
Antiprotozoal Effects of Pediococcus acidilactici-Derived Postbiotic on Blastocystis Subtypes ST1/ST3

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Postbiotic Production Process
2.2. Establishment of Experimental Groups
- Metronidazole group: Metronidazole (Nidazol, I.E. Ulagay Pharmaceutical Industry, İstanbul, Türkiye) was used as the reference drug. A stock solution was prepared by dissolving 0.8 mg of metronidazole in 1 mL of distilled water to achieve a final concentration of 0.05 mg/mL in the culture medium.
- Control group: Sterile distilled water was used as a negative control.
- MRS broth group: MRS broth was dissolved in distilled water to prepare a culture medium at a final concentration of 210 mg/mL. The MRS broth medium (210 mg/mL) without added bacteria was centrifuged at 4200× g for 10 min at 4 °C, and the supernatant was used by filtering using 0.45 µm filters. Subsequently, serial dilutions were performed with sterile distilled water to obtain final concentrations of 140, 70, 35, 17.5, and 8.75 mg/mL.
- P. acidilactici group: The fresh P. acidilactici postbiotic obtained after lyophilization was dissolved in sterile distilled water to prepare a stock solution at a concentration of 210 mg/mL. Subsequently, serial dilutions were performed with sterile distilled water to obtain final concentrations of 140, 70, 35, 17.5, and 8.75 mg/mL.
2.3. The Isolation of a Blastocystis Isolate
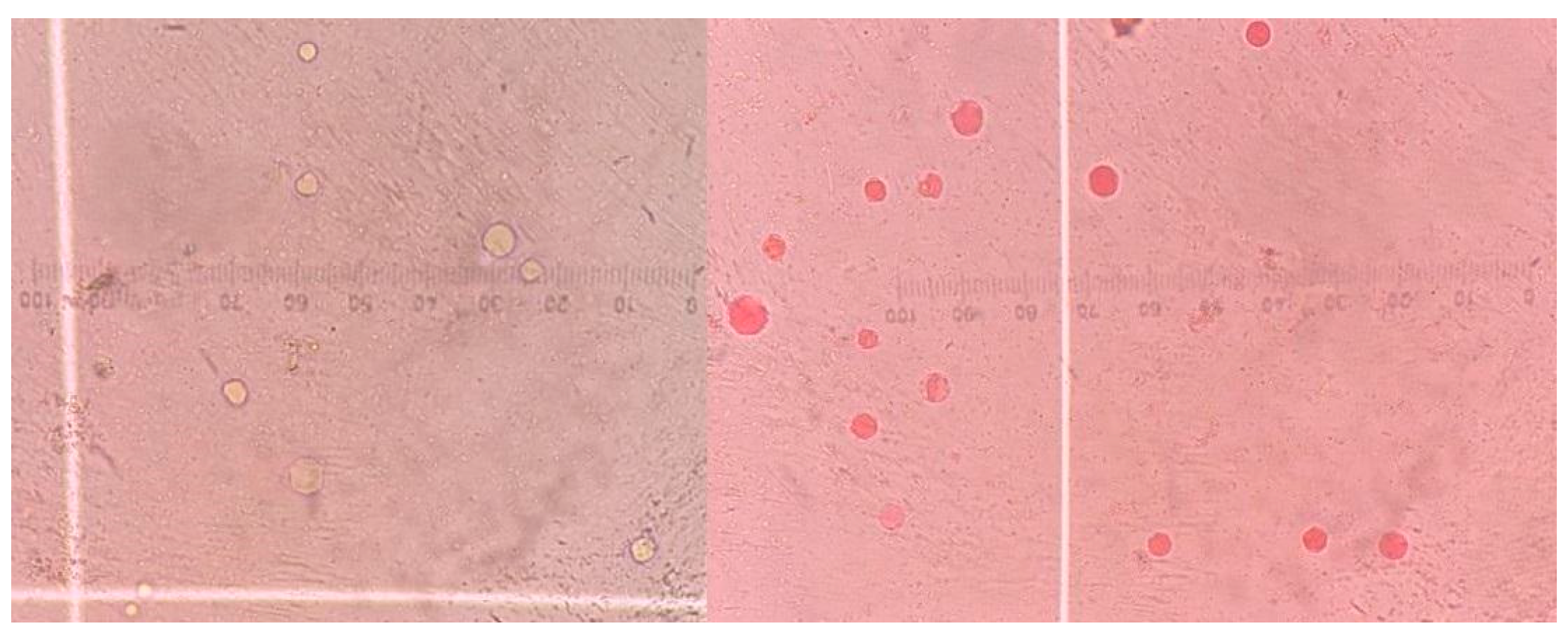
2.4. Antiprotozoal Activity of the Postbiotic on Blastocystis
2.5. pH Measurements of the Culture Media
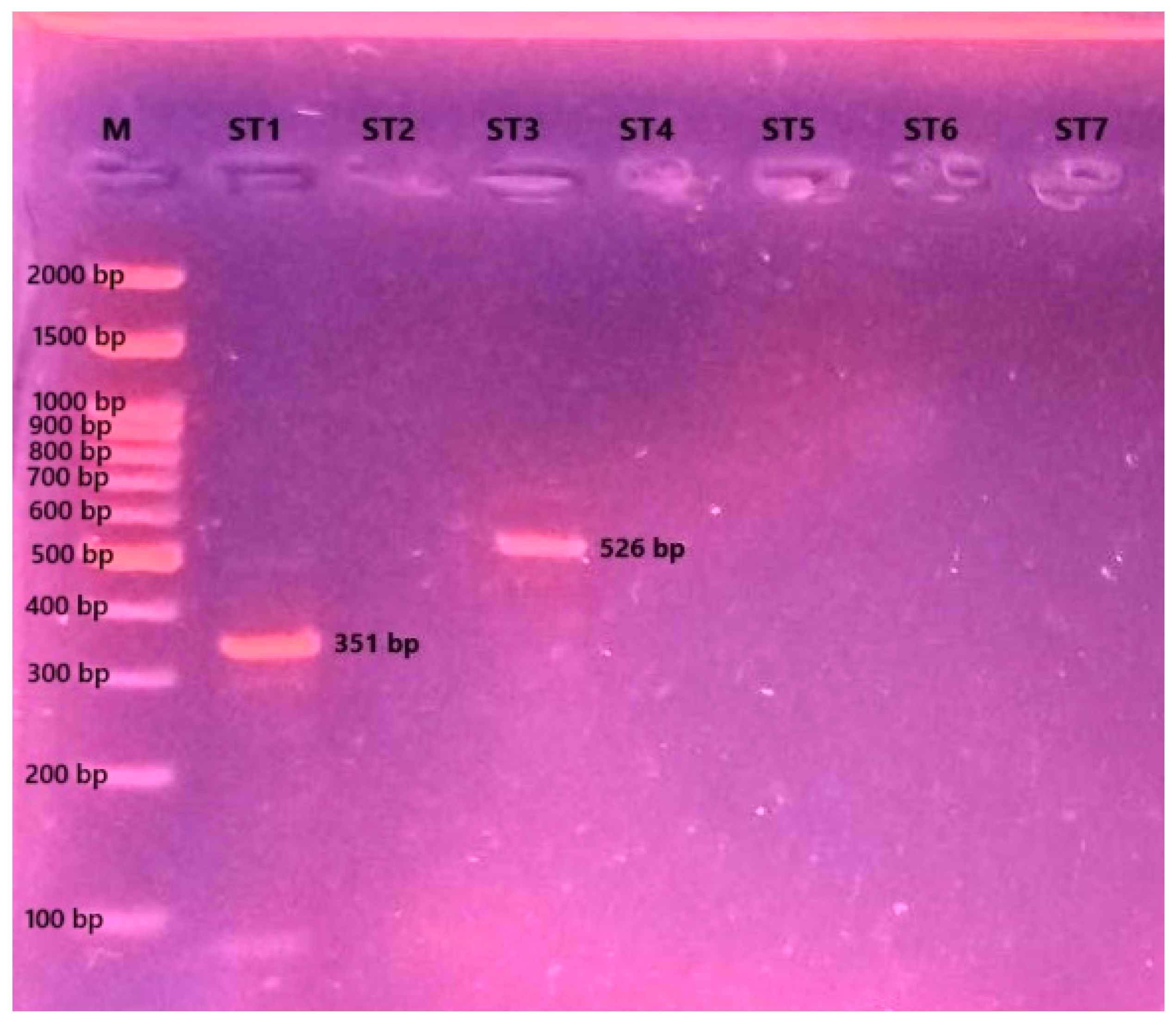
2.6. Determination of Blastocystis Subtypes
2.7. Postbiotic Characterization
2.7.1. Analysis of the Phenolic Compounds
2.7.2. Analysis of the Organic Acid Content
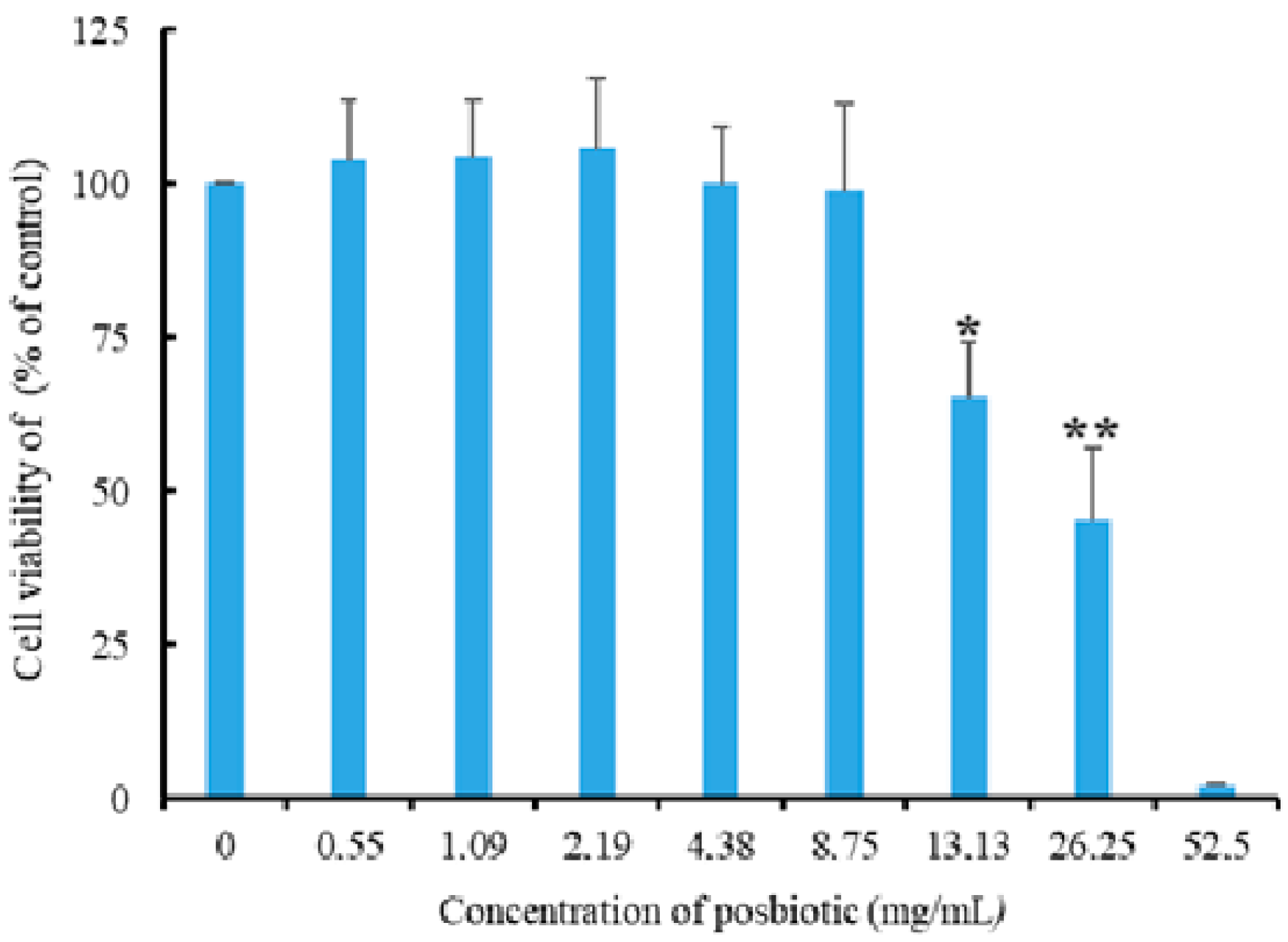
2.8. Cell Viability Assay
2.9. Statistical Analysis
3. Results
3.1. Postbiotic Production
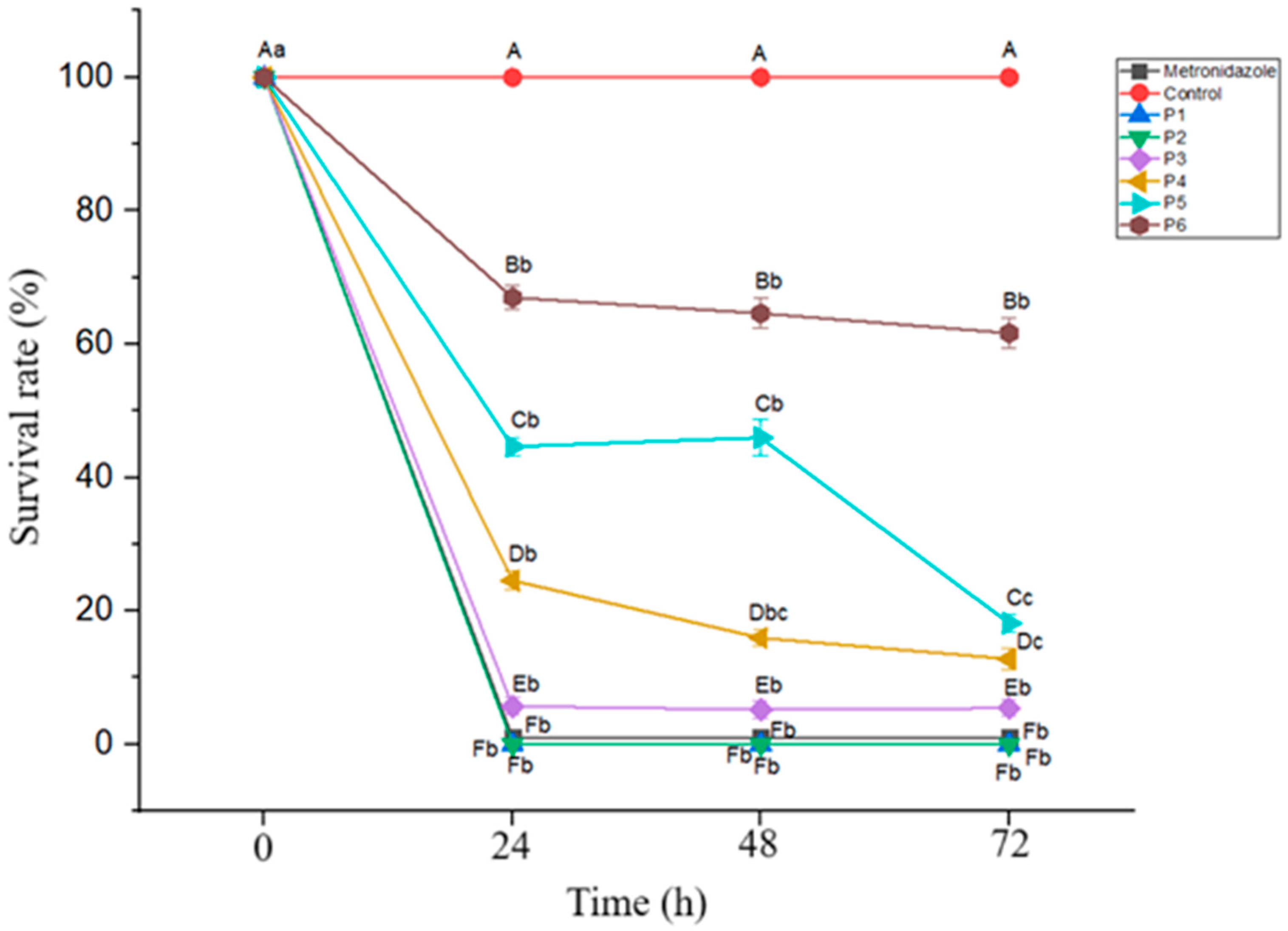
3.2. Antiprotozoal Activity of the Postbiotic on Blastocystis
3.3. pH Measurement of the Culture Medium
3.4. Blastocystis Subtype Identification
3.5. Characterization of the Postbiotic
3.6. Cytotoxic Effect of the Postbiotic
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Aydemir, S.; Barlik, F.; Yurekturk, S.; Saygin, M.; Unlu, A.H.; Ekici, A.; Yilmaz, H. Prevalence of Blastocystis infection in humans in Turkiye: A systematic review and meta-analysis. Microb. Pathog. 2024, 195, 106876. [Google Scholar] [CrossRef] [PubMed]
- Sulzyc-Bielicka, V.; Kolodziejczyk, L.; Adamska, M.; Skotarczak, B.; Jaczewska, S.; Safranow, K.; Bielicki, P.; Kładny, J.; Bielicki, D. Colorectal cancer and Blastocystis sp. infection. Parasites Vectors 2021, 14, 200. [Google Scholar] [CrossRef]
