
Molecular Detection and Characterization of Tick-Borne Pathogens in Ixodes ricinus Ticks Collected from Humans

 , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Tick Collection
2.2. Morphological Identification of Ticks
2.3. Sample Preparation and DNA Extraction
2.4. Molecular Analysis
2.4.1. Real-Time PCR
2.4.2. Nested PCR for Molecular Identification of Borrelia spp.
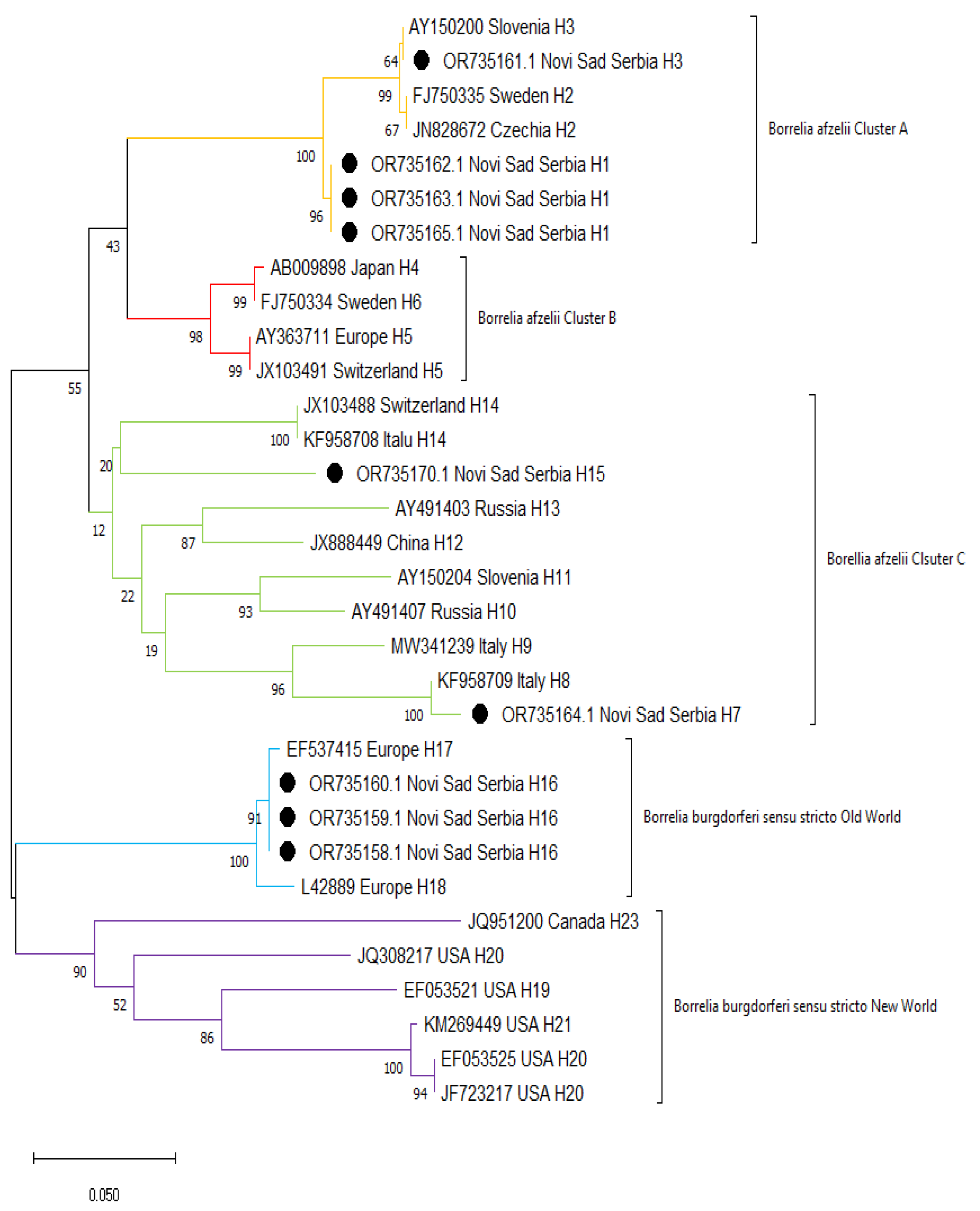
2.5. Molecular Typing of Borrelia spp.
2.6. Phylogenetic Analysis
2.7. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sonenshine, D.E. Range Expansion of Tick Disease Vectors in North America: Implications for Spread of Tick-Borne Disease. Int. J. Environ. Res. Public Health 2018, 15, 478. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, J.; Antunes, S.; Bonnet, S.; Cabezas-Cruz, A.; Domingos, A.G.; Estrada-Peña, A.; Johnson, N.; Kocan, K.M.; Mansfield, K.L.; Nijhof, A.M.; et al. Tick-Pathogen Interactions and Vector Competence: Identification of Molecular Drivers for Tick-Borne Diseases. Front. Cell. Infect. Microbiol. 2017, 7, 114. [Google Scholar] [CrossRef] [PubMed]
- Migné, C.V.; Hönig, V.; Bonnet, S.I.; Palus, M.; Rakotobe, S.; Galon, C.; Heckmann, A.; Vyletova, E.; Devillers, E.; Attoui, H.; et al. Evaluation of two artificial infection methods of live ticks as tools for studying interactions between tick-borne viruses and their tick vectors. Sci. Rep. 2022, 12, 491. [Google Scholar] [CrossRef]
- Potkonjak, A.; Gutiérrez, R.; Savić, S.; Vračar, V.; Nachum-Biala, Y.; Jurišić, A.; Kleinerman, G.; Rojas, A.; Petrović, A.; Baneth, G.; et al. Molecular detection of emerging tick-borne pathogens in Vojvodina, Serbia. Ticks Tick-Borne Dis. 2016, 7, 199–203. [Google Scholar] [CrossRef]
- Michael, L. Focus on Common Small Animal Vector-Borne Diseases in Central and Southeastern Europe. Acta Vet. Sciendo 2020, 70, 147–169. [Google Scholar] [CrossRef]
- Potkonjak, A.; Zekic-Stosic, M. Tick-borne infections of dogs in Serbia: A review of research. Vet. Glas. 2020, 74, 107–124. [Google Scholar] [CrossRef]
