
First Molecular Identification of Cotylophoron cotylophorum in Ecuador and Its Phylogenetic Relationship with Fasciola hepatica
 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
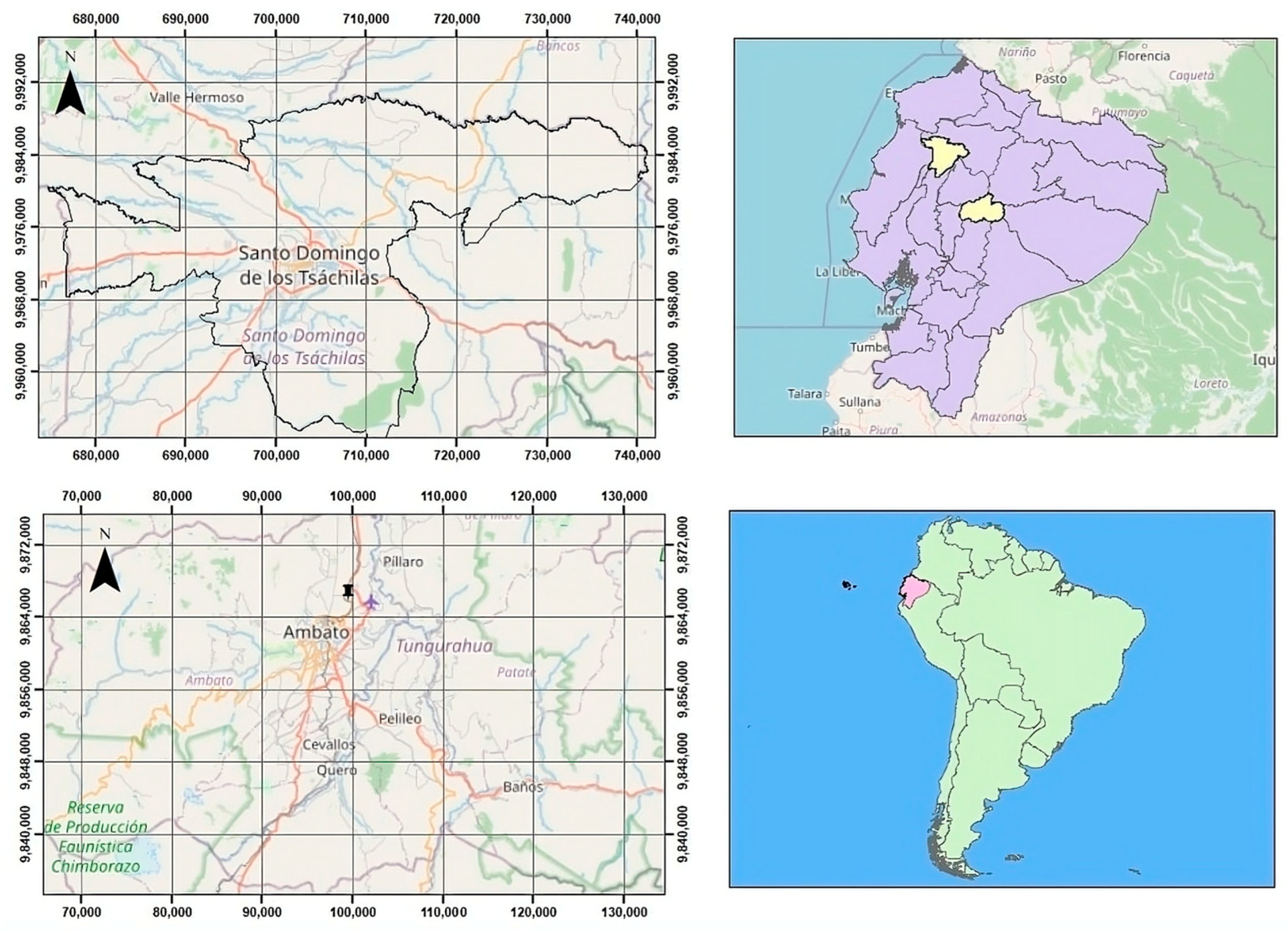
2.1. Sample Collection
2.2. DNA Extraction and Polymerase Chain Reaction (PCR)
2.3. Sequence and Phylogenetic Analysis
3. Results
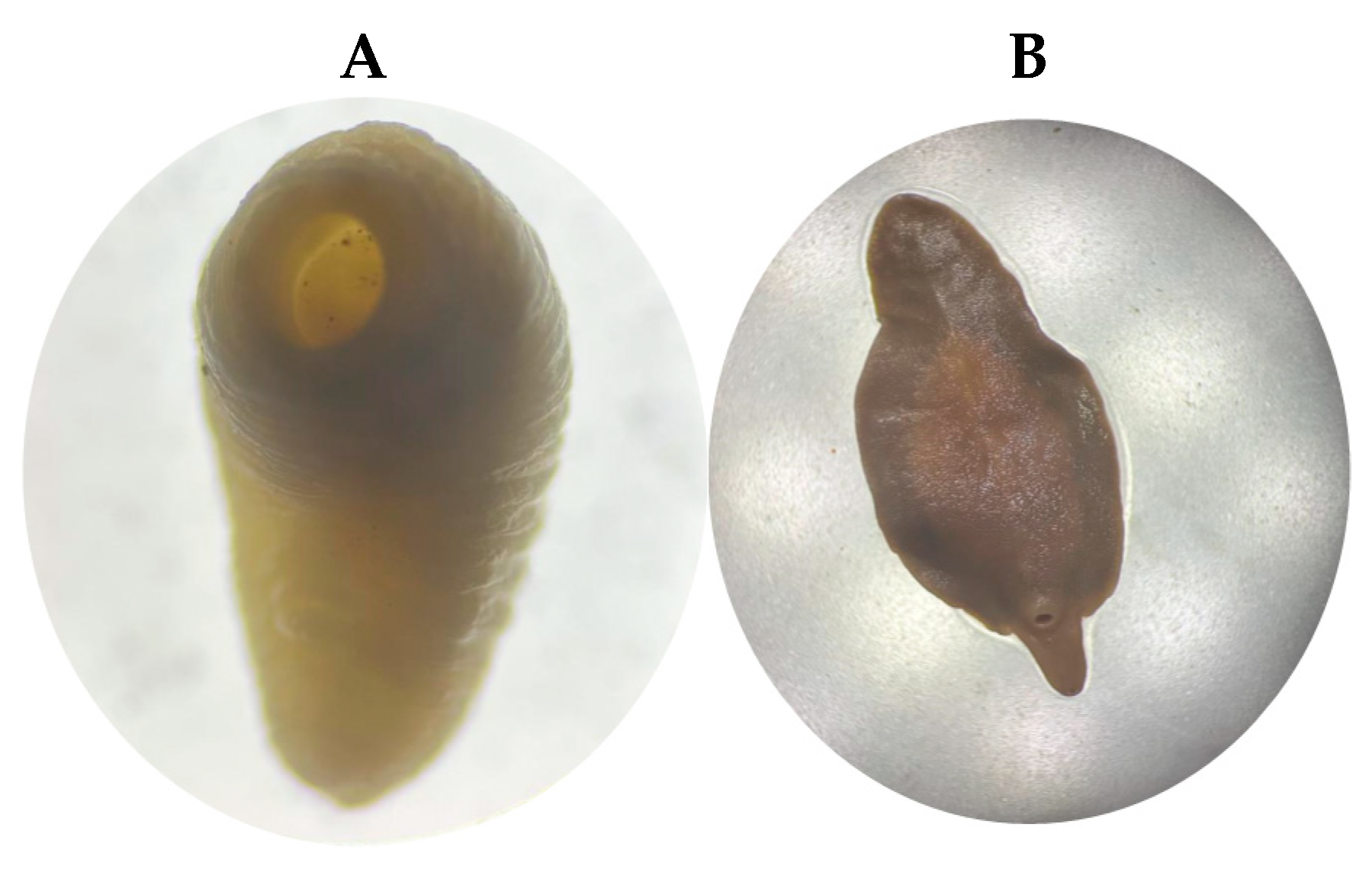
3.1. Specimens Studied
3.2. PCR Detection
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taipe, M.V.T.; Guambi, L.A.D.; Solorzano, J.A.S.; Hidrovo, C.A.M.; Tito, Z.L.; de la Cueva, F.I.C.; Méndez, J.A.A. Realidades de la ganadería bovina en la provincia de Manabí. Cienc. Lat. Rev. Científica Multidiscip. 2022, 6, 311–338. [Google Scholar] [CrossRef]
- Andrade, R.B.; Forero, J.; Ospina, J. Prevalence of Trematodes and Host Risk Factors Associated with Dairy Cattle in Colombia. Rev. Vet. 2021, 31, 171–177. [Google Scholar] [CrossRef]
- Angulo Cubillán, F.; Molero, M.; Escalona, F.; Muñoz, J.; Ramírez Barrios, R. Prevalencia y dinámica de hpg mensual de fasciola hepatica y otros helmintos en un rebaño bovino de una zona inundable tropical. Rev. Científica 2007, 17, 111–116. [Google Scholar]
- Torrel, S.; Rojas-Moncada, J.; Saldaña, K.; Silva, M.; Gallardo, I.; del P. Cadenillas, R.; Alfaro, D.; Irigoín, C.; Murga-Moreno, C.A.; Vargas-Rocha, L.; et al. Trematodos Del Ganado Vacuno Lechero al Pastoreo En Cajamarca, Perú: Fasciola Hepatica y Calicophoron Microbothrioides. Rev. Investig. Vet. Perú 2023, 34, e24296. [Google Scholar] [CrossRef]
- Longa, A.; Traviezo-Valles, L.; Perdomo, R. Primer caso humano parasitado por Paramphistomidae (Trematoda: Digenea) en Venezuela. Boletín Malariol. Salud Ambient. 2010, 50, 301–306. [Google Scholar]
- Mas-Coma, S.; Valero, M.A.; Bargues, M.D. Chapter 2. Fasciola, Lymnaeids and Human Fascioliasis, with a Global Overview on Disease Transmission, Epidemiology, Evolutionary Genetics, Molecular Epidemiology and Control. Adv. Parasitol. 2009, 69, 41–146. [Google Scholar] [CrossRef]