- Matovelle, C.; Quilez, J.; Tejedor, M.T.; Beltran, A.; Chueca, P.; Monteagudo, L.V. Subtype distribution of Blastocystis spp. in patients with gastrointestinal symptoms in northern Spain. Microorganisms 2024, 12, 1084. [Google Scholar] [CrossRef]
- Mokhtar, A.B.; Ahmed, S.A.; Eltamany, E.E.; Karanis, P. Anti-Blastocystis activity in vitro of Egyptian herbal extracts (Family: Asteraceae) with emphasis on Artemisia judaica. Int. J. Environ. Res. Public Health 2019, 16, 1555. [Google Scholar] [CrossRef] [PubMed]
- Rudzinska, M.; Sikorska, K. Epidemiology of Blastocystis infection: A review of data from Poland in relation to other reports. Pathogens 2023, 12, 1050. [Google Scholar] [CrossRef]
- Abedi, S.H.; Fazlzadeh, A.; Mollalo, A.; Sartip, B.; Mahjour, S.; Bahadory, S.; Taghipour, A.; Rostami, A. The neglected role of Blastocystis sp. and Giardia lamblia in development of irritable bowel syndrome: A systematic review and meta-analysis. Microb. Pathog. 2022, 162, 105215. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, F.; Babaei, E.; Badirzadeh, A.; Riabi, T.R.; Abdoli, A. Blastocystis, urticaria, and skin disorders: Review of the current evidences. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1027–1042. [Google Scholar] [CrossRef]
- Deng, L.; Wojciech, L.; Gascoigne, N.R.J.; Peng, G.N.; Tan, K.S.W. New insights into the interactions between Blastocystis, the gut microbiota, and host immunity. PLoS Pathog. 2021, 17, e1009253. [Google Scholar] [CrossRef]
- Deng, L.; Lee, J.W.J.; Tan, K.S.W. Infection with pathogenic Blastocystis ST7 is associated with decreased bacterial diversity and altered gut microbiome profiles in diarrheal patients. Parasites Vectors 2022, 15, 312. [Google Scholar] [CrossRef]
- Ahmed, S.A.A.; Schou, C.; Mokhtar, A.B.; Karanis, P.; Gad, S.E.M. Blastocystis species growth inhibition in vitro by plant extracts. Microb. Pathog. 2024, 196, 106970. [Google Scholar] [CrossRef]
- Singh, A.; Priyadarshi, K.; Rai, T.; Banerjee, T. A case report of Blastocystis infection and Stevens—Johnson syndrome. Trop. Biomed. 2019, 36, 987–992. [Google Scholar] [PubMed]
- Montero, J.A.; Álvarez, S.S.; Anaut, M.B.; Medrano, R.L.; Esteras, M.A.R.; Ruiz, M.R.; García, F.M.I. Blastocystis hominis-associated acute appendicular peritonitis in a 9-year-old boy: A case report and a comprehensive review of the literature. Pediatr. Infect. Dis. J. 2024, 43, 327–330. [Google Scholar] [CrossRef]