- Ivanović, I.; Stošić, M.Ž.; Sabljić, E.R.; Kišek, T.C.; Špik, V.C.; Popović, A.; Savić, S. Ecology and prevalence of Borrelia burgdorferi s.l. in Ixodes ricinus (Acari: Ixodidae) ticks. Acta Vet. Hung. 2022, 70, 15–23. [Google Scholar] [CrossRef]
- Audino, T.; Pautasso, A.; Bellavia, V.; Carta, V.; Ferrari, A.; Verna, F.; Grattarola, C.; Iulini, B.; Pintore, M.D.; Bardelli, M.; et al. Ticks infesting humans and associated pathogens: A cross-sectional study in a 3-year period (2017–2019) in northwest Italy. Parasites Vectors 2021, 14, 136. [Google Scholar] [CrossRef]
- Banović, P.; Piloto-Sardiñas, E.; Mijatović, D.; Foucault-Simonin, A.; Simin, V.; Bogdan, I.; Obregón, D.; Mateos-Hernández, L.; Moutailler, S.; Cabezas-Cruz, A. Differential detection of tick-borne pathogens in human platelets and whole blood using microfluidic PCR. Acta Trop. 2023, 238, 106756. [Google Scholar] [CrossRef]
- Gray, J.S.; Dautel, H.; Estrada-Peña, A.; Kahl, O.; Lindgren, E. Effects of climate change on ticks and tick-borne diseases in Europe. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 593232. [Google Scholar] [CrossRef]
- Korotkov, Y.; Kozlova, T.; Kozlovskaya, L. Observations on changes in abundance of questing Ixodes ricinus, castor bean tick, over a 35-year period in the eastern part of its range (Russia, Tula region). Med. Vet. Èntomol. 2015, 29, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F. Climate change, biodiversity, ticks and tick-borne diseases: The butterfly effect. Int. J. Parasitol. Parasites Wildl. 2015, 4, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Voyiatzaki, C.; Papailia, S.I.; Venetikou, M.S.; Pouris, J.; Tsoumani, M.E.; Papageorgiou, E.G. Climate Changes Exacerbate the Spread of Ixodes ricinus and the Occurrence of Lyme Borreliosis and Tick-Borne Encephalitis in Europe—How Climate Models Are Used as a Risk Assessment Approach for Tick-Borne Diseases. Int. J. Environ. Res. Public. Heal. 2022, 19, 6516. [Google Scholar] [CrossRef]
- Tomanović, S.; Radulović, Ž.; Masuzawa, T.; Milutinović, M. Coexistence of emerging bacterial pathogens in Ixodes ricinusticks in Serbia. Parasite 2010, 17, 211–217. [Google Scholar] [CrossRef]