- Bargues, M.D.; Artigas, P.; Khoubbane, M.; Flores, R.; Glöer, P.; Rojas-García, R.; Ashrafi, K.; Falkner, G.; Mas-Coma, S. Lymnaea Schirazensis, an Overlooked Snail Distorting Fascioliasis Data: Genotype, Phenotype, Ecology, Worldwide Spread, Susceptibility, Applicability. PLoS ONE 2011, 6, e24567. [Google Scholar] [CrossRef]
- Caron, Y.; Celi-Erazo, M.; Hurtrez-Boussès, S.; Lounnas, M.; Pointier, J.-P.; Saegerman, C.; Losson, B.; Benítez-Ortíz, W. Is Galba schirazensis (Mollusca, Gastropoda) an Intermediate Host of Fasciola hepatica (Trematoda, Digenea) in Ecuador? Parasite 2017, 24, 24. [Google Scholar] [CrossRef]
- Villavicencio, A.F.; Bargues, M.D.; Artigas, P.; Guamán, R.; Ulloa, S.M.; Romero, J.; Osca, D.; Mas-Coma, S. Lymnaeid Snail Vectors of Fascioliasis, Including the First Finding of Lymnaea neotropica in Ecuador, Assessed by Ribosomal DNA Sequencing in the Southern Zone Close to the Peru Border. Acta Parasitol. 2019, 64, 839–849. [Google Scholar] [CrossRef]
- Cauquil, L.; Hüe, T.; Hurlin, J.-C.; Mitchell, G.; Searle, K.; Skuce, P.; Zadoks, R. Prevalence and Sequence-Based Identity of Rumen Fluke in Cattle and Deer in New Caledonia. PLoS ONE 2016, 11, e0152603. [Google Scholar] [CrossRef]
- Ghatani, S.; Shylla, J.A.; Roy, B.; Tandon, V. Multilocus Sequence Evaluation for Differentiating Species of the Trematode Family Gastrothylacidae, with a Note on the Utility of Mitochondrial COI Motifs in Species Identification. Gene 2014, 548, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Zintl, A.; Garcia-Campos, A.; Trudgett, A.; Chryssafidis, A.L.; Talavera-Arce, S.; Fu, Y.; Egan, S.; Lawlor, A.; Negredo, C.; Brennan, G.; et al. Bovine Paramphistomes in Ireland. Vet. Parasitol. 2014, 204, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Itagaki, T.; Tsumagari, N.; Tsutsumi, K.; Chinone, S. Discrimination of Three Amphistome Species by PCR-RFLP Based on rDNA ITS2 Markers. J. Vet. Med. Sci. 2003, 65, 931–933. [Google Scholar] [CrossRef]
- Reddy, P.V.; Subramanyan, S. Chromosome Studies in the Liver Fluke, Fasciola Gigantica Cobbold, 1856 from Andhra Pradesh. Curr. Sci. 1973, 42, 288–291. [Google Scholar]