- Kurt, Ö.; Dogruman Al, F.; Tanyüksel, M. Eradication of Blastocystis in humans: Really necessary for all? Parasitol. Int. 2016, 65, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Cobuccio, L.G.; Laurent, M.; Gardiol, C.; Wampfler, R.; Poppert, S.; Senn, N.; Eperon, G.; Genton, B.; Locatelli, I.; Valliere, S. Should we treat Blastocystis sp.? A double-blind placebo-controlled randomized pilot trial. J. Travel Med. 2023, 30, 143. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, T.C.; Cruz, A.G.; Pereira, E.; da Costa, W.K.A.; Rocha, R.d.S.; Pedrosa, G.T.d.S.; Rocha, C.d.S.; Alves, J.M.; Alvarenga, V.O.; Sant’aNa, A.S.; et al. Postbiotics: An overview of concepts, inactivation technologies, health effects, and driver trends. Trends Food Sci. Technol. 2023, 138, 199–214. [Google Scholar] [CrossRef]
- Teame, T.; Wang, A.; Xie, M.; Zhang, Z.; Yang, Y.; Ding, Q.; Gao, C.; Olsen, R.E.; Ran, C.; Zhou, Z. Paraprobiotics and postbiotics of probiotic Lactobacilli, their positive effects on the host and action mechanisms: A review. Front. Nutr. 2020, 7, 570344. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Incili, G.K.; Karatepe, P.; Akgöl, M.; Kaya, B.; Kanmaz, H.; Hayaloglu, A.A. Characterization of Pediococcus acidilactici postbiotic and impact of postbiotic-fortified chitosan coating on the microbial and chemical quality of chicken breast fillets. Int. J. Biol. Macromol. 2021, 184, 429–437. [Google Scholar] [CrossRef]
- Ramesh, L.; Dharumadurai, D. Antiprotozoan activity of postbiotics. In Postbiotics; Springer: Berlin/Heidelberg, Germany, 2023; pp. 207–210. [Google Scholar]
- Cuellar-Guevara, F.L.; Barron-Gonzalez, M.P.; Menchaca-Arredondo, J.L. Effect of Lactobacillus postbiotics on Entamoeba histolytica trophozoites. Rev. Investig. Clin. 2019, 71, 402–407. [Google Scholar] [CrossRef]
- Incili, G.; Akgöl, M.; Karatepe, P.; Kanmaz, H.; Kaya, B.; Tekin, A.; Hayaloğlu, A.A. Inhibitory effect of bioactive compounds derived from freeze-dried paraprobiotic of Pediococcus acidilactici against food-borne pathogens: In vitro and food model studies. Food Res. Int. 2023, 170, 113045. [Google Scholar] [CrossRef]
- Yıldız, S.; Aydemir, S.; Ekici, A.; Deniz, N.Y.; Yilmaz, H. Inhibitory effect of thymoquinone and capsaicin on Blastocystis grown in vitro. Indian J. Exp. Biol. 2024, 62, 423–428. [Google Scholar]
- El-Sayed, N.M. Evaluation of the in vitro effects of ethanol extracts of Ocimum basilicum and Thymus vulgaris for anti-Blastocystis hominis activity. Egypt J. Med. Sci. 2009, 30, 1229–1243. [Google Scholar]
- Aykur, M.; Karakavuk, E.; Karakavuk, M.; Akıl, M.; Can, H.; Döşkaya, M.; Gürüz, Y.; Dağcı, H. Inhibitory effect of Tunceli garlic (Allium tuncelianum) on Blastocystis subtype 3 grown in vitro. Expert Opin. Orphan Drugs. 2020, 8, 489–496. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Wu, Z.; Kimata, I.; Iseki, M.; Ali, I.K.M.D.; Hossain, M.B.; Zaman, V.; Haque, R.; Takahashi, Y. Polymerase chain reaction-based genotype classification among human Blastocystis hominis populations isolated from different countries. Parasitol. Res. 2004, 92, 22–29. [Google Scholar] [PubMed]