- Savić, S.; Vidić, B.; Lazić, S.; Lako, B.; Potkonjak, A.; Lepšanović, Z. Borrelia burgdorferi in ticks and dogs in the province of Vojvodina, Serbia. Parasite 2010, 17, 357–361. [Google Scholar] [CrossRef]
- Potkonjak, A.; Kleinerman, G.; Gutiérrez, R.; Savić, S.; Vračar, V.; Nachum-Biala, Y.; Jurišić, A.; Rojas, A.; Petrović, A.; Ivanović, I.; et al. Occurrence of Borrelia burgdorferi Sensu Lato in Ixodes ricinus Ticks with First Identification of Borrelia miyamotoi in Vojvodina, Serbia. Vector-Borne Zoonotic Dis. 2016, 16, 631–635. [Google Scholar] [CrossRef]
- Banović, P.; Díaz-Sánchez, A.A.; Galon, C.; Simin, V.; Mijatović, D.; Obregón, D.; Moutailler, S.; Cabezas-Cruz, A. Humans infested with Ixodes ricinus are exposed to a diverse array of tick-borne pathogens in Serbia. Ticks Tick Borne Dis. 2021, 12, 101609. [Google Scholar] [CrossRef]
- Banović, P.; Díaz-Sánchez, A.A.; Simin, V.; Foucault-Simonin, A.; Galon, C.; Wu-Chuang, A.; Mijatović, D.; Obregón, D.; Moutailler, S.; Cabezas-Cruz, A. Clinical Aspects and Detection of Emerging Rickettsial Pathogens: A “One Health” Approach Study in Serbia, 2020. Front. Microbiol. 2022, 12, 797399. [Google Scholar] [CrossRef] [PubMed]
- Tomanović, S.; Chochlakis, D.; Radulović, Ž.; Milutinović, M.; Ćakić, S.; Mihaljica, D.; Tselentis, Y.; Psaroulaki, A. Analysis of pathogen co-occurrence in host-seeking adult hard ticks from Serbia. Exp. Appl. Acarol. 2013, 59, 367–376. [Google Scholar] [CrossRef]
- Debeljak, Z.; Medić, S.; Baralić, M.; Andrić, A.; Tomić, A.; Vidanović, D.; Šekler, M.; Matović, K.; Vasković, N. Clinical, epidemiological and epizootic features of a Q fever outbreak in the border region between Serbia and Montenegro. J. Infect. Dev. Ctries. 2018, 12, 290–296. [Google Scholar] [CrossRef]
- Jovanović, V. Annual Report on Infectious Diseases in the Republic of Serbia for 2021. Belgrade. 2021. Available online: https://www.batut.org.rs/index.php?content=2523 (accessed on 10 March 2025).
