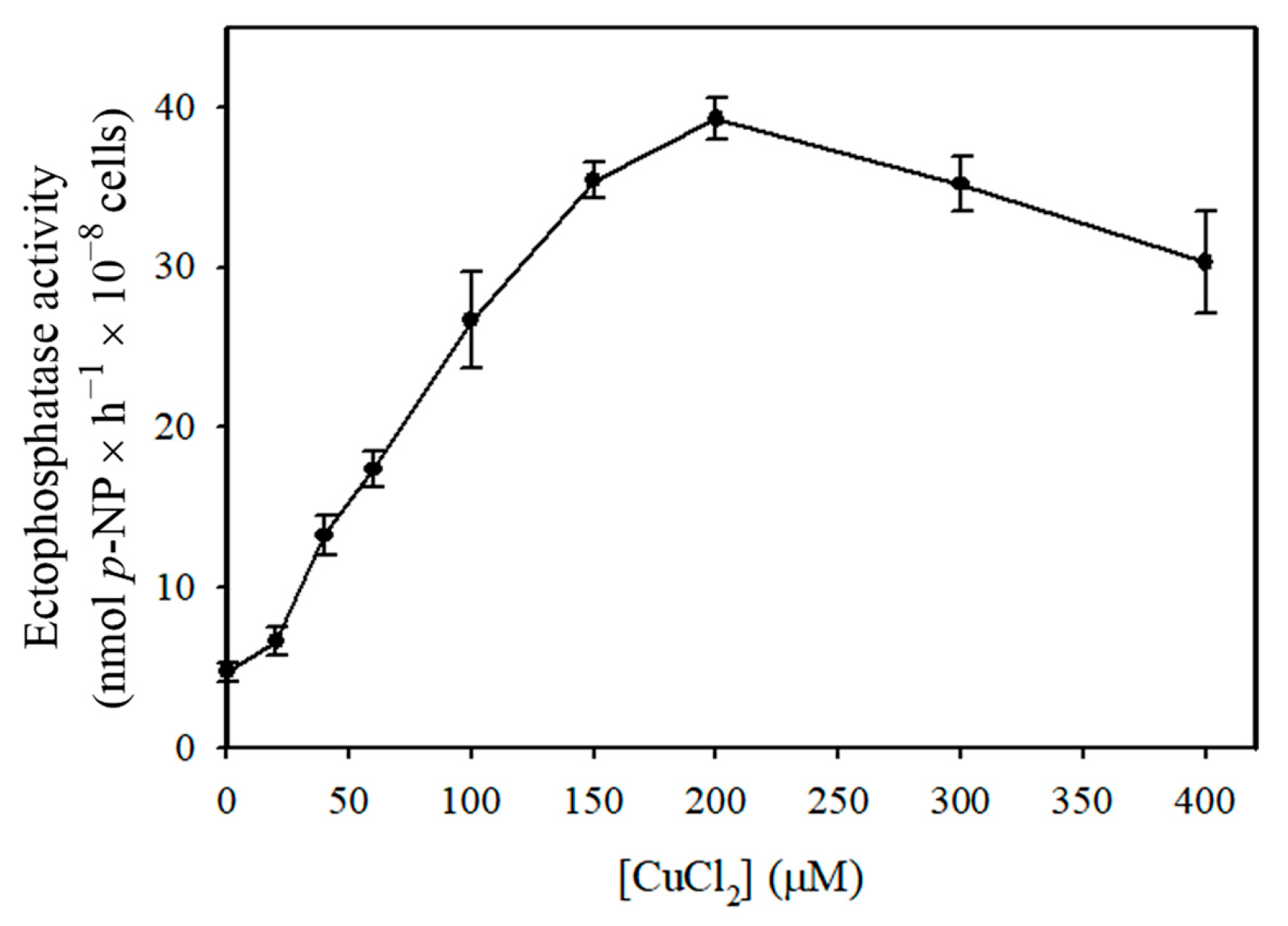
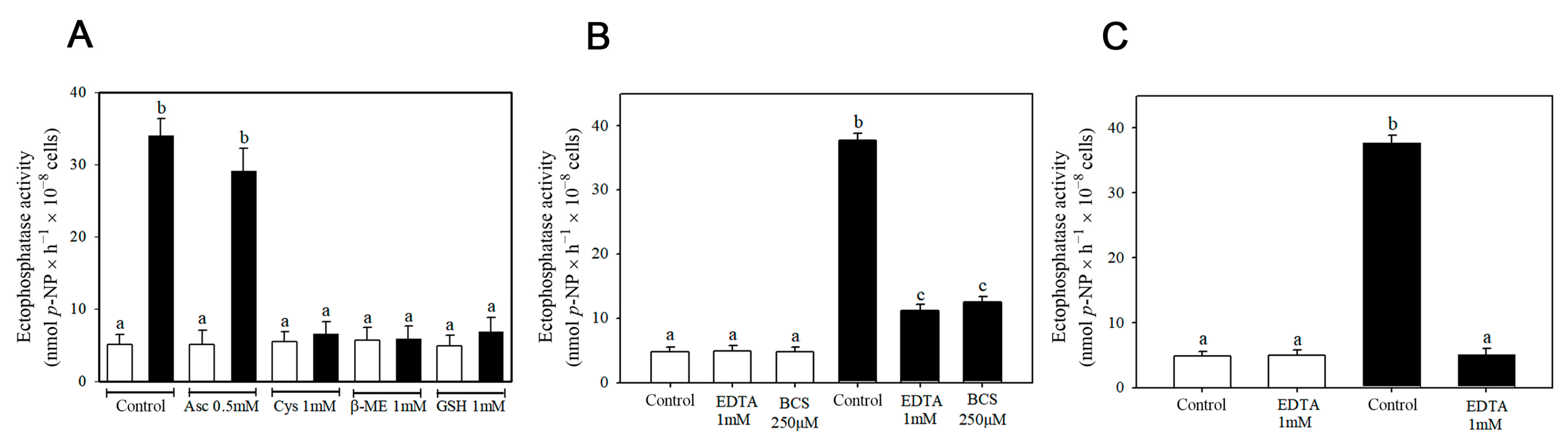
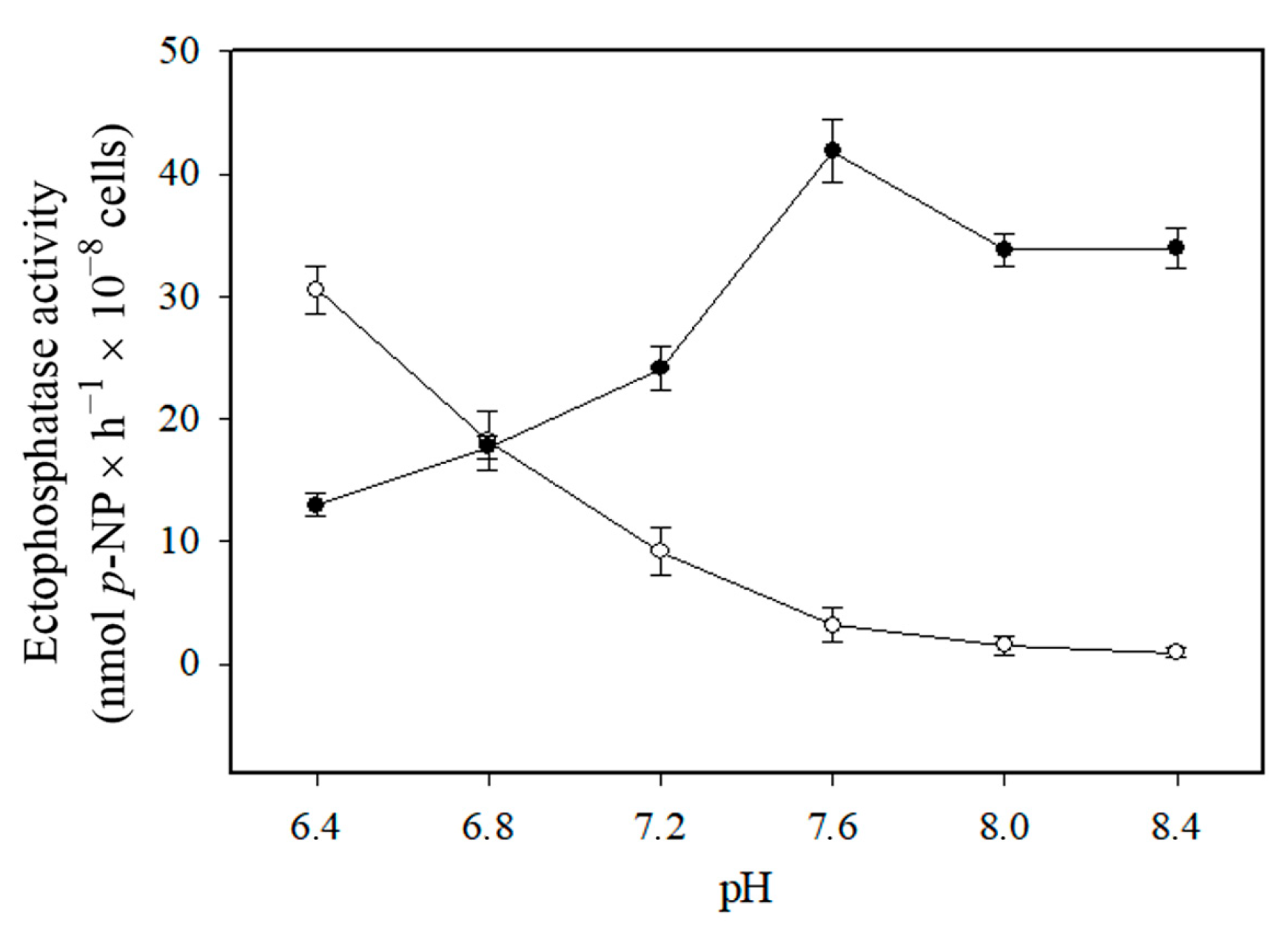
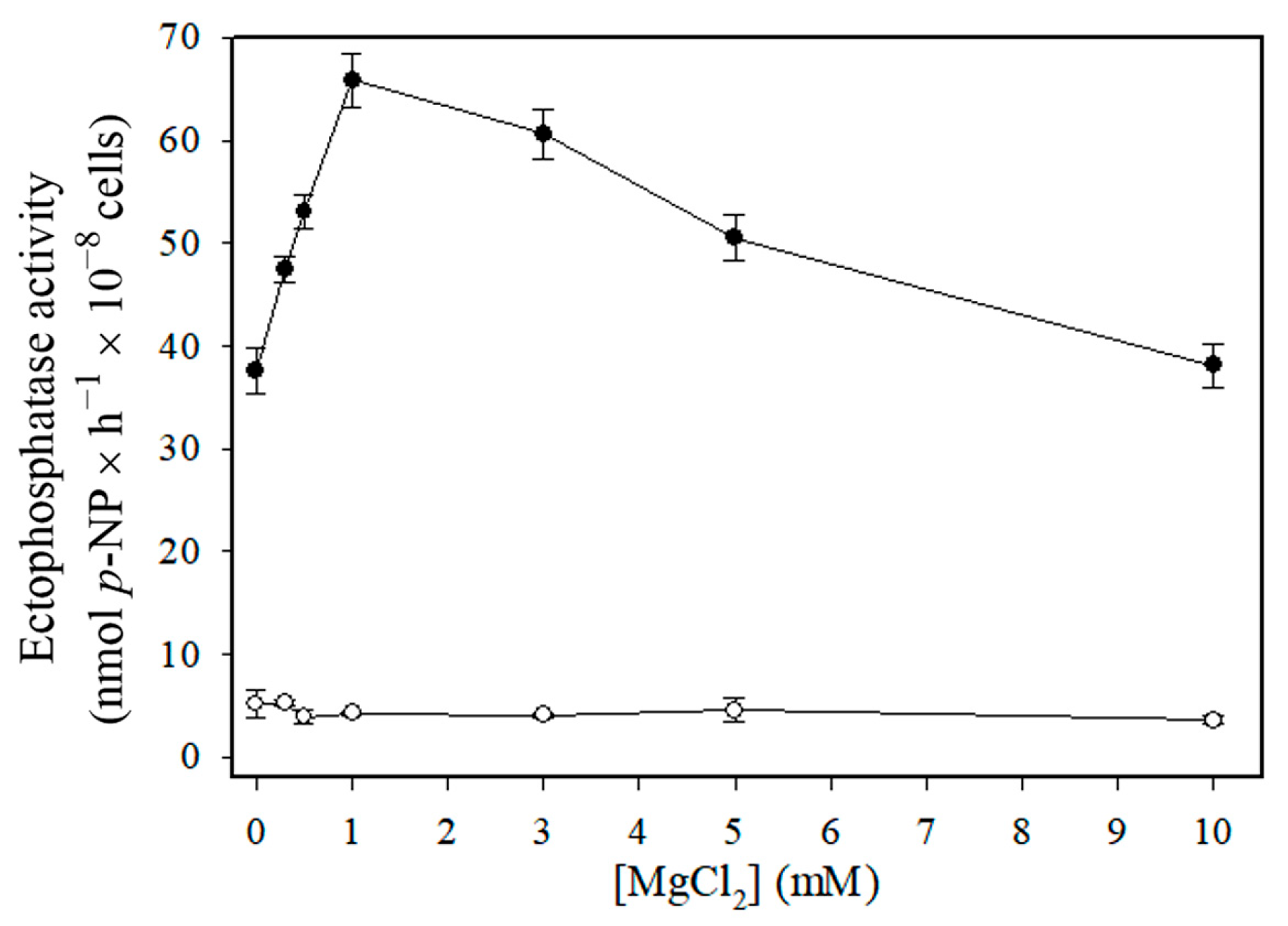
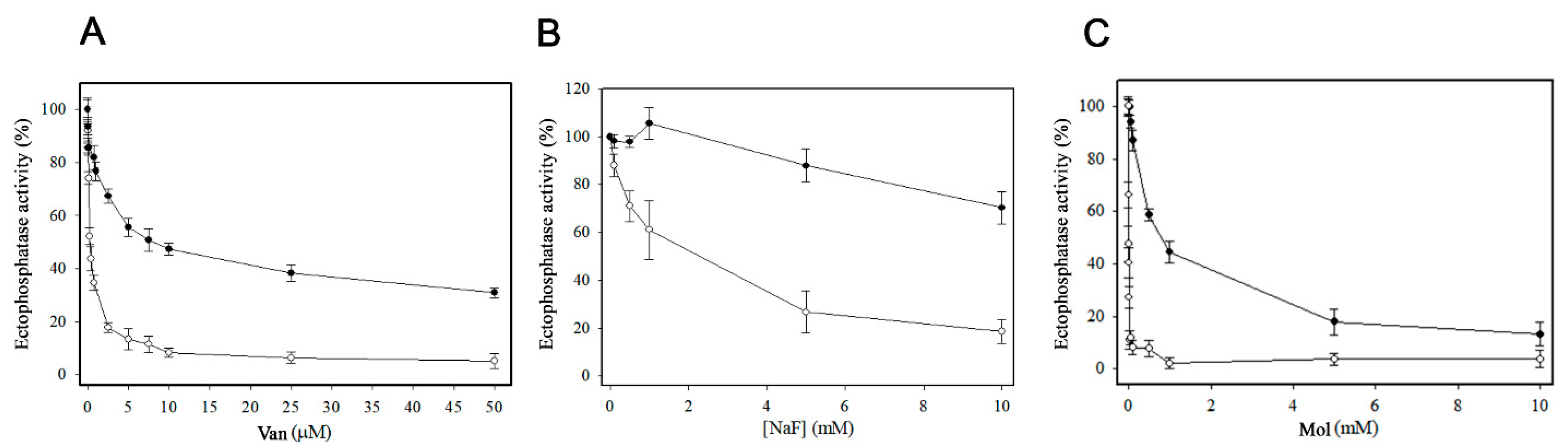
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| p-NPP | p-nitrophenyl phosphate |
| Asc | Ascorbate |
| Cys | Cysteine |
| β-ME | β-mercaptoethanol |
| GSH | Glutathione |
| EDTA | Ethylenediaminetetraacetic acid |
| BCS | Bathocuproinedisulfonic acid disodium salt |
| Van | Sodium orthovanadate |
| NaF | Sodium fluoride |
| Mol | Ammonium molybdate |
| ROS | Reactive oxygen species |
References
- Kriegl, L.; Egger, M.; Boyer, J.; Hoenigl, M.; Krause, R. New treatment options for critically important WHO fungal priority pathogens. Clin. Microbiol. Infect. 2024, in press. [Google Scholar] [CrossRef]
- Li, C.X.; Gleason, J.E.; Zhang, S.X.; Bruno, V.M.; Cormack, B.P.; Culotta, V.C. Candida albicans adapts to host copper during infection by swapping metal cofactors for superoxide dismutase. Proc. Natl. Acad. Sci. USA 2015, 112, E5336–E5342. [Google Scholar] [CrossRef]
- Miceli, M.H.; Díaz, J.A.; Lee, S.A. Emerging opportunistic yeast infections. Lancet Infect. Dis. 2011, 11, 142–151. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef]
- Keyvanfar, A.; Najafiarab, H.; Talebian, N.; Falah Tafti, M.; Adeli, G.; Ghasemi, Z.; Tehrani, S. Drug-resistant oral candidiasis in patients with HIV infection: A systematic review and meta-analysis. BMC Infect. Dis. 2024, 24, 546. [Google Scholar] [CrossRef]