- Sakaguchi, Y.; Nakagawa, C. A Note on the Chromosomes of the Common Liver Fluke (Fasciola sp.) from Japan. Chromosome Inf. Serv. 1975, 19, 20–21. [Google Scholar]
- Sakaguchi; Yoneda, A. Further Chromosome Study of the Common Liver Fluke (Fasciola sp.) in Japan. Chromosome Inf. Serv. 1976, 20, 25–26. [Google Scholar]
- Terasaki, K.; Noda, Y.; Shibahara, T.; Itagaki, T. Morphological Comparisons and Hypotheses on the Origin of Polyploids in Parthenogenetic Fasciola sp. J. Parasitol. 2000, 86, 724–729. [Google Scholar] [CrossRef]
- Itagaki, T.; Kikawa, M.; Sakaguchi, K.; Shimo, J.; Terasaki, K.; Shibahara, T.; Fukuda, K. Genetic Characterization of Parthenogenic Fasciola Sp. in Japan on the Basis of the Sequences of Ribosomal and Mitochondrial DNA. Parasitology 2005, 131, 679–685. [Google Scholar] [CrossRef]
- de S. Mariante, A.; Egipto, A. Animal genetic resources in Brazil: Result of five centuries of natural selection. Theriogenology 2002, 57, 223–235. [Google Scholar] [CrossRef]
- Colli, L.; Milanesi, M.; Vajana, E.; Iamartino, D.; Bomba, L.; Puglisi, F.; Del Corvo, M.; Nicolazzi, E.L.; Ahmed, S.S.E.; Herrera, J.R.V.; et al. New Insights on Water Buffalo Genomic Diversity and Post-Domestication Migration Routes From Medium Density SNP Chip Data. Front. Genet. 2018, 9, 53. [Google Scholar] [CrossRef]
- Kasahara, S.; Ohari, Y.; Jin, S.; Calvopina, M.; Takagi, H.; Sugiyama, H.; Itagaki, T. Molecular Characterization Revealed Fasciola Specimens in Ecuador Are All Fasciola hepatica, None at All of Fasciola Gigantica or Parthenogenic Fasciola Species. Parasitol. Int 2021, 80, 102215. [Google Scholar] [CrossRef] [PubMed]
- Celi-Erazo, M.; Alda, P.; Montenegro-Franco, M.; Pavon, D.; Minda-Aluisa, E.; Calvopiña, M.; Pointier, J.P.; Hurtrez-Boussès, S.; Cevallos, W.; Benítez-Ortíz, W.; et al. Prevalence of Fasciola Hepatica Infection in Galba Cousini and Galba Schirazensis from an Andean Region of Ecuador. Vet. Parasitol. Reg. Stud. Rep. 2020, 20, 100390. [Google Scholar] [CrossRef]