- Filipović, M.M.K.; Beletić, A.D.; Božović, A.V.I.; Milanović, Z.; Tyrrell, P.; Buch, J.; Breitschwerdt, E.B.; Birkenheuer, A.J.; Chandrashekar, R. Molecular and Serological Prevalence of Anaplasma phagocytophilum, A. platys, Ehrlichia canis, E. chaffeenses, E. ewingii, Borrelia burgdorferi, Babesia canis, B. gibsoni and B. vogeli among Clinically Healthy Outdoor Dogs in Serbia. Vet. Parasitol. Reg. Stud. Rep. 2018, 14, 117–122. [Google Scholar] [CrossRef]
- Vasić, A.; Nieder, M.; Zdravković, N.; Bojkovski, J.; Bugarski, D.; Pavlović, I.; Silaghi, C. Tick infestation and occurrence of Anaplasma phagocytophilum and piroplasms in cattle in the Republic of Serbia. Parasitol. Res. 2018, 117, 1813–1818. [Google Scholar] [CrossRef] [PubMed]
- Zutic, J.; Vojinovic, D.; Stanojevic, S.; Kureljusic, B.; Milicevic, V.; Kureljusic, J.; Spalevic, L. Seroprevalence of Coxiella burnetii in cattle in the Belgrade epizootiological area. Biotechnol. Anim. Husb. 2020, 36, 359–369. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Bouattour, A.J.L.C.; Camicas, J.L.; Walker, A.R. Ticks of Domestic Animals in the Mediterranean Region. In A Guide to Identification of Species; University of Zaragoza: Zaragoza, Spain, 2004. [Google Scholar]
- Wang, I.-N.; E Dykhuizen, D.; Qiu, W.; Dunn, J.J.; Bosler, E.M.; Luft, B.J. Genetic Diversity of ospC in a Local Population of Borrelia burgdorferi sensu stricto. Genetics 1999, 151, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Pearson, P.; Skaltsis, O.; Luo, C.-Y.; Xu, G.; Oppler, Z.; Brisson, D.; Rich, S.M. A Borrelia burgdorferi outer surface protein C (OspC) genotyping method using Luminex technology. PLoS ONE 2022, 17, e0269266. [Google Scholar] [CrossRef] [PubMed]
- Staden, R.; Beal, K.F.; Bonfield, J.K. The Staden Package. In Bioinformatics Methods and Protocols; Humana Press: Totowa, NJ, USA, 1998; pp. 115–130. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Milutinović, M.; Masuzawa, T.; Tomanović, S.; Radulović, Ž.; Fukui, T.; Okamoto, Y. Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum, Francisella tularensis and their co-infections in host-seeking Ixodes ricinus ticks collected in Serbia. Exp. Appl. Acarol. 2008, 45, 171–183. [Google Scholar] [CrossRef]