- Başar, Y.; Yiğit, A.; Tunç, A.K.; Sarıtaş, B.M. Lavandula stoechas extract; synthesis of silver nanoparticles (nature-friendly green synthesis method), characterization, antimicrobial activity and in silico molecular docking study. Curr. Perspect. Med. Aromat. Plants 2023, 7, 24–33. [Google Scholar] [CrossRef]
- Aksoy, A.; Altunatmaz, S.S.; Aksu, F.; Tokatlı Demirok, N.; Yazıcı, K.; Yıkmış, S. Bee bread as a functional product: Phenolic compounds, amino acid, sugar, and organic acid profiles. Foods 2024, 13, 795. [Google Scholar] [CrossRef]
- Ooi, M.F.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Rahim, R.A.; Ariff, A. A refined medium to enhance the antimicrobial activity of postbiotic produced by Lactiplantibacillus plantarum RS5. Sci. Rep. 2021, 11, 7617. [Google Scholar] [CrossRef]
- Amiri, S.; Mokarram, R.R.; Khiabani, M.S.; Bari, M.R.; Alizadeh, M. Optimization of food-grade medium for co-production of bioactive substances by Lactobacillus acidophilus LA-5 for explaining pharmabiotic mechanisms of probiotic. J. Food Sci. Technol. 2021, 58, 1–12. [Google Scholar] [CrossRef]
- Incili, G.K.; Karatepe, P.; Akgol, M.; Gungoren, A.; Koluman, A.; Ilhak, O.I.; Kanmaz, H.; Kaya, B.; Hayaloğlu, A.A. Characterization of lactic acid bacteria postbiotics: Evaluation of in vitro antibacterial effect, microbial and chemical quality on chicken drumsticks. Food Microbiol. 2022, 104, 104001. [Google Scholar] [CrossRef]
- Gao, R.; Zhang, X.; Huang, L.; Shen, R.; Qin, H. Gut microbiota alteration after long-term consumption of probiotics in the elderly. Probiotics Antimicrob. Proteins 2019, 11, 655–666. [Google Scholar] [CrossRef]
- Bourebaba, Y.; Marycz, K.; Mularczyk, M.; Bourebaba, L. Postbiotics as potential new therapeutic agents for metabolic disorders management. Biomed. Pharmacother. 2022, 153, 113138. [Google Scholar] [CrossRef] [PubMed]
- Lepczynska, M.; Dzika, E. The influence of probiotic bacteria and human gut microorganisms causing opportunistic infections on Blastocystis ST3. Gut Pathog. 2019, 11, 6. [Google Scholar] [CrossRef]
- Alkady, S.F.H.; Naggar, H.M.A.E.; Thabet, H.S.; El-Sayed, H.S.; Magdy, M.; Fahmy, I.A.; Abou-Seri, H.M. Assessment of Lactobacillus acidophilus (L. acidophilus) therapeutic and prophylactic role in rats experimentally infected with Blastocystis subtype 3 (ST3). Parasitol. Res. 2025, 124, 11. [Google Scholar] [CrossRef]