- Lounnas, M.; Vázquez, A.A.; Alda, P.; Sartori, K.; Pointier, J.-P.; David, P.; Hurtrez-Boussès, S. Isolation, Characterization and Population-Genetic Analysis of Microsatellite Loci in the Freshwater Snail Galba Cubensis (Lymnaeidae). J. Molluscan Stud. 2017, 83, 63–68. [Google Scholar] [CrossRef]
- Paraense, W.L. Lymnaea Viatrix and Lymnaea Columella in the Neotropical Region: A Distributional Outline. Mem. Inst. Oswaldo Cruz 1982, 77, 181–188. [Google Scholar] [CrossRef]
- Paraense, W.L. Lymnaea Cousini Jousseaume, 1887, from Ecuador (Gastropoda: Lymnaeidae). Mem. Inst. Oswaldo Cruz 1995, 90, 605–609. [Google Scholar] [CrossRef]
- Paraense, W.L. Planorbidae, Lymnaeidae and Physidae of Ecuador (Mollusca: Basommatophora). Mem. Inst. Oswaldo Cruz 2004, 99, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Thang, T.N.; Hakim, H.; Rahimi, R.R.; Ichikawa-Seki, M. Molecular Analysis Reveals Expansion of Fasciola Hepatica Distribution from Afghanistan to China. Parasitol. Int. 2019, 72, 101930. [Google Scholar] [CrossRef]
- Mitchell, G.; Zadoks, R.N.; Skuce, P.J. A Universal Approach to Molecular Identification of Rumen Fluke Species Across Hosts, Continents, and Sample Types. Front. Vet. Sci. 2020, 7, 605259. [Google Scholar] [CrossRef]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular Evolution and Phylogenetic Inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef]
- Alasaad, S.; Huang, C.Q.; Li, Q.Y.; Granados, J.E.; García-Romero, C.; Pérez, J.M.; Zhu, X.Q. Characterization of Fasciola Samples from Different Host Species and Geographical Localities in Spain by Sequences of Internal Transcribed Spacers of rDNA. Parasitol. Res. 2007, 101, 1245–1250. [Google Scholar] [CrossRef]
- Dowling, T.E.; Moritz, C.; Palmer, J.D. Nucleic Acids II: Restriction Site Analysis. In Molecular Systematics; Sinauer: Sunderland, MA, USA, 1990; pp. 250–317. [Google Scholar]
- Jones, R.A.; Williams, H.W.; Dalesman, S.; Brophy, P.M. Confirmation of Galba Truncatula as an Intermediate Host Snail for Calicophoron Daubneyi in Great Britain, with Evidence of Alternative Snail Species Hosting Fasciola Hepatica. Parasites Vectors 2015, 8, 656. [Google Scholar] [CrossRef] [PubMed]