- Bogunović, D.; Stević, N.; Sidi-Boumedine, K.; Mišić, D.; Tomanović, S.; Kulišić, Z.; Magaš, V.; Radojičić, S. Molecular Evidence of Q Fever Agent Coxiella Burnetii in Ixodid Ticks Collected from Stray Dogs in Belgrade (Serbia). Acta Vet. Brno. 2018, 68, 257–268. [Google Scholar] [CrossRef]
- Banović, P.; Čapo, I.; Ogorelica, D.; Vranješ, N.; Simin, V.; Lalošević, D. Mysterious path of Borrelia spielmanii: Spreading without morphological alteration of collagen type I and IV. Futur. Microbiol. 2019, 14, 1469–1475. [Google Scholar] [CrossRef]
- Simin, V.; Lalosevic, D.; Mijatovic, D.; Tomanovic, S.; Miljevic, M.; Cabrilo, B.; Bogdan, I.; Banovic, P. Borellia burgdorferi infection in removed ticks and anti-borrelia antibodies in infested patients admitted to the Pasteur institute, Novi Sad. Vet. Glas. 2020, 74, 164–177. [Google Scholar] [CrossRef]
- Banovic, P. Early-Stage Diagnosis and Risk Factors for Tick Infestation and Development of Lyme Borreliosis in Residents of South Bačka District. Ph.D. Thesis, University of Novi Sad, Novi Sad, Serbia, 2022. [Google Scholar]
- Anderson, M.L. Infectious causes of bovine abortion during mid- to late-gestation. Theriogenology 2007, 68, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Kalmár, Z.; Dumitrache, M.O.; D’amico, G.; Matei, I.A.; Ionică, A.M.; Gherman, C.M.; Lupșe, M.; Mihalca, A.D. Multiple Tick-Borne Pathogens in Ixodes ricinus Ticks Collected from Humans in Romania. Pathogens 2020, 9, 390. [Google Scholar] [CrossRef] [PubMed]
- Maiwald, M.; Oehme, R.; March, O.; Petney, T.N.; Kimmig, P.; Naser, K.; Zappe, H.A.; Hassler, D.; Doeberitz, M.V.K. Transmission risk of Borrelia burgdorferi sensu lato from Ixodes ricinus ticks to humans in southwest Germany. Epidemiol. Infect. 1998, 121, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Lasić, L.; Ušanović, L.; Ćakić, S.; Hanjalić, J.; Stroil, B.K. First molecular detection of Borrelia burgdorferi in Ixodes ricinu ticks collected from humans in the Sarajevo Canton. Syst. Appl. Acarol. 2020, 25, 169–172. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
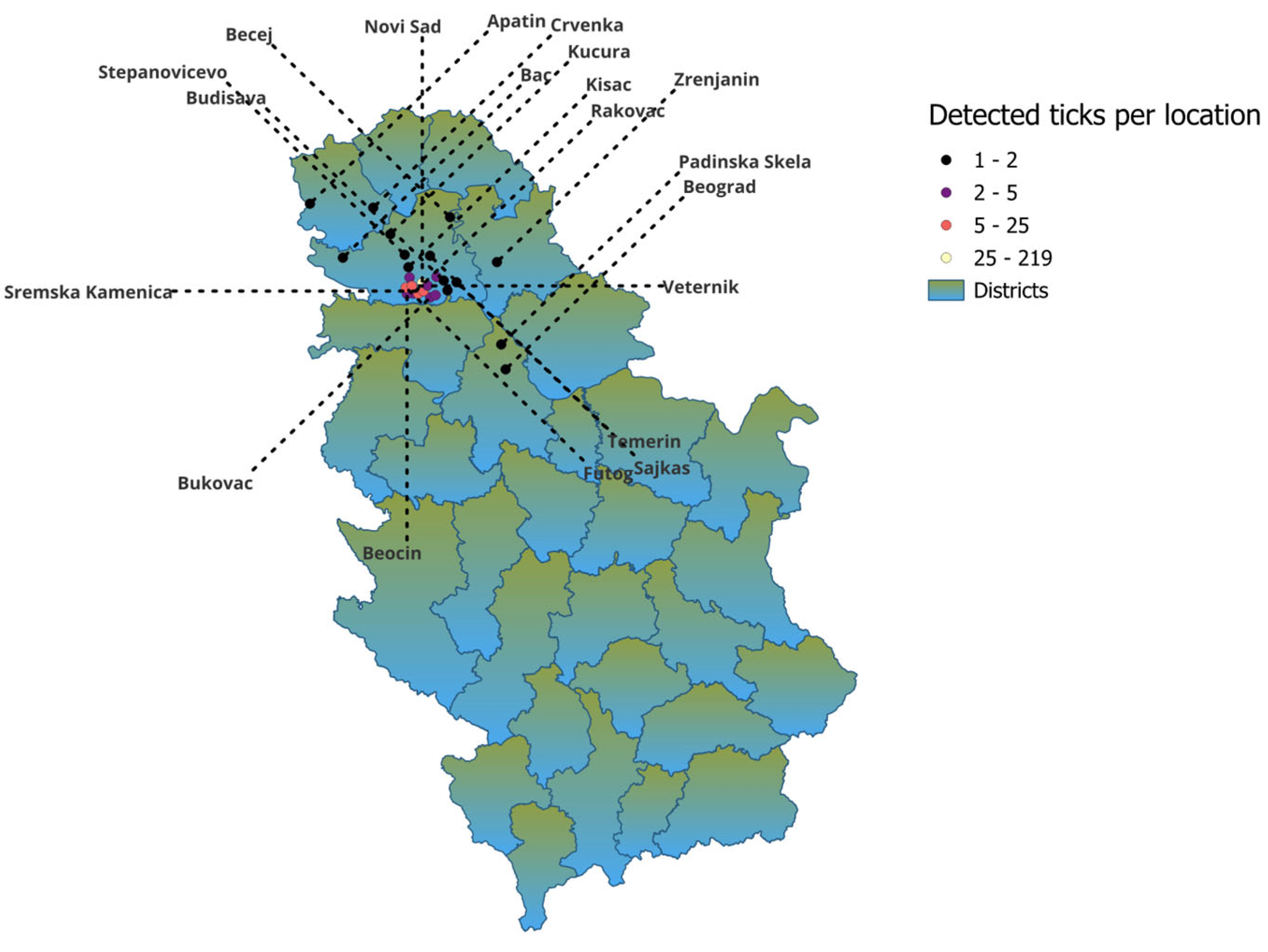
| City | Number of Ticks |
|---|---|
| Novi Sad | 219 |
| Temerin | 1 |
| Kac | 4 |
| Rakovac | 4 |
| Ledinci | 6 |
| Beočin | 4 |
| Bukovac | 3 |
| Sremska kamenica | 25 |
| Stepanovicevo | 1 |
| Futog | 7 |
| Becej | 1 |
| Rumenka | 4 |
| Budisava | 1 |
| Sremski Karlovci | 3 |
| Beograd | 1 |
| Kovilj | 2 |
| Apatin | 1 |
| Veternik | 8 |
| Crvenka | 1 |
