
Viperin: A Multifunctional Protein in Antiviral Immunity and Disease Pathogenesis


Abstract
1. Introduction
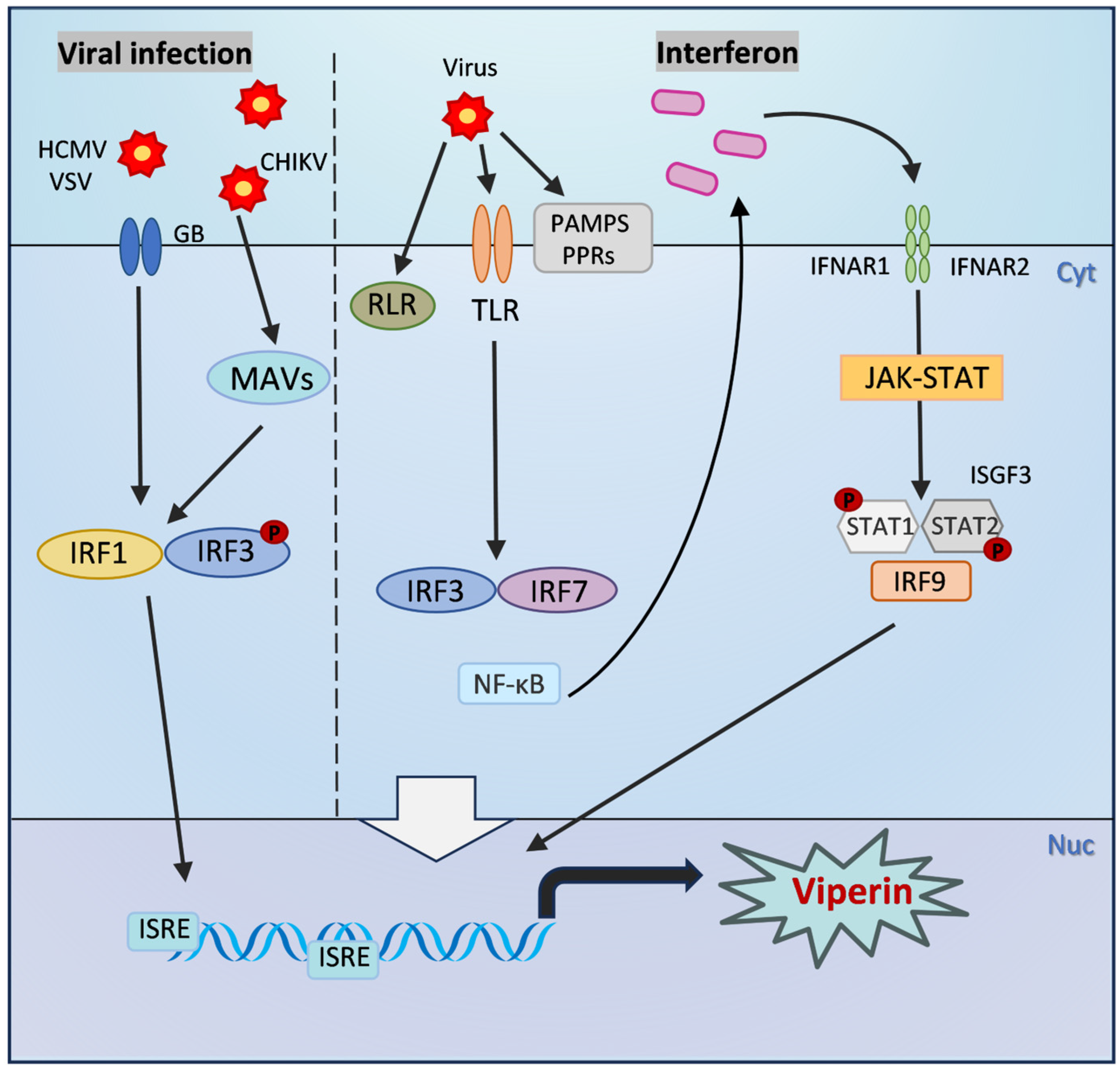
2. Structure and Expression Regulation of Viperin
3. Mechanism of Antiviral Action of Viperin
3.1. Direct Antiviral Effects of Viperin
3.2. Indirect Antiviral Effects of Viperin
4. Role of Viperin in Disease
4.1. Viperin in Diseases Associated with Viral Infections
4.2. Potential Function of Viperin in Autoimmune Diseases
4.3. The Role of Viperin in Immunometabolic Reprogramming of Tumors
5. Conclusions and Perspectives
Funding
Conflicts of Interest
Abbreviations
| DENV | Dengue virus |
| ZIKV | Zika virus |
| TBEV | Tick-borne encephalitis virus |
| WNV | West Nile virus |
| HCV | Hepatitis C virus |
| HIV | Human immunodeficiency virus |
| BUNV | Bunyamwera virus |
| EIAV | Equine infectious anemia virus |
| RABV | Rabies virus |
| SVA | Senecavirus A |
| IAV | Influenza A virus |
References
- Stertz, S.; Hale, B.G. Interferon system deficiencies exacerbating severe pandemic virus infections. Trends Microbiol. 2021, 29, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type I interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-stimulated genes: A complex web of host defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.E.; Hughes, J.; Gu, Q.; Behdenna, A.; Singer, J.B.; Dennis, T.; Orton, R.J.; Varela, M.; Gifford, R.J.; Wilson, S.J.; et al. Fundamental properties of the mammalian innate immune system revealed by multispecies comparison of type I interferon responses. PLoS Biol. 2017, 15, e2004086. [Google Scholar] [CrossRef]
- Schoggins, J.W.; Wilson, S.J.; Panis, M.; Murphy, M.Y.; Jones, C.T.; Bieniasz, P.; Rice, C.M. A diverse range of gene products are effectors of the type I interferon antiviral response. Nature 2011, 472, 481–485. [Google Scholar] [CrossRef]
- Chiang, H.S.; Liu, H.M. The Molecular Basis of Viral Inhibition of IRF- and STAT-Dependent Immune Responses. Front. Immunol. 2018, 9, 3086. [Google Scholar] [CrossRef]
- Savan, R.; Gale, M., Jr. Innate immunity and interferon in SARS-CoV-2 infection outcome. Immunity 2023, 56, 1443–1450. [Google Scholar] [CrossRef]
- Karki, R.; Kanneganti, T.D. Innate immunity, cytokine storm, and inflammatory cell death in COVID-19. J. Transl. Med. 2022, 20, 542. [Google Scholar] [CrossRef]
- Ramasamy, S.; Subbian, S. Critical Determinants of Cytokine Storm and Type I Interferon Response in COVID-19 Pathogenesis. Clin. Microbiol. Rev. 2021, 34, e00299-20. [Google Scholar] [CrossRef]
- Chin, K.C.; Cresswell, P. Viperin (cig5), an IFN-inducible antiviral protein directly induced by human cytomegalovirus. Proc. Natl. Acad. Sci. USA 2001, 98, 15125–15130. [Google Scholar] [CrossRef]
- Nasr, N.; Maddocks, S.; Turville, S.G.; Harman, A.N.; Woolger, N.; Helbig, K.J.; Wilkinson, J.; Bye, C.R.; Wright, T.K.; Rambukwelle, D.; et al. HIV-1 infection of human macrophages directly induces viperin which inhibits viral production. Blood 2012, 120, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Szretter, K.J.; Brien, J.D.; Thackray, L.B.; Virgin, H.W.; Cresswell, P.; Diamond, M.S. The Interferon-Inducible Gene viperin Restricts West Nile Virus Pathogenesis. J. Virol. 2011, 85, 11557–11566. [Google Scholar] [CrossRef] [PubMed]
- Van der Hoek, K.H.; Eyre, N.S.; Shue, B.; Khantisitthiporn, O.; Glab-Ampi, K.; Carr, J.M.; Gartner, M.J.; Jolly, L.A.; Thomas, P.Q.; Adikusuma, F.; et al. Viperin is an important host restriction factor in control of Zika virus infection. Sci. Rep. 2017, 7, 4475. [Google Scholar] [CrossRef]
- Wang, X.; Hinson, E.R.; Cresswell, P. The Interferon-Inducible Protein Viperin Inhibits Influenza Virus Release by Perturbing Lipid Rafts. Cell Host Microbe 2007, 2, 96–105. [Google Scholar] [CrossRef]
- Gizzi, A.S.; Grove, T.L.; Arnold, J.J.; Jose, J.; Jangra, R.K.; Garforth, S.J.; Du, Q.; Cahill, S.M.; Dulyaninova, N.G.; Love, J.D.; et al. A naturally occurring antiviral ribonucleotide encoded by the human genome. Nature 2018, 558, 610–614. [Google Scholar] [CrossRef]
- Pan, M.; Yin, Y.; Hu, T.; Wang, X.; Jia, T.; Sun, J.; Wang, Q.; Meng, W.; Zhu, J.; Dai, C.; et al. UXT attenuates the CGAS-STING1 signaling by targeting STING1 for autophagic degradation. Autophagy 2023, 19, 440–456. [Google Scholar] [CrossRef]
- Fitzgerald, K.A. The interferon inducible gene: Viperin. J. Interferon Cytokine Res. 2011, 31, 131–135. [Google Scholar] [CrossRef]
- Helbig, K.J.; Beard, M.R. The Role of Viperin in the Innate Antiviral Response. J. Mol. Biol. 2014, 426, 1210–1219. [Google Scholar] [CrossRef]
- Seo, J.-Y.; Yaneva, R.; Cresswell, P. Viperin: A Multifunctional, Interferon-Inducible Protein that Regulates Virus Replication. Cell Host Microbe 2011, 10, 534–539. [Google Scholar] [CrossRef]
- Hinson, E.R.; Cresswell, P. The antiviral protein, viperin, localizes to lipid droplets via its N-terminal amphipathic alpha-helix. Proc. Natl. Acad. Sci. USA 2009, 106, 20452–20457. [Google Scholar] [CrossRef]
- Teng, T.-S.; Foo, S.-S.; Simamarta, D.; Lum, F.-M.; Teo, T.-H.; Lulla, A.; Yeo, N.K.W.; Koh, E.G.L.; Chow, A.; Leo, Y.-S.; et al. Viperin restricts chikungunya virus replication and pathology. J. Clin. Investig. 2012, 122, 4447–4460. [Google Scholar] [CrossRef] [PubMed]
- Hinson, E.R.; Cresswell, P. The N-terminal amphipathic alpha-helix of viperin mediates localization to the cytosolic face of the endoplasmic reticulum and inhibits protein secretion. J. Biol. Chem. 2009, 284, 4705–4712. [Google Scholar] [CrossRef]
- Tang, Y.-D.; Na, L.; Zhu, C.-H.; Shen, N.; Yang, F.; Fu, X.-Q.; Wang, Y.-H.; Fu, L.-H.; Wang, J.-Y.; Lin, Y.-Z.; et al. Equine Viperin Restricts Equine Infectious Anemia Virus Replication by Inhibiting the Production and/or Release of Viral Gag, Env, and Receptor via Distortion of the Endoplasmic Reticulum. J. Virol. 2014, 88, 12296–12310. [Google Scholar] [CrossRef]
- Grunkemeyer, T.J.; Ghosh, S.; Patel, A.M.; Sajja, K.; Windak, J.; Basrur, V.; Kim, Y.; Nesvizhskii, A.I.; Kennedy, R.T.; Marsh, E.N.G. The antiviral enzyme viperin inhibits cholesterol biosynthesis. J. Biol. Chem. 2021, 297, 100824. [Google Scholar] [CrossRef]
- Fenwick, M.K.; Li, Y.; Cresswell, P.; Modis, Y.; Ealick, S.E. Structural studies of viperin, an antiviral radical SAM enzyme. Proc. Natl. Acad. Sci. USA 2017, 114, 6806–6811. [Google Scholar] [CrossRef]
- Carlton-Smith, C.; Elliott, R.M. Viperin, MTAP44, and Protein Kinase R Contribute to the Interferon-Induced Inhibition of Bunyamwera Orthobunyavirus Replication. J. Virol. 2012, 86, 11548–11557. [Google Scholar] [CrossRef]
- Jiang, D.; Guo, H.; Xu, C.; Chang, J.; Gu, B.; Wang, L.; Block, T.M.; Guo, J.T. Identification of three interferon-inducible cellular enzymes that inhibit the replication of hepatitis C virus. J. Virol. 2008, 82, 1665–1678. [Google Scholar] [CrossRef]
- Jiang, D.; Weidner, J.M.; Qing, M.; Pan, X.B.; Guo, H.; Xu, C.; Zhang, X.; Birk, A.; Chang, J.; Shi, P.Y.; et al. Identification of five interferon-induced cellular proteins that inhibit west nile virus and dengue virus infections. J. Virol. 2010, 84, 8332–8341. [Google Scholar] [CrossRef]
- Upadhyay, A.S.; Vonderstein, K.; Pichlmair, A.; Stehling, O.; Bennett, K.L.; Dobler, G.; Guo, J.-T.; Superti-Furga, G.; Lill, R.; Överby, A.K.; et al. Viperin is an iron-sulfur protein that inhibits genome synthesis of tick-borne encephalitis virus via radical SAM domain activity. Cell. Microbiol. 2014, 16, 834–848. [Google Scholar] [CrossRef]
- Fink, J.; Gu, F.; Ling, L.; Tolfvenstam, T.; Olfat, F.; Chin, K.C.; Aw, P.; George, J.; Kuznetsov, V.A.; Schreiber, M.; et al. Host gene expression profiling of dengue virus infection in cell lines and patients. PLoS Negl. Trop. Dis. 2007, 1, e86. [Google Scholar] [CrossRef]
- Fenwick, M.K.; Su, D.; Dong, M.; Lin, H.; Ealick, S.E. Structural Basis of the Substrate Selectivity of Viperin. Biochemistry 2020, 59, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.M.; Marsh, E.N.G. The Antiviral Enzyme, Viperin, Activates Protein Ubiquitination by the E3 Ubiquitin Ligase, TRAF6. J. Am. Chem. Soc. 2021, 143, 4910–4914. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Feng, L.; Chen, P.; Li, A.; Guo, S.; Jiao, X.; Zhang, C.; Zhao, Y.; Jin, X.; Zhong, K.; et al. Viperin inhibits classical swine fever virus replication by interacting with viral nonstructural 5A protein. J. Med. Virol. 2020, 92, 149–160. [Google Scholar] [CrossRef]
- Hsu, J.C.; Laurent-Rolle, M.; Pawlak, J.B.; Xia, H.; Kunte, A.; Hee, J.S.; Lim, J.; Harris, L.D.; Wood, J.M.; Evans, G.B.; et al. Viperin triggers ribosome collision-dependent translation inhibition to restrict viral replication. Mol. Cell 2022, 82, 1631–1642.e6. [Google Scholar] [CrossRef]
- Wang, W.; Xu, L.; Su, J.; Peppelenbosch, M.P.; Pan, Q. Transcriptional Regulation of Antiviral Interferon-Stimulated Genes. Trends Microbiol. 2017, 25, 573–584. [Google Scholar] [CrossRef]
- Liu, D.; Zheng, H.; Li, Y.; Zhou, P.; Jin, H.; Luo, R. Molecular cloning and functional characterization of duck Janus kinase 1. Mol. Immunol. 2020, 117, 29–36. [Google Scholar] [CrossRef]
- Giugliano, S.; Kriss, M.; Golden-Mason, L.; Dobrinskikh, E.; Stone, A.E.; Soto-Gutierrez, A.; Mitchell, A.; Khetani, S.R.; Yamane, D.; Stoddard, M.; et al. Hepatitis C virus infection induces autocrine interferon signaling by human liver endothelial cells and release of exosomes, which inhibits viral replication. Gastroenterology 2015, 148, 392–402.e13. [Google Scholar] [CrossRef]
- Chmielewski, S.; Olejnik, A.; Sikorski, K.; Pelisek, J.; Błaszczyk, K.; Aoqui, C.; Nowicka, H.; Zernecke, A.; Heemann, U.; Wesoly, J.; et al. STAT1-dependent signal integration between IFNγ and TLR4 in vascular cells reflect pro-atherogenic responses in human atherosclerosis. PLoS ONE 2014, 9, e113318. [Google Scholar] [CrossRef]
- Olofsson, P.S.; Jatta, K.; Wågsäter, D.; Gredmark, S.; Hedin, U.; Paulsson-Berne, G.; Söderberg-Nauclér, C.; Hansson, G.K.; Sirsjö, A. The antiviral cytomegalovirus inducible gene 5/viperin is expressed in atherosclerosis and regulated by proinflammatory agents. Arterioscler. Thromb. Vasc. Biol. 2005, 25, e113–e116. [Google Scholar] [CrossRef]
- Yuan, Y.; Miao, Y.; Qian, L.; Zhang, Y.; Liu, C.; Liu, J.; Zuo, Y.; Feng, Q.; Guo, T.; Zhang, L.; et al. Targeting UBE4A Revives Viperin Protein in Epithelium to Enhance Host Antiviral Defense. Mol. Cell 2020, 77, 734–747.e7. [Google Scholar] [CrossRef]
- Yuan, Y.; Miao, Y.; Ren, T.; Huang, F.; Qian, L.; Chen, X.; Zuo, Y.; Zhang, H.G.; He, J.; Qiao, C.; et al. High salt activates p97 to reduce host antiviral immunity by restricting Viperin induction. EMBO Rep. 2021, 23, e53466. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Qian, L.; Miao, Y.; Cui, Q.; Cao, T.; Yu, Y.; Zhang, T.; Zhao, Q.; Zhang, R.; Ren, T.; et al. Targeting Viperin prevents coxsackievirus B3-induced acute heart failure. Cell Discov. 2025, 11, 34. [Google Scholar] [CrossRef]
- Tang, H.B.; Lu, Z.L.; Wei, X.K.; Zhong, T.Z.; Zhong, Y.Z.; Ouyang, L.X.; Luo, Y.; Xing, X.W.; Liao, F.; Peng, K.K.; et al. Viperin inhibits rabies virus replication via reduced cholesterol and sphingomyelin and is regulated upstream by TLR4. Sci. Rep. 2016, 6, 30529. [Google Scholar] [CrossRef]
- Honda, K.; Takaoka, A.; Taniguchi, T. Type I interferon [corrected] gene induction by the interferon regulatory factor family of transcription factors. Immunity 2006, 25, 349–360. [Google Scholar] [CrossRef]
- Panda, D.; Gjinaj, E.; Bachu, M.; Squire, E.; Novatt, H.; Ozato, K.; Rabin, R.L. IRF1 Maintains Optimal Constitutive Expression of Antiviral Genes and Regulates the Early Antiviral Response. Front. Immunol. 2019, 10, 1019. [Google Scholar] [CrossRef]
- Seo, J.Y.; Yaneva, R.; Hinson, E.R.; Cresswell, P. Human cytomegalovirus directly induces the antiviral protein viperin to enhance infectivity. Science 2011, 332, 1093–1097. [Google Scholar] [CrossRef]
- Boudinot, P.; Riffault, S.; Salhi, S.; Carrat, C.; Sedlik, C.; Mahmoudi, N.; Charley, B.; Benmansour, A. Vesicular stomatitis virus and pseudorabies virus induce a vig1/cig5 homologue in mouse dendritic cells via different pathways. J. Gen. Virol. 2000, 81, 2675–2682. [Google Scholar] [CrossRef]
- Ru, J.; Sun, H.; Fan, H.; Wang, C.; Li, Y.; Liu, M.; Tang, H. MiR-23a facilitates the replication of HSV-1 through the suppression of interferon regulatory factor 1. PLoS ONE 2014, 9, e114021. [Google Scholar] [CrossRef]
- White, L.K.; Sali, T.; Alvarado, D.; Gatti, E.; Pierre, P.; Streblow, D.; Defilippis, V.R. Chikungunya virus induces IPS-1-dependent innate immune activation and protein kinase R-independent translational shutoff. J. Virol. 2011, 85, 606–620. [Google Scholar] [CrossRef]
- Khantisitthiporn, O.; Shue, B.; Eyre, N.S.; Nash, C.W.; Turnbull, L.; Whitchurch, C.B.; Van der Hoek, K.H.; Helbig, K.J.; Beard, M.R. Viperin interacts with PEX19 to mediate peroxisomal augmentation of the innate antiviral response. Life Sci. Alliance 2021, 4, e202000915. [Google Scholar] [CrossRef]
- Mattijssen, S.; Pruijn, G.J. Viperin, a key player in the antiviral response. Microbes Infect. 2012, 14, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.-L.; Chang, T.-H.; Liao, C.-L.; Lin, Y.-L. The Cellular Antiviral Protein Viperin Is Attenuated by Proteasome-Mediated Protein Degradation in Japanese Encephalitis Virus-Infected Cells. J. Virol. 2008, 82, 10455–10464. [Google Scholar] [CrossRef] [PubMed]
- Panayiotou, C.; Lindqvist, R.; Kurhade, C.; Vonderstein, K.; Pasto, J.; Edlund, K.; Upadhyay, A.S.; Överby, A.K. Viperin Restricts Zika Virus and Tick-Borne Encephalitis Virus Replication by Targeting NS3 for Proteasomal Degradation. J. Virol. 2018, 92, e02054-17. [Google Scholar] [CrossRef]
- Vanwalscappel, B.; Tada, T.; Landau, N.R. Toll-like receptor agonist R848 blocks Zika virus replication by inducing the antiviral protein viperin. Virology 2018, 522, 199–208. [Google Scholar] [CrossRef]
- Lindqvist, R.; Överby, A.K. The Role of Viperin in Antiflavivirus Responses. DNA Cell Biol. 2018, 37, 725–730. [Google Scholar] [CrossRef]
- Helbig, K.J.; Carr, J.M.; Calvert, J.K.; Wati, S.; Clarke, J.N.; Eyre, N.S.; Narayana, S.K.; Fiches, G.N.; McCartney, E.M.; Beard, M.R. Viperin is induced following dengue virus type-2 (DENV-2) infection and has anti-viral actions requiring the C-terminal end of viperin. PLoS Negl. Trop. Dis. 2013, 7, e2178. [Google Scholar] [CrossRef]
- Chakravarti, A.; Selvadurai, K.; Shahoei, R.; Lee, H.; Fatma, S.; Tajkhorshid, E.; Huang, R.H. Reconstitution and substrate specificity for isopentenyl pyrophosphate of the antiviral radical SAM enzyme viperin. J. Biol. Chem. 2018, 293, 14122–14133. [Google Scholar] [CrossRef]
- Vanwalscappel, B.; Gadea, G.; Desprès, P. A Viperin Mutant Bearing the K358R Substitution Lost its Anti-ZIKA Virus Activity. Int. J. Mol. Sci. 2019, 20, 1574. [Google Scholar] [CrossRef]
- Kim, J.J.; Hong, S.; Seo, J.Y. A Cysteine Residue of Human Cytomegalovirus vMIA Protein Plays a Crucial Role in Viperin Trafficking to Control Viral Infectivity. J. Virol. 2023, 97, e0187422. [Google Scholar] [CrossRef]
- Vonderstein, K.; Nilsson, E.; Hubel, P.; Nygård Skalman, L.; Upadhyay, A.; Pasto, J.; Pichlmair, A.; Lundmark, R.; Överby, A.K.; Pfeiffer, J.K. Viperin Targets Flavivirus Virulence by Inducing Assembly of Noninfectious Capsid Particles. J. Virol. 2018, 92, e01751-17. [Google Scholar] [CrossRef]
- Helbig, K.J.; Eyre, N.S.; Yip, E.; Narayana, S.; Li, K.; Fiches, G.; McCartney, E.M.; Jangra, R.K.; Lemon, S.M.; Beard, M.R. The antiviral protein viperin inhibits hepatitis C virus replication via interaction with nonstructural protein 5A. Hepatology 2011, 54, 1506–1517. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, X.; Pan, T.; Song, W.; Wang, Y.; Zhang, F.; Yuan, Z. Viperin inhibits hepatitis C virus replication by interfering with binding of NS5A to host protein hVAP-33. J. Gen. Virol. 2012, 93, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Wu, Z.; Zeng, P.; Yang, X.; Shi, Y.; Guo, J.; Zhou, J.; Song, J.; Liu, J. RSAD2 suppresses viral replication by interacting with the Senecavirus A 2 C protein. Vet. Res. 2024, 55, 115. [Google Scholar] [CrossRef]
- Khan, K.A.; Marineau, A.; Doyon, P.; Acevedo, M.; Durette, É.; Gingras, A.C.; Servant, M.J. TRK-Fused Gene (TFG), a protein involved in protein secretion pathways, is an essential component of the antiviral innate immune response. PLoS Pathog. 2021, 17, e1009111. [Google Scholar] [CrossRef]
- Kim, J.J.; Kim, K.S.; Eom, J.; Lee, J.B.; Seo, J.Y. Viperin Differentially Induces Interferon-Stimulated Genes in Distinct Cell Types. Immune Netw. 2019, 19, e33. [Google Scholar] [CrossRef]
- Severa, M.; Coccia, E.M.; Fitzgerald, K.A. Toll-like Receptor-dependent and -independent Viperin Gene Expression and Counter-regulation by PRDI-binding Factor-1/BLIMP1. J. Biol. Chem. 2006, 281, 26188–26195. [Google Scholar] [CrossRef]
- Saitoh, T.; Satoh, T.; Yamamoto, N.; Uematsu, S.; Takeuchi, O.; Kawai, T.; Akira, S. Antiviral Protein Viperin Promotes Toll-like Receptor 7- and Toll-like Receptor 9-Mediated Type I Interferon Production in Plasmacytoid Dendritic Cells. Immunity 2011, 34, 352–363. [Google Scholar] [CrossRef]
- Zhu, H.; Zheng, J.; Zhou, Y.; Wu, T.; Zhu, T. Knockdown of RSAD2 attenuates B cell hyperactivity in patients with primary Sjögren’s syndrome (pSS) via suppressing NF-κb signaling pathway. Mol. Cell. Biochem. 2021, 476, 2029–2037. [Google Scholar] [CrossRef]
- Rousset, F. Innate immunity: The bacterial connection. Trends Immunol. 2023, 44, 945–953. [Google Scholar] [CrossRef]
- Li, X.; Dong, Z.; Liu, Y.; Song, W.; Pu, J.; Jiang, G.; Wu, Y.; Liu, L.; Huang, X. A Novel Role for the Regulatory Nod-Like Receptor NLRP12 in Anti-Dengue Virus Response. Front. Immunol. 2021, 12, 744880. [Google Scholar] [CrossRef]
- Yang, S.; Wang, L.; Pan, X.; Liang, Y.; Zhang, Y.; Li, J.; Zhou, B. 5-Methoxyflavone-induced AMPKα activation inhibits NF-κB and P38 MAPK signaling to attenuate influenza A virus-mediated inflammation and lung injury in vitro and in vivo. Cell. Mol. Biol. Lett. 2022, 27, 82. [Google Scholar] [CrossRef] [PubMed]
- Helbig, K.J.; Lau, D.T.Y.; Semendric, L.; Harley, H.A.J.; Beard, M.R. Analysis of ISG Expression in Chronic Hepatitis C Identifies Viperin as a Potential Antiviral Effector. Hepatology 2005, 42, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Liao, Y.; Pei, M.; Qiu, Z.; Liu, Z.; Jin, D.; Zhang, J.; Ma, Z.; Yang, X. RSAD2 Is an Effective Target for High-Yield Vaccine Production in MDCK Cells. Viruses 2022, 14, 2587. [Google Scholar] [CrossRef]
- Seo, J.Y.; Cresswell, P. Viperin regulates cellular lipid metabolism during human cytomegalovirus infection. PLoS Pathog. 2013, 9, e1003497. [Google Scholar] [CrossRef]
- Mantovani, S.; Daga, S.; Fallerini, C.; Baldassarri, M.; Benetti, E.; Picchiotti, N.; Fava, F.; Gallì, A.; Zibellini, S.; Bruttini, M.; et al. Rare variants in Toll-like receptor 7 results in functional impairment and downregulation of cytokine-mediated signaling in COVID-19 patients. Genes Immun. 2022, 23, 51–56. [Google Scholar] [CrossRef]
- Chinthapatla, R.; Sotoudegan, M.; Srivastava, P.; Anderson, T.K.; Moustafa, I.M.; Passow, K.T.; Kennelly, S.A.; Moorthy, R.; Dulin, D.; Feng, J.Y.; et al. Interfering with nucleotide excision by the coronavirus 3′-to-5′ exoribonuclease. Nucleic Acids Res. 2023, 51, 315–336. [Google Scholar] [CrossRef]
- Dong, Z.; Yan, Q.; Cao, W.; Liu, Z.; Wang, X. Identification of key molecules in COVID-19 patients significantly correlated with clinical outcomes by analyzing transcriptomic data. Front. Immunol. 2022, 13, 930866. [Google Scholar] [CrossRef]
- Huang, T.; He, J.; Zhou, X.; Pan, H.; He, F.; Du, A.; Yu, B.; Jiang, N.; Li, X.; Yuan, K.; et al. Discovering common pathogenetic processes between COVID-19 and tuberculosis by bioinformatics and system biology approach. Front. Cell. Infect. Microbiol. 2023, 13, 1280223. [Google Scholar] [CrossRef]
- Fassan, M.; Collesei, A.; Angerilli, V.; Sbaraglia, M.; Fortarezza, F.; Pezzuto, F.; De Gaspari, M.; Businello, G.; Moni, M.; Rizzo, S.; et al. Multi-Design Differential Expression Profiling of COVID-19 Lung Autopsy Specimens Reveals Significantly Deregulated Inflammatory Pathways and SFTPC Impaired Transcription. Cells 2022, 11, 1011. [Google Scholar] [CrossRef]
- Chen, S.; Ye, J.; Lin, Y.; Chen, W.; Huang, S.; Yang, Q.; Qian, H.; Gao, S.; Hua, C. Crucial Roles of RSAD2/viperin in Immunomodulation, Mitochondrial Metabolism and Autoimmune Diseases. Inflammation 2025, 48, 520–540. [Google Scholar] [CrossRef]
- Pisetsky, D.S. Pathogenesis of autoimmune disease. Nature reviews. Nephrology 2023, 19, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Rönnblom, L.; Leonard, D. Interferon pathway in SLE: One key to unlocking the mystery of the disease. Lupus Sci. Med. 2019, 6, e000270. [Google Scholar] [CrossRef]
- Garcia-Romo, G.S.; Caielli, S.; Vega, B.; Connolly, J.; Allantaz, F.; Xu, Z.; Punaro, M.; Baisch, J.; Guiducci, C.; Coffman, R.L.; et al. Netting neutrophils are major inducers of type I IFN production in pediatric systemic lupus erythematosus. Sci. Transl. Med. 2011, 3, 73ra20. [Google Scholar] [CrossRef]
- Deng, Y.; Zheng, Y.; Li, D.; Hong, Q.; Zhang, M.; Li, Q.; Fu, B.; Wu, L.; Wang, X.; Shen, W.; et al. Expression characteristics of interferon-stimulated genes and possible regulatory mechanisms in lupus patients using transcriptomics analyses. EBioMedicine 2021, 70, 103477. [Google Scholar] [CrossRef]
- Nehar-Belaid, D.; Hong, S.; Marches, R.; Chen, G.; Bolisetty, M.; Baisch, J.; Walters, L.; Punaro, M.; Rossi, R.J.; Chung, C.H.; et al. Mapping systemic lupus erythematosus heterogeneity at the single-cell level. Nat. Immunol. 2020, 21, 1094–1106. [Google Scholar] [CrossRef]
- Psarras, A.; Wittmann, M.; Vital, E.M. Emerging concepts of type I interferons in SLE pathogenesis and therapy. Nat. Rev. Rheumatol. 2022, 18, 575–590. [Google Scholar] [CrossRef]
- He, P.; Zhang, Z.; Liao, W.; Xu, D.; Fu, M.; Kang, Y. Screening of gene signatures for rheumatoid arthritis and osteoarthritis based on bioinformatics analysis. Mol. Med. Rep. 2016, 14, 1587–1593. [Google Scholar] [CrossRef]
- Raterman, H.G.; Vosslamber, S.; de Ridder, S.; Nurmohamed, M.T.; Lems, W.F.; Boers, M.; van de Wiel, M.; Dijkmans, B.A.; Verweij, C.L.; Voskuyl, A.E. The interferon type I signature towards prediction of non-response to rituximab in rheumatoid arthritis patients. Arthritis Res. Ther. 2012, 14, R95. [Google Scholar] [CrossRef]
- Lübbers, J.; Brink, M.; van de Stadt, L.A.; Vosslamber, S.; Wesseling, J.G.; van Schaardenburg, D.; Rantapää-Dahlqvist, S.; Verweij, C.L. The type I IFN signature as a biomarker of preclinical rheumatoid arthritis. Ann. Rheum. Dis. 2013, 72, 776–780. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Duan, C.; Xie, C.; Wang, H.; Li, Z.; Li, B.; Wang, T. Identification of key interferon-stimulated genes for indicating the condition of patients with systemic lupus erythematosus. Front. Immunol. 2022, 13, 962393. [Google Scholar] [CrossRef]
- Sezin, T.; Vorobyev, A.; Sadik, C.D.; Zillikens, D.; Gupta, Y.; Ludwig, R.J. Gene Expression Analysis Reveals Novel Shared Gene Signatures and Candidate Molecular Mechanisms between Pemphigus and Systemic Lupus Erythematosus in CD4(+) T Cells. Front. Immunol. 2017, 8, 1992. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Zhao, G.; Ren, D.; Liu, F.; Dong, G.; Hou, Y. Gender differences of B cell signature related to estrogen-induced IFI44L/BAFF in systemic lupus erythematosus. Immunol. Lett. 2017, 181, 71–78. [Google Scholar] [CrossRef]
- Weinstein, A.G.; Godet, I.; Gilkes, D.M. The rise of viperin: The emerging role of viperin in cancer progression. J. Clin. Investig. 2022, 132, e165907. [Google Scholar] [CrossRef]
- Xin, Y.; He, Z.; Mei, Y.; Li, X.; Zhao, Z.; Zhao, M.; Yang, M.; Wu, H. Interferon-α regulates abnormally increased expression of RSAD2 in Th17 and Tfh cells in systemic lupus erythematosus patients. Eur. J. Immunol. 2023, 53, e2350420. [Google Scholar] [CrossRef]
- Morita, M.; Sato, T.; Nomura, M.; Sakamoto, Y.; Inoue, Y.; Tanaka, R.; Ito, S.; Kurosawa, K.; Yamaguchi, K.; Sugiura, Y.; et al. PKM1 Confers Metabolic Advantages and Promotes Cell-Autonomous Tumor Cell Growth. Cancer Cell 2018, 33, 355–367.e7. [Google Scholar] [CrossRef]
- Elstrom, R.L.; Bauer, D.E.; Buzzai, M.; Karnauskas, R.; Harris, M.H.; Plas, D.R.; Zhuang, H.; Cinalli, R.M.; Alavi, A.; Rudin, C.M.; et al. Akt stimulates aerobic glycolysis in cancer cells. Cancer Res. 2004, 64, 3892–3899. [Google Scholar] [CrossRef]
- Isaacs, J.S.; Jung, Y.J.; Mole, D.R.; Lee, S.; Torres-Cabala, C.; Chung, Y.L.; Merino, M.; Trepel, J.; Zbar, B.; Toro, J.; et al. HIF overexpression correlates with biallelic loss of fumarate hydratase in renal cancer: Novel role of fumarate in regulation of HIF stability. Cancer Cell 2005, 8, 143–153. [Google Scholar] [CrossRef]
- Nikiforov, M.A.; Chandriani, S.; O’Connell, B.; Petrenko, O.; Kotenko, I.; Beavis, A.; Sedivy, J.M.; Cole, M.D. A functional screen for Myc-responsive genes reveals serine hydroxymethyltransferase, a major source of the one-carbon unit for cell metabolism. Mol. Cell. Biol. 2002, 22, 5793–5800. [Google Scholar] [CrossRef]
- Shi, L.Z.; Bonner, J.A. Bridging Radiotherapy to Immunotherapy: The IFN-JAK-STAT Axis. Int. J. Mol. Sci. 2021, 22, 12295. [Google Scholar] [CrossRef]
- Ahmed, D.; Cassol, E. Role of cellular metabolism in regulating type I interferon responses: Implications for tumour immunology and treatment. Cancer Lett. 2017, 409, 20–29. [Google Scholar] [CrossRef]
- Platanias, L.C. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Stark, G.R.; Darnell, J.E., Jr. The JAK-STAT pathway at twenty. Immunity 2012, 36, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Man, K.; Miasari, M.; Shi, W.; Xin, A.; Henstridge, D.C.; Preston, S.; Pellegrini, M.; Belz, G.T.; Smyth, G.K.; Febbraio, M.A.; et al. The transcription factor IRF4 is essential for TCR affinity-mediated metabolic programming and clonal expansion of T cells. Nat. Immunol. 2013, 14, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Takami, M.; Yamada, A.; Wang, X.; Koga, T.; Hu, X.; Tamura, T.; Ozato, K.; Choi, Y.; Ivashkiv, L.B.; et al. Interferon regulatory factor-8 regulates bone metabolism by suppressing osteoclastogenesis. Nat. Med. 2009, 15, 1066–1071. [Google Scholar] [CrossRef]
- Sadler, A.J.; Williams, B.R. Interferon-inducible antiviral effectors. Nat. Rev. Immunol. 2008, 8, 559–568. [Google Scholar] [CrossRef]
- Choi, K.M.; Kim, J.J.; Yoo, J.; Kim, K.S.; Gu, Y.; Eom, J.; Jeong, H.; Kim, K.; Nam, K.T.; Park, Y.S.; et al. The interferon-inducible protein viperin controls cancer metabolic reprogramming to enhance cancer progression. J. Clin. Investig. 2022, 132, e157302. [Google Scholar] [CrossRef]
- Wang, J.; Xiang, D.; Dai, Z.; Zhu, J.; Du, Y.; Fu, G.; Chu, X. Unveiling the immunogenomic landscape of cholangiocarcinoma: Identifying new prognostic markers and therapeutic targets based on CCL5 expression. J. Gene Med. 2024, 26, e3630. [Google Scholar] [CrossRef]
- Lewis, M.W.; Wisniewska, K.; King, C.M.; Li, S.; Coffey, A.; Kelly, M.R.; Regner, M.J.; Franco, H.L. Enhancer RNA Transcription Is Essential for a Novel CSF1 Enhancer in Triple-Negative Breast Cancer. Cancers 2022, 14, 1852. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, F.; Wu, X.; Du, K.; Yang, Q.; Xia, T. The m6A-regulation and single cell effect pattern in sunitinib resistance on clear cell renal cell carcinoma: Identification and validation of targets. Front. Pharmacol. 2023, 14, 1131610. [Google Scholar] [CrossRef]
- Wei, C.; Zheng, C.; Sun, J.; Luo, D.; Tang, Y.; Zhang, Y.; Ke, X.; Liu, Y.; Zheng, Z.; Wang, H. Viperin Inhibits Enterovirus A71 Replication by Interacting with Viral 2C Protein. Viruses 2018, 11, 13. [Google Scholar] [CrossRef]
- Shen, G.; Wang, K.; Wang, S.; Cai, M.; Li, M.-l.; Zheng, C.; Sandri-Goldin, R.M. Herpes Simplex Virus 1 Counteracts Viperin via Its Virion Host Shutoff Protein UL41. J. Virol. 2014, 88, 12163–12166. [Google Scholar] [CrossRef] [PubMed]
- Hayderi, A.; Kumawat, A.K.; Shavva, V.S.; Dreifaldt, M.; Sigvant, B.; Petri, M.H.; Kragsterman, B.; Olofsson, P.S.; Sirsjö, A.; Ljungberg, L.U. RSAD2 is abundant in atherosclerotic plaques and promotes interferon-induced CXCR3-chemokines in human smooth muscle cells. Sci. Rep. 2024, 14, 8196. [Google Scholar] [CrossRef] [PubMed]
- Shomar, H.; Georjon, H.; Feng, Y.; Olympio, B.; Guillaume, M.; Tesson, F.; Cury, J.; Wu, F.; Bernheim, A. Viperin immunity evolved across the tree of life through serial innovations on a conserved scaffold. Nat. Ecol. Evol. 2024, 8, 1667–1679. [Google Scholar] [CrossRef]
- Kennelly, S.A.; Sawyer, J.M.; Payne, A.F.; Ciota, A.T.; Harki, D.A. Development of 3′-Deoxy-3′,4′-didehydro-nucleoside Prodrug Inhibitors of West Nile and Zika Viruses. ACS Med. Chem. Lett. 2024, 15, 1334–1339. [Google Scholar] [CrossRef]
- Ding, X.; Zhou, Y.; Qiu, X.; Xu, X.; Hu, X.; Qin, J.; Chen, Y.; Zhang, M.; Ke, J.; Liu, Z.; et al. RSAD2: A pathogenic interferon-stimulated gene at the maternal-fetal interface of patients with systemic lupus erythematosus. Cell Rep. Med. 2025, 6, 101974. [Google Scholar] [CrossRef]
- Negatu, S.G.; Jurado, K.A. Targeting pathogenic interferon-stimulated gene RSAD2 improves pregnancy outcomes in systemic lupus erythematosus models. Cell Rep. Med. 2025, 6, 102034. [Google Scholar] [CrossRef]
- Cruz, P.D.; Wargowsky, R.; Gonzalez-Almada, A.; Sifontes, E.P.; Shaykhinurov, E.; Jaatinen, K.; Jepson, T.; Lafleur, J.E.; Yamane, D.; Perkins, J.; et al. Blood RNA Biomarkers Identify Bacterial and Biofilm Coinfections in COVID-19 Intensive Care Patients. J. Intensive Care Med. 2024, 39, 1071–1082. [Google Scholar] [CrossRef]
- Li, Y.; Cui, Q.; Zhou, B.; Zhang, J.; Guo, R.; Wang, Y.; Xu, X. RSAD2, a pyroptosis-related gene, predicts the prognosis and immunotherapy response for colorectal cancer. Am. J. Cancer Res. 2024, 14, 2507–2522. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Species | Virus | Mechanism | Critical Domains | Ref. |
|---|---|---|---|---|
| Viruses of the genus Flavivirus | DENV | The carboxyl terminus of Viperin interacts with the NS3 protein of DENV-2 to reduce early RNA production by interfering with DENV-2 replication complex synthesis. | C′-terminus | [30,56] |
| ZIKV | Interfere with early viral RNA synthesis by inducing the degradation of NS3 protein in ZIKV. | C′-terminus | [53,54] | |
| TBEV | Specifically degrade the nonstructural protein NS3 of TBEV and its cofactors, thereby restricting the synthesis of viral proteins of TBEV in cells and inhibiting its replication. | SAM domain | [57] | |
| WNV | Disruption of intracellular lipid raft synthesis. | SAM domain | [19,28] | |
| HCV | Interfere with the binding of NS5A protein to the host protein hVAP-33. | C′-terminus and amphipathic helix | [61,62] | |
| HIV | Inhibits viral egress. | SAM domain | [11] | |
| BUNV | Inhibition of the Bunyamwera virus replication process. | SAM domain | [26] | |
| EIAV | Disruption of the structure of the endoplasmic reticulum inhibits the expression of capsular membrane proteins. | N-terminal hydrophobic α-helix domain | [23] | |
| RABV | Disruption of the cell membrane of cholesterol and sphingolipid. | N-terminal domain | [43] | |
| SVA | Direct interaction between the RSAD2 aa 43–70 region and the SVA 2 C protein. | N-terminal domain | [63] | |
| IAV | Inhibits viral budding from the plasma membrane, possibly via interacting with FPPS. | nd | [14] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Q.; Miao, Y.; Li, M.; Zheng, H.; Yuan, Y. Viperin: A Multifunctional Protein in Antiviral Immunity and Disease Pathogenesis. Pathogens 2025, 14, 510. https://doi.org/10.3390/pathogens14050510
Cui Q, Miao Y, Li M, Zheng H, Yuan Y. Viperin: A Multifunctional Protein in Antiviral Immunity and Disease Pathogenesis. Pathogens. 2025; 14(5):510. https://doi.org/10.3390/pathogens14050510
Chicago/Turabian StyleCui, Qun, Ying Miao, Min Li, Hui Zheng, and Yukang Yuan. 2025. "Viperin: A Multifunctional Protein in Antiviral Immunity and Disease Pathogenesis" Pathogens 14, no. 5: 510. https://doi.org/10.3390/pathogens14050510
APA StyleCui, Q., Miao, Y., Li, M., Zheng, H., & Yuan, Y. (2025). Viperin: A Multifunctional Protein in Antiviral Immunity and Disease Pathogenesis. Pathogens, 14(5), 510. https://doi.org/10.3390/pathogens14050510