- Abrous, M.; Rondelaud, D.; Dreyfuss, G.; Cabaret, J. Unusual Transmission of the Liver Fluke, Fasciola Hepatica, by Lymnaea Glabra or Planorbis Leucostoma in France. J. Parasitol. 1998, 84, 1257–1259. [Google Scholar] [CrossRef] [PubMed]
- Bowman, D.D. Georgis Parasitología Para Veterinarios, 9th ed.; Elsevier: Barcelona, Spain, 2011; ISBN 978-84-8086-705-4. [Google Scholar]
- Szmidt-Adjidé, V.; Abrous, M.; Adjidé, C.C.; Dreyfuss, G.; Lecompte, A.; Cabaret, J.; Rondelaud, D. Prevalence of Paramphistomum daubneyi Infection in Cattle in Central France. Vet. Parasitol. 2000, 87, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Dinnik, J.A. Paramphistomum daubneyi sp.nov. from Cattle and Its Snail Host in the Kenya Highlands. Parasitology 1962, 52, 143–151. [Google Scholar] [CrossRef]
- Bargues, M.D.; Valero, M.A.; Trueba, G.A.; Fornasini, M.; Villavicencio, A.F.; Guamán, R.; De Elías-Escribano, A.; Pérez-Crespo, I.; Artigas, P.; Mas-Coma, S. DNA Multi-Marker Genotyping and CIAS Morphometric Phenotyping of Fasciola gigantica-Sized Flukes from Ecuador, with an Analysis of the Radix Absence in the New World and the Evolutionary Lymnaeid Snail Vector Filter. Animals 2021, 11, 2495. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Access Codes ITS2 | Species | Location | Variation Position (147) |
|---|---|---|---|
| PV473765 | Cotylophoron cotylophorum | Ecuador | Y |
| JX678257.1 | Cotylophoron cotylophorum | India | C |
| LN610457.1 | Calicophoron daubneyi | Algeria | C |
| AB973394.1 | Calicophoron daubneyi | Ireland | C |
| GU133057.1 | Calicophoron calicophorum | India | C |
| JX678259.1 | Calicophoron calicophorum | India | C |
| AB042188.1 | Calicophoron calicophorum | Japan | C |
| KR337554.1 | Calicophoron microbothrioides | China | C |
| KF791030.1 | Calicophoron microbothrioides | Peru | C |
| HM209064.1 | Paramphistomum leydeni | Argentina | C |
| AB973398.1 | Paramphistomum leydeni | Ireland | C |
| KX274233.1 | Paramphistomum cervi | Croatia | C |
| HM026462.1 | Paramphistomum cervi | Slovakia | C |
| KF642983.1 | Paramphistomum epiclitum | India | C |
| KX840345.1 | Paramphistomum epiclitum | India | C |
| Access Codes ITS2 | Species | Location | Variation Position (212) |
|---|---|---|---|
| PV473766 | Fasciola hepatica | Ecuador | Y |
| MN970007.1 | Fasciola hepatica | Australia | C |
| AJ557567.1 | Fasciola hepatica | France | C |
| AJ557568.1 | Fasciola hepatica | China | C |
| OQ689976.1 | Fascioloides magna | Romania | C |
| KU232369.1 | Fascioloides magna | United States | C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barragán-López, G.; Bedoya-Páez, F.; Lugo-Almarza, M.; Fonseca-Restrepo, C.; Angulo-Cubillán, F.; Romero, E.J.; de Waard, J.H.; Reyna-Bello, A. First Molecular Identification of Cotylophoron cotylophorum in Ecuador and Its Phylogenetic Relationship with Fasciola hepatica. Pathogens 2025, 14, 659. https://doi.org/10.3390/pathogens14070659
Barragán-López G, Bedoya-Páez F, Lugo-Almarza M, Fonseca-Restrepo C, Angulo-Cubillán F, Romero EJ, de Waard JH, Reyna-Bello A. First Molecular Identification of Cotylophoron cotylophorum in Ecuador and Its Phylogenetic Relationship with Fasciola hepatica. Pathogens. 2025; 14(7):659. https://doi.org/10.3390/pathogens14070659
Chicago/Turabian StyleBarragán-López, Geanella, Fausto Bedoya-Páez, María Lugo-Almarza, Carolina Fonseca-Restrepo, Francisco Angulo-Cubillán, Edison J. Romero, Jacobus H. de Waard, and Armando Reyna-Bello. 2025. "First Molecular Identification of Cotylophoron cotylophorum in Ecuador and Its Phylogenetic Relationship with Fasciola hepatica" Pathogens 14, no. 7: 659. https://doi.org/10.3390/pathogens14070659
APA StyleBarragán-López, G., Bedoya-Páez, F., Lugo-Almarza, M., Fonseca-Restrepo, C., Angulo-Cubillán, F., Romero, E. J., de Waard, J. H., & Reyna-Bello, A. (2025). First Molecular Identification of Cotylophoron cotylophorum in Ecuador and Its Phylogenetic Relationship with Fasciola hepatica. Pathogens, 14(7), 659. https://doi.org/10.3390/pathogens14070659