
Identification of Novel Gene-Specific Markers for Differentiating Various Pathogenic Campylobacter Species Using a Pangenome Analysis Approach

 , ,
, ,  and
and
Abstract
1. Introduction