- Szajewska, H.; Kolodziej, M.; Gieruszczak-Bialek, D.; Skorka, A.; Ruszczynski, M.; Shamir, R. Systematic review with meta-analysis: Lactobacillus rhamnosus GG for treating acute gastroenteritis in children—A 2019 update. Aliment. Pharmacol. Ther. 2019, 49, 1376–1384. [Google Scholar] [CrossRef]
- Warda, A.K.; Clooney, A.G.; Ryan, F.; Bettio, P.H.d.A.; Di Benedetto, G.; Ross, R.P.; Hill, C.; Ercolini, D. A postbiotic consisting of heat-treated lactobacilli has a bifidogenic effect in pure culture and in human fermented fecal communities. Appl. Environ. Microbiol. 2021, 87, e02459-20. [Google Scholar] [CrossRef] [PubMed]
- Chelliah, R.; Saravanakumar, K.; Daliri, E.B.; Kim, J.H.; Lee, J.K.; Jo, H.Y.; Kim, S.H.; Ramakrishnan, S.R.; Madar, I.H.; Wei, S.; et al. Unveiling the potentials of bacteriocin (Pediocin L50) from Pediococcus acidilactici with antagonist spectrum in a Caenorhabditis elegans model. Int. J. Biol. Macromol. 2020, 143, 555–572. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, X.; Yue, Y.; Wang, X.; Zeng, X.; Guo, Q.; Yan, X.; Du, G.; Yuan, Y.; Yue, T. Enrichment and distribution of selenium in Pediococcus acidilactici MRS-7: Impact on its biochemical composition, microstructure, and gastrointestinal survival. J. Agric. Food Chem. 2022, 70, 14877–14885. [Google Scholar] [CrossRef] [PubMed]
- Ayinde, O.; Ross, J.D. The frequency and duration of side-effects associated with the use of oral metronidazole: A prospective study of VITA trial participants. Int. J. STD AIDS 2023, 34, 897–902. [Google Scholar] [CrossRef]
- Ceruelos, A.H.; Romero-Quezada, L.; Ledezma, J.R.; Contreras, L.L. Therapeutic uses of metronidazole and its side effects: An update. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 397–401. [Google Scholar]
- Rajamanikam, A.; Hooi, H.S.; Kudva, M.; Samudi, C.; Kumar, S. Resistance towards metronidazole in Blastocystis sp.: A pathogenic consequence. PLoS ONE 2019, 14, e0212542. [Google Scholar] [CrossRef]
- Farah Haziqah, M.T.; Chandrawathani, P.; Douadi, B.; Suresh, K.; Wilson, J.J.; Mohd Khalid, M.K.N.; Rajamanikam, A.; Lewis, J.W.; Mohd Zain, S.N. Impact of pH on the viability and morphology of Blastocystis isolates. Trop. Biomed. 2018, 35, 501–510. [Google Scholar] [PubMed]
- Abdel-Hafeez, E.; Ahmed, A.; Abdellatif, M.; Kamal, A.; Toni, M. The efficacy of pomegranate (Punica granatum) peel extract on experimentally infected rats with Blastocystis spp. J. Infect. Dis. Prev. Med. 2016, 4, 131. [Google Scholar] [CrossRef]
- Girish, S.; Kumar, S.; Aminudin, N. Eurycoma longifolia (Tongkat Ali): A possible therapeutic candidate against Blastocystis sp. Parasites Vectors 2015, 8, 332. [Google Scholar] [CrossRef] [PubMed]