- Portela, M.B.; Kneipp, L.F.; Ribeiro de Souza, I.P.; Holandino, C.; Alviano, C.S.; Meyer-Fernandes, J.R.; de Araújo Soares, R.M. Ectophosphatase activity in Candida albicans influences fungal adhesion: Study between HIV-positive and HIV-negative isolates. Oral Dis. 2010, 16, 431–437. [Google Scholar] [CrossRef]
- Talapko, J.; Juzbašić, M.; Matijević, T.; Pustijanac, E.; Bekić, S.; Kotris, I.; Škrlec, I. Candida albicans—The virulence factors and clinical manifestations of infection. J. Fungi 2021, 7, 79. [Google Scholar] [CrossRef]
- Collopy-Junior, I.; Esteves, F.F.; Nimrichter, L.; Rodrigues, M.L.; Alviano, C.S.; Meyer-Fernandes, J.R. An ectophosphatase activity in Cryptococcus neoformans. FEMS Yeast Res. 2006, 6, 1010–1017. [Google Scholar] [CrossRef]
- Cosentino-Gomes, D.; Rocco-Machado, N.; Santi, L.; Broetto, L.; Vainstein, M.H.; Meyer-Fernandes, J.R.; Schrank, A.; Beys-da-Silva, W.O. Inhibition of ectophosphatase activity in conidia reduces adhesion and virulence of Metarhizium anisopliae on the host insect Dysdercus peruvianus. Curr. Microbiol. 2013, 66, 467–474. [Google Scholar] [CrossRef]
- Kiffer-Moreira, T.; Pinheiro, A.A.S.; Alviano, W.S.; Barbosa, F.M.; Souto-Padrón, T.; Nimrichter, L.; Rodrigues, M.L.; Alviano, C.S.; Meyer-Fernandes, J.R. An ectophosphatase activity in Candida parapsilosis influences the interaction of fungi with epithelial cells. FEMS Yeast Res. 2007, 7, 621–628. [Google Scholar] [CrossRef]