| Kisač | 2 |
| Bač | 1 |
| Kucura | 1 |
| Šajkaš | 1 |
| Zrenjanin | 1 |
| Padinska skela | 1 |
| Petrovaradin | 5 |
| Total | 308 |
| Primer | Position in ospC | Sequence (5′–3′) | Tm |
|---|---|---|---|
| OC6 (+) | 6 | AAA GAA TAC ATT AAG TGC GAT ATT | 54 °C |
| OC623 (−) | 623 | TTA AGG TTT TTT TTG GAC TTT CTG C | 54 °C |
| Year | Development Stage | Borrelia Positive (n/Tested) | Borrelia Prevalence Per Year (%) | Anaplasma Positive (n/Tested) | Anaplasma Prevalence Per Year (%) |
|---|---|---|---|---|---|
| 2019 | Nymph | 2/14 | 10.34 | 1/14 | 6.89 |
| Adult Female | 1/15 | 1/15 | |||
| 2020 | Nymph | 5/24 | 24.6 | 0/24 | 3.28 |
| Adult Female | 10/37 | 2/37 | |||
| 2021 | Nymph | 1/18 | 5.55 | 0/18 | 1.85 |
| Adult Female | 2/36 | 1/36 | |||
| 2022 | Nymph | 2/24 | 10.41 | 0/24 | 0 |
| Adult Female | 3/24 | 0/24 | |||
| 2023 | Nymph | 4/11 | 32.26 | 0/11 | 0 |
| Adult Female | 6/20 | 0/20 | |||
| 2024 | Nymph | 4/20 | 23.53 | 0/20 | 0 |
| Adult Female | 16/65 | 0/65 | |||
| Total | Nymph | 19/111 | 20.45 | 1/111 | 1.62 |
| Adult Female | 44/197 | 4/197 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Žekić, M.; Kišek, T.C.; Cvitković-Špik, V.; Ružić-Sabljić, E.; Gajdov, V.; Potkonjak, A.; Jurišić, A.; Savić, S. Molecular Detection and Characterization of Tick-Borne Pathogens in Ixodes ricinus Ticks Collected from Humans. Pathogens 2025, 14, 528. https://doi.org/10.3390/pathogens14060528
Žekić M, Kišek TC, Cvitković-Špik V, Ružić-Sabljić E, Gajdov V, Potkonjak A, Jurišić A, Savić S. Molecular Detection and Characterization of Tick-Borne Pathogens in Ixodes ricinus Ticks Collected from Humans. Pathogens. 2025; 14(6):528. https://doi.org/10.3390/pathogens14060528
Chicago/Turabian StyleŽekić, Marina, Tjaša Cerar Kišek, Vesna Cvitković-Špik, Eva Ružić-Sabljić, Vladimir Gajdov, Aleksandar Potkonjak, Aleksandar Jurišić, and Sara Savić. 2025. "Molecular Detection and Characterization of Tick-Borne Pathogens in Ixodes ricinus Ticks Collected from Humans" Pathogens 14, no. 6: 528. https://doi.org/10.3390/pathogens14060528
APA StyleŽekić, M., Kišek, T. C., Cvitković-Špik, V., Ružić-Sabljić, E., Gajdov, V., Potkonjak, A., Jurišić, A., & Savić, S. (2025). Molecular Detection and Characterization of Tick-Borne Pathogens in Ixodes ricinus Ticks Collected from Humans. Pathogens, 14(6), 528. https://doi.org/10.3390/pathogens14060528