- Kovanda, L.; Zhang, W.; Wei, X.; Luo, J.; Wu, X.; Atwill, E.R.; Vaessen, S.; Li, X.; Liu, Y. In vitro antimicrobial activities of organic acids and their derivatives on several species of Gram-negative and Gram-positive bacteria. Molecules 2019, 24, 3770. [Google Scholar] [CrossRef]
- Kwak, A.M.; Lee, I.K.; Lee, S.Y.; Yun, B.S.; Kang, H.W. Oxalic acid from Lentinula edodes culture filtrate: Antimicrobial activity on phytopathogenic bacteria and qualitative and quantitative analyses. Mycobiology 2016, 44, 338–342. [Google Scholar] [CrossRef]
- In, Y.W.; Kim, J.J.; Kim, H.J.; Oh, S.W. Antimicrobial activities of acetic acid, citric acid and lactic acid against Shigella species. J. Food Saf. 2013, 33, 79–85. [Google Scholar] [CrossRef]
- Borah, H.J.; Borah, A.; Yadav, A.; Hazarika, S. Extraction of malic acid from Dillenia indica in organic solvents and its antimicrobial activity. Sep. Sci. Technol. 2023, 58, 314–325. [Google Scholar] [CrossRef]
- Haque, M.; Chowdhury, R.; Islam, K.; Akbar, M. Propionic acid is an alternative to antibiotics in poultry diet. Bangladesh J. Anim. Sci. 2009, 38, 115–122. [Google Scholar] [CrossRef]
- Palomo-Ligas, L.; Estrada-Camacho, J.; Garza-Ontiveros, M.; Vargas-Villanueva, J.R.; Gutiérrez-Gutiérrez, F.; Nery-Flores, S.D.; Montoya, J.A.C.; Ascacio-Valdés, J.; Campos-Muzquiz, L.G.; Rodriguez-Herrera, R. Polyphenolic extract from Punica granatum peel causes cytoskeleton-related damage on Giardia lamblia trophozoites in vitro. PeerJ 2022, 10, e13350. [Google Scholar] [CrossRef]
- Méabed, E.M.; El-Sayed, N.M.; Abou-Sreea, A.I.; Roby, M.H. Chemical analysis of aqueous extracts of Origanum majorana and Foeniculum vulgare and their efficacy on Blastocystis spp. cysts. Phytomedicine 2018, 43, 158–163. [Google Scholar] [CrossRef]
- Saha, S.; Srivastava, R.; Sarma, P.; Bhatt, T.K.; Prakash, A.; Kumar, D. Identification of potential inhibitors of Leishmania donovani sterol 24-C-methyltransferase: In silico and in vitro studies. Mol. Simul. 2023, 49, 1311–1323. [Google Scholar] [CrossRef]
- Monzote, L.; Córdova, W.H.P.; García, M.; Piñón, A.; Setzer, W.N. In vitro and in vivo activities of phenolic compounds against cutaneous leishmaniasis. Rec. Nat. Prod. 2016, 10, 269–276. [Google Scholar]
- Oliveira, C.B.; Meurer, Y.S.; Oliveira, M.G.; Medeiros, W.M.; Silva, F.O.; Brito, A.C.; Pontes, D.D.L.; Andrade-Neto, V.F. Comparative study on the antioxidant and anti-Toxoplasma activities of vanillin and its resorcinarene derivative. Molecules 2014, 19, 5898–5912. [Google Scholar] [CrossRef] [PubMed]
- Alson, S.G.; Jansen, O.; Cieckiewicz, E.; Rakotoarimanana, H.; Rafatro, H.; Degotte, G.; Francotte, P.; Frederich, M. In vitro and in vivo antimalarial activity of caffeic acid and some of its derivatives. J. Pharm. Pharmacol. 2018, 70, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Aldaba-Muruato, L.R.; Ventura-Juárez, J.; Perez-Hernandez, A.M.; Hernández-Morales, A.; Muñoz-Ortega, M.H.; Martínez-Hernández, S.L.; Alvarado, B.; Macías, J.R. Therapeutic perspectives of p-coumaric acid: Anti-necrotic, anti-cholestatic and anti-amoebic activities. World Acad. Sci. J. 2021, 3, 47. [Google Scholar] [CrossRef]
- Ren, L.; Wang, S.; Liu, S.; Prasanthi, H.A.C.; Li, Y.; Cao, J.; Zhong, F.; Guo, L.; Lu, F.; Luo, X. Postbiotic of Pediococcus acidilactici GQ01, a novel probiotic strain isolated from natural fermented wolfberry, attenuates hyperuricaemia in mice through modulating uric acid metabolism and gut microbiota. Foods 2024, 13, 923. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subtype (ST) | Primers | Long (bp) |
|---|---|---|
| ST1 | SB83F 5′-GAAGGACTCTCTGACGATGA-3′ SB83R 5′-GTCCAAATGAAAGGCAGC-3′ | 351 |
| ST2 | SB340F 5′-TGTTCTTGTGTCTTCTCAGCTC-3′ SB340R 5′-TTCTTTCACACTCCCGTCAT-3′ | 704 |
| ST3 | SB227F 5′-TAGGATTTGGTGTTTGGAGA-3′ SB227R 5′-TTAGAAGTGAAGGAGATGGAAG-3′ | 526 |
| ST4 | SB337F 5′-GTCTTTCCCTGTCTATTCTGCA-3′ SB337R 5′-AATTCGGTCTGCTTCTTCTG-3′ | 487 |
| ST5 | SB336F 5′-GTGGGTAGAGGAAGGAAAACA-3′ SB336R 5′-AGAACAAGTCGATGAAGTGAGAT-3′ | 317 |
| ST6 | SB332F 5′-GCATCCAGACTACTATCAACATT-3′ SB332R 5′-CCATTTTCAGACAACCACTTA-3′ | 338 |
| ST7 | SB155F 5′-ATCAGCCTACAATCTCCTC-3′ SB155R 5′-ATCGCCACTTCTCCAAT-3′ | 650 |
| Group | Concentration (mg/mL) * | After 24 h | After 48 h | After 72 h | ||||
|---|---|---|---|---|---|---|---|---|
| Number ** ± SD | %Inhibition | Number ** ± SD | %Inhibition | Number ** ± SD | %Inhibition | p *** | ||
| Control | 0.00 | 13.25 ± 4.99 | 0.00 | 40.25 ± 13.89 | 0.00 | 64.50 ± 20.40 | 0.00 a | 0.001 |
| Metronidazole | 0.05 | 0.00 ± 0.00 | 100.00 a | 0.00 ± 0.00 | 100.00 | 0.00 ± 0.00 | 100.00 g | |
| MRS broth medium | 0.55 | 13.67 ± 3.79 | 0.00 | 42.33 ± 3.06 | 0.00 | 64.50 ± 6.03 | 0.00 a | |
| 1.09 | 13.00 ± 4.58 | 1.88 | 41.30 ± 4.93 | 0.00 | 67.00 ± 8.89 | 0.00 a | ||
| 2.19 | 14.00 ± 2.00 | 0.00 | 40.67± 3.06 | 0.00 | 68.33 ± 8.50 | 0.00 a | ||
| 4.38 | 12.67 ± 4.04 | 4.38 | 38.33 ± 3.51 | 4.77 | 61.67 ± 11.93 | 4.38 a | ||
| 8.75 | 12.33 ± 3.21 | 6.94 | 36.67 ± 3.79 | 8.89 | 60.33 ± 8.02 | 6.46 a | ||
| 13.13 | 11.67 ± 2.08 | 11.92 | 35.67 ± 3,06 | 11.37 | 55.00 ± 6.24 | 14.73 b | ||
| Postbiotic | 0.55 | 8.88 ± 2.65 | 33.02 | 26.00 ± 1.75 | 35.40 | 39.80 ±10.80 | 38.37 c | |
| 1.09 | 5.92 ± 2.25 | 55.35 | 18.50 ± 4.75 | 54.04 | 10.40 ± 2.20 | 83.85 d | ||
| 2.19 | 3.25 ± 0.75 | 75.47 | 6.42 ± 3.59 | 84.06 | 8.3 ± 1.1 | 87.21 e | ||
| 4.38 | 0.75 ± 0.25 | 94.34 | 2.08 ± 0.58 | 94.82 | 3.5 ± 0.5 | 94.57 f | ||
| 8.75 | 0.00 ± 0.00 | 100.00 a | 0.00 ± 0.00 | 100.00 | 0.00 ± 0.00 | 100.00 g | ||
| 13.13 | 0.00 ± 0.00 | 100.00 a | 0.00 ± 0.00 | 100.00 | 0.00 ± 0.00 | 100.00 g | ||
| Concentration (mg/mL) | pH Value | p * |
|---|---|---|
| 0 (Control) | 7.43 ± 0.11 a | 0.001 |
| 0.55 | 7.23 ± 0.07 a | |
| 1.09 | 7.28 ± 0.05 a | |
| 2.19 | 7.23 ± 0.08 a | |
| 4.38 | 7.09 ± 0.03 a | |
| 8.75 | 6.26 ± 0.1 b | |
| 13.13 | 5.71 ± 0.1 c |
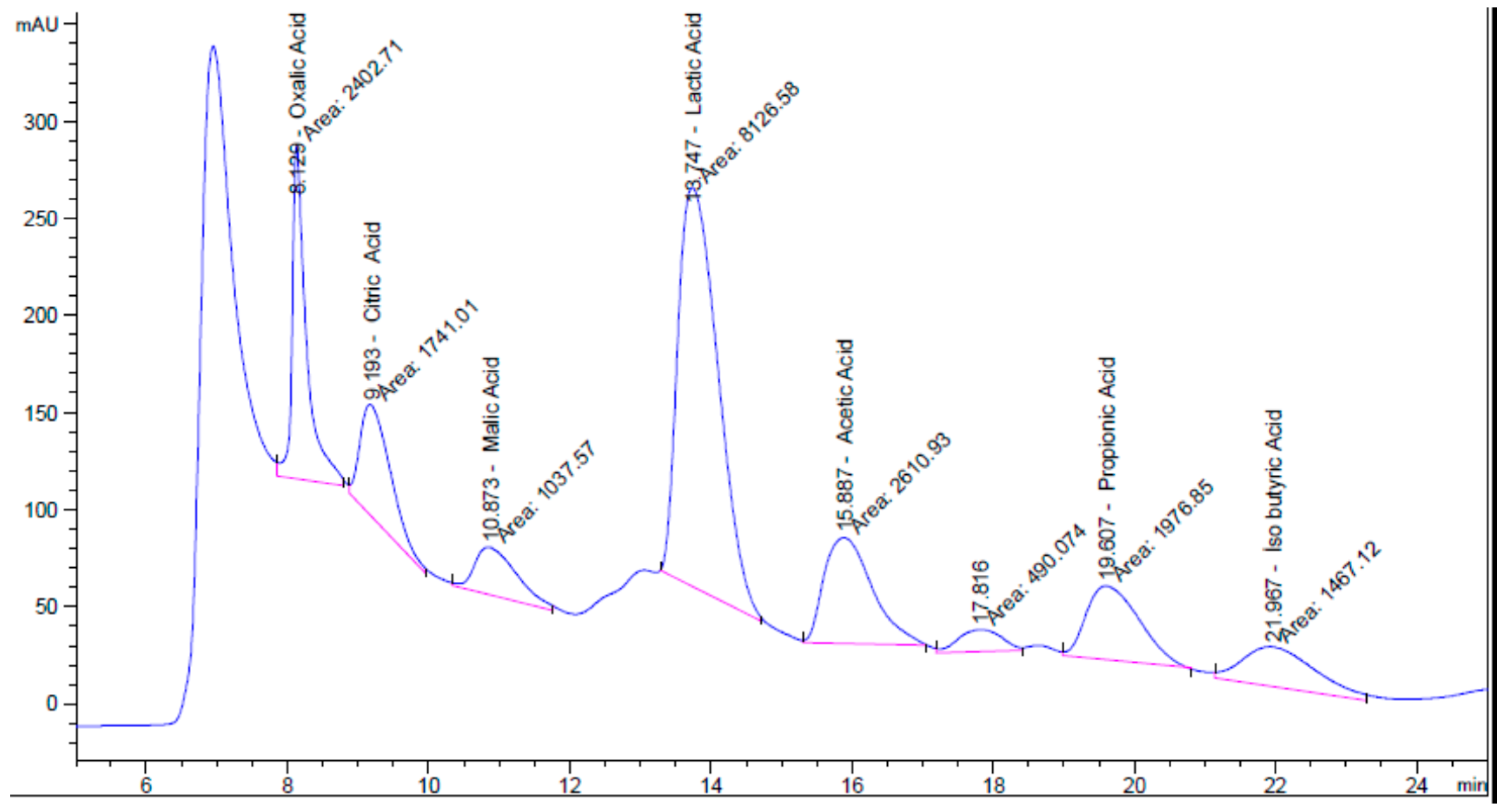
| Organic Acid | Retention Time (min) | Peak Area (mAU·s) | Concentration (ng/µL) |
|---|---|---|---|
| Oxalic Acid | 8.129 | 2402.71 | 188.95 |
| Citric Acid | 9.193 | 1741.01 | 2102.46 |
| Malic Acid | 10.873 | 1037.57 | 1414.62 |
| Lactic Acid | 13.747 | 8126.58 | 14,040.20 |
| Acetic Acid | 15.887 | 2610.93 | 10,050.10 |
| Propionic Acid | 19.607 | 1976.85 | 529.55 |
| Isobutyric Acid | 21.967 | 1467.12 | 321.87 |
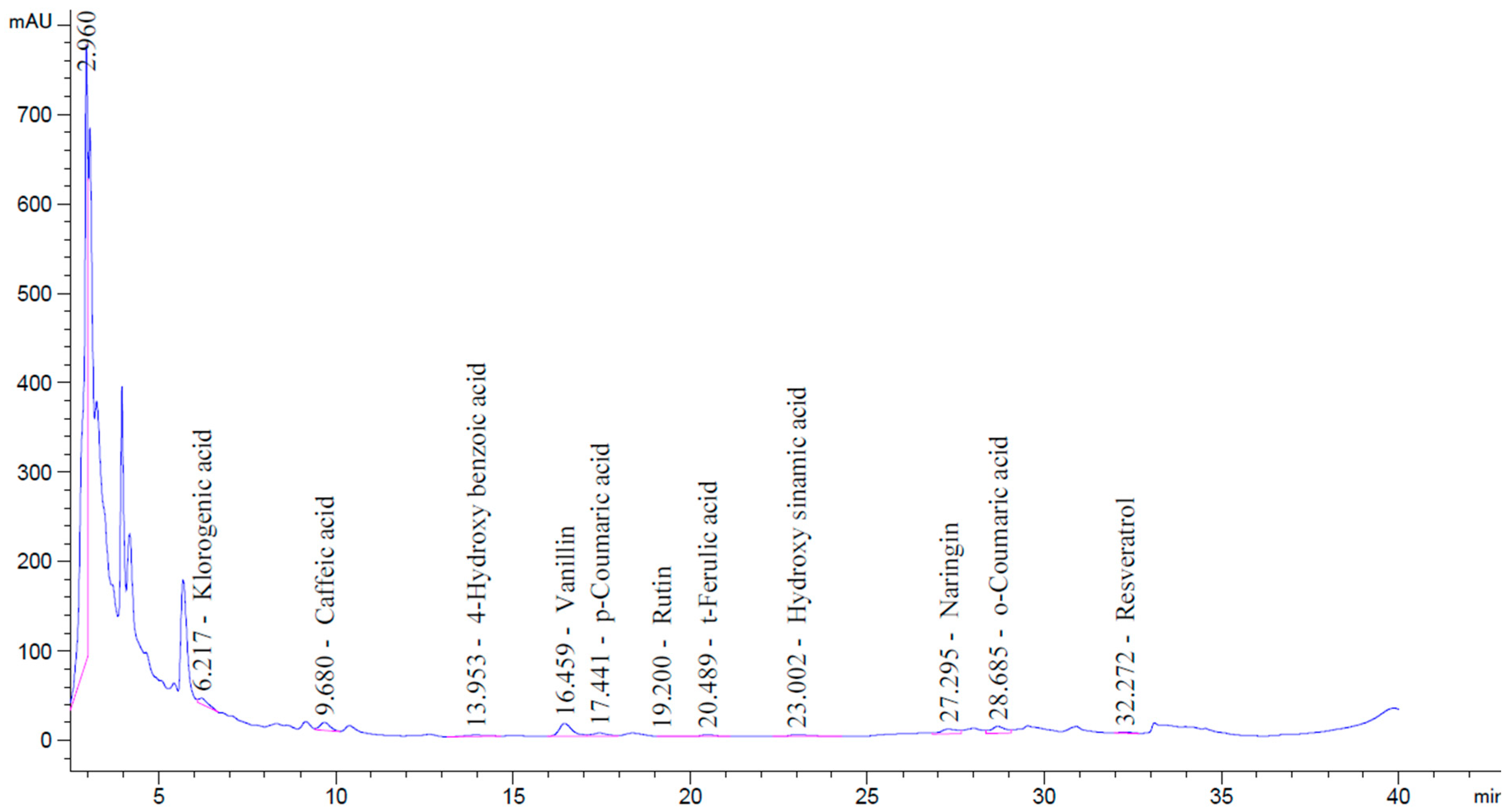
| Compound Name | Retention Time (min) | Peak Area (mAU·s) | Concentration (ng/µL) |
|---|---|---|---|
| Chlorogenic Acid | 6.217 | 137.621 | 8.515 |
| Caffeic Acid | 9.680 | 177.898 | 5.024 |
| 4-Hydroxy benzoic Acid | 13.953 | 76.895 | 2.561 |
| Vanillin | 16.459 | 370.830 | 12.85 |
| p-Coumaric Acid | 17.441 | 116.670 | 3.095 |
| Rutin | 19.200 | 10.501 | 0.298 |
| t-Ferulic Acid | 20.489 | 52.024 | 1.577 |
| Hydroxycinnamic Acid | 23.002 | 72.441 | 1.686 |
| Naringin | 27.295 | 194.189 | 19.465 |
| o-Coumaric Acid | 28.685 | 212.727 | 5.030 |
| Resveratrol | 32.272 | 26.048 | 0.666 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aydemir, S.; Arvas, Y.E.; Aydemir, M.E.; Barlık, F.; Gürbüz, E.; Yazgan, Y.; Ekici, A. Antiprotozoal Effects of Pediococcus acidilactici-Derived Postbiotic on Blastocystis Subtypes ST1/ST3. Pathogens 2025, 14, 664. https://doi.org/10.3390/pathogens14070664
Aydemir S, Arvas YE, Aydemir ME, Barlık F, Gürbüz E, Yazgan Y, Ekici A. Antiprotozoal Effects of Pediococcus acidilactici-Derived Postbiotic on Blastocystis Subtypes ST1/ST3. Pathogens. 2025; 14(7):664. https://doi.org/10.3390/pathogens14070664
Chicago/Turabian StyleAydemir, Selahattin, Yunus Emre Arvas, Mehmet Emin Aydemir, Fethi Barlık, Esra Gürbüz, Yener Yazgan, and Abdurrahman Ekici. 2025. "Antiprotozoal Effects of Pediococcus acidilactici-Derived Postbiotic on Blastocystis Subtypes ST1/ST3" Pathogens 14, no. 7: 664. https://doi.org/10.3390/pathogens14070664
APA StyleAydemir, S., Arvas, Y. E., Aydemir, M. E., Barlık, F., Gürbüz, E., Yazgan, Y., & Ekici, A. (2025). Antiprotozoal Effects of Pediococcus acidilactici-Derived Postbiotic on Blastocystis Subtypes ST1/ST3. Pathogens, 14(7), 664. https://doi.org/10.3390/pathogens14070664