- Kneipp, L.F.; Magalhães, A.S.; Abi-Chacra, E.A.; Souza, L.O.P.; Alviano, C.S.; Santos, A.L.S.; Meyer-Fernandes, J.R. Surface phosphatase in Rhinocladiella aquaspersa: Biochemical properties and its involvement with adhesion. Med. Mycol. 2012, 50, 570–578. [Google Scholar] [CrossRef]
- Kneipp, L.F.; Rodrigues, M.L.; Holandino, C.; Esteves, F.F.; Souto-Padrón, T.; Alviano, C.S.; Travassos, L.R.; Meyer-Fernandes, J.R. Ectophosphatase activity in conidial forms of Fonsecaea pedrosoi is modulated by exogenous phosphate and influences fungal adhesion to mammalian cells. Microbiology 2004, 150, 3355–3362. [Google Scholar] [CrossRef]
- Freitas-Mesquita, A.L.; Meyer-Fernandes, J.R. Biochemical properties and possible roles of ectophosphatase activities in fungi. Int. J. Mol. Sci. 2014, 15, 2289–2304. [Google Scholar] [CrossRef]
- Douglas, L.M.; Konopka, J.B. Plasma membrane architecture protects Candida albicans from killing by copper. PLoS Genet. 2019, 15, e1007911. [Google Scholar] [CrossRef]
- Falcone, E.; Stellato, F.; Vileno, B.; Bouraguba, M.; Lebrun, V.; Ilbert, M.; Morante, S.; Faller, P. Revisiting the pro-oxidant activity of copper: Interplay of ascorbate, cysteine, and glutathione. Metallomics 2023, 15, mfad040. [Google Scholar] [CrossRef]
- Weissman, Z.; Berdicevsky, I.; Cavari, B.Z.; Kornitzer, D. The high copper tolerance of Candida albicans is mediated by a P-type ATPase. Proc. Natl. Acad. Sci. USA 2000, 97, 3520–3525. [Google Scholar] [CrossRef]
- Fonzi, W.A.; Irwin, M.Y. Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar] [CrossRef]
- Fernandes, E.C.; Meyer-Fernandes, J.R.; Silva-Neto, M.A.C.; Vercesi, A.E. Trypanosoma brucei: Ecto-phosphatase activity on the surface of intact procyclic forms. Z. Naturforsch. C 1997, 52, 351–355. [Google Scholar] [CrossRef]
- Patterson, M.K., Jr. Measurement of growth and viability of cells in culture. Methods Enzymol. 1979, 58, 141–152. [Google Scholar]
- Fernandes, E.C.; Granjeiro, J.M.; Aoyama, H.; Fonseca, F.V.; Meyer-Fernandes, J.R.; Vercesi, A.E. A metallo phosphatase activity present on the surface of Trypanosoma brucei procyclic forms. Vet. Parasitol. 2003, 118, 19–28. [Google Scholar] [CrossRef]
- Freitas-Mesquita, A.L.; Carvalho-Kelly, L.F.; Majerowicz, T.S.S.; Meyer-Fernandes, J.R. Euglena gracilis: Biochemical properties of a membrane bound ecto-phosphatase activity modulated by fluoroaluminate complexes and different trophic conditions. Eur. J. Protistol. 2023, 91, 126010. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Inouhe, M.; Tohoyama, H.; Joho, M. Effect of copper on acid phosphatase activity in yeast Yarrowia lipolytica. Z. Naturforsch. C 2007, 62, 70–76. [Google Scholar] [CrossRef]
- Jonsson, C.M.; Aoyama, H. Effect of copper on the activation of the acid phosphatase from the green algae Pseudokirchneriella subcapitata. Biometals 2010, 23, 93–98. [Google Scholar] [CrossRef]
- Srivastava, D.K.; Pandey, K.C. Effect of copper on tissue acid and alkaline phosphatases in the green snakehead, Ophiocephalus punctatus (Bloch). Toxicol. Lett. 1982, 11, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Graham, N.M.H.; Sorensen, D.; Odaka, N.; Brookmeyer, R.; Chan, D.; Willett, W.C.; Morris, J.S.; Saah, A.J. Relationship of serum copper and zinc levels to HIV-1 seropositivity and progression to AIDS. J. Acquir. Immune Defic. Syndr. 1991, 4, 976–980. [Google Scholar]
- Kassu, A.; Yabutani, T.; Mulu, A.; Tessema, B.; Ota, F. Serum zinc, copper, selenium, calcium, and magnesium levels in pregnant and non-pregnant women in Gondar, Northwest Ethiopia. Biol. Trace Elem. Res. 2008, 122, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Moreno, T.; Artacho, R.; Navarro, M.; Pérez, A.; Ruiz-López, M.D. Serum copper concentration in HIV-infection patients and relationships with other biochemical indices. Sci. Total Environ. 1998, 217, 21–26. [Google Scholar] [CrossRef]
- Onwuli, D.; Waribo-Anthony, H.; Anyalebechi, O.; Ugochukwu, C. CD4+ T-Cell Count, Serum Zinc, Copper and Selenium Levels in HIV Sero-Positive Subjects on ART and ART Naïve Subjects in Port Harcourt, Nigeria. J. Biosci. Med. 2022, 10, 105–116. [Google Scholar]
- Ding, X.; Hua, Y.; Chen, Y.; Zhang, C.; Kong, X. Heavy metal complexation of thiol-containing peptides from soy glycinin hydrolysates. Int. J. Mol. Sci. 2015, 16, 8040–8058. [Google Scholar] [CrossRef]
- Krężel, A.; Lesniak, W.; Jezowska-Bojczuk, M.; Mlynarz, P.; Brasuñ, J.; Kozlowski, H.; Bal, W. Coordination of heavy metals by dithiothreitol, a commonly used thiol group protectant. J. Inorg. Biochem. 2001, 84, 77–88. [Google Scholar] [CrossRef]
- Ufnalska, I.; Drew, S.C.; Zhukov, I.; Szutkowski, K.; Wawrzyniak, U.E.; Wróblewski, W.; Frączyk, T.; Bal, W. Intermediate Cu(II)-thiolate species in the reduction of Cu(II)GHK by glutathione: A handy chelate for biological Cu(II) reduction. Inorg. Chem. 2021, 60, 18048–18057. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Griffiths, P.T.; Campbell, S.J.; Utinger, B.; Kalberer, M.; Paulson, S.E. Ascorbate oxidation by iron, copper and reactive oxygen species: Review, model development, and derivation of key rate constants. Sci. Rep. 2021, 11, 7417. [Google Scholar] [CrossRef]
- Paletta-Silva, R.; Vieira-Bernardo, R.; Cosentino-Gomes, D.; Meyer-Fernandes, J.R. Leishmania amazonensis: Inhibition of 3’-nucleotidase activity by Cu2⁺ ions. Exp. Parasitol. 2012, 131, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Rubino, J.T.; Franz, K.J. Coordination chemistry of copper proteins: How nature handles a toxic cargo for essential function. J. Inorg. Biochem. 2012, 107, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Hung, H.-C.; Chang, G.-G. Differentiation of the slow-binding mechanism for magnesium ion activation and zinc ion inhibition of human placental alkaline phosphatase. Protein Sci. 2001, 10, 131–140. [Google Scholar] [CrossRef]
- Nam, K.; Arattu Thodika, A.R.; Tischlik, S.; Phoeurk, C.; Nagy, T.M.; Schierholz, L.; Ådén, J.; Rogne, P.; Drescher, M.; Sauer-Eriksson, A.E.; et al. Magnesium-induced structural reorganization in the active site of adenylate kinase. Sci. Adv. 2024, 10, eado5504. [Google Scholar] [CrossRef]
- Mackie, J.; Szabo, E.K.; Urgast, D.S.; Ballou, E.R.; Childers, D.S.; MacCallum, D.M.; Feldmann, J.; Brown, A.J.P. Host-imposed copper poisoning impacts fungal micronutrient acquisition during systemic Candida albicans infections. PLoS ONE 2016, 11, e0158683. [Google Scholar] [CrossRef]
- Li, C.; Li, Y.; Ding, C. The role of copper homeostasis at the host-pathogen axis: From bacteria to fungi. Int. J. Mol. Sci. 2019, 20, 175. [Google Scholar] [CrossRef]
- Jawale, C.V.; Biswas, P.S. Local antifungal immunity in the kidney in disseminated candidiasis. Curr. Opin. Microbiol. 2021, 62, 1–7. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freitas-Mesquita, A.L.; Esteves, F.F.; Meyer-Fernandes, J.R. Copper-Induced Stimulation of Ectophosphatase Activity of Candida albicans. Pathogens 2025, 14, 667. https://doi.org/10.3390/pathogens14070667
Freitas-Mesquita AL, Esteves FF, Meyer-Fernandes JR. Copper-Induced Stimulation of Ectophosphatase Activity of Candida albicans. Pathogens. 2025; 14(7):667. https://doi.org/10.3390/pathogens14070667
Chicago/Turabian StyleFreitas-Mesquita, Anita Leocadio, Fabiano Ferreira Esteves, and José Roberto Meyer-Fernandes. 2025. "Copper-Induced Stimulation of Ectophosphatase Activity of Candida albicans" Pathogens 14, no. 7: 667. https://doi.org/10.3390/pathogens14070667
APA StyleFreitas-Mesquita, A. L., Esteves, F. F., & Meyer-Fernandes, J. R. (2025). Copper-Induced Stimulation of Ectophosphatase Activity of Candida albicans. Pathogens, 14(7), 667. https://doi.org/10.3390/pathogens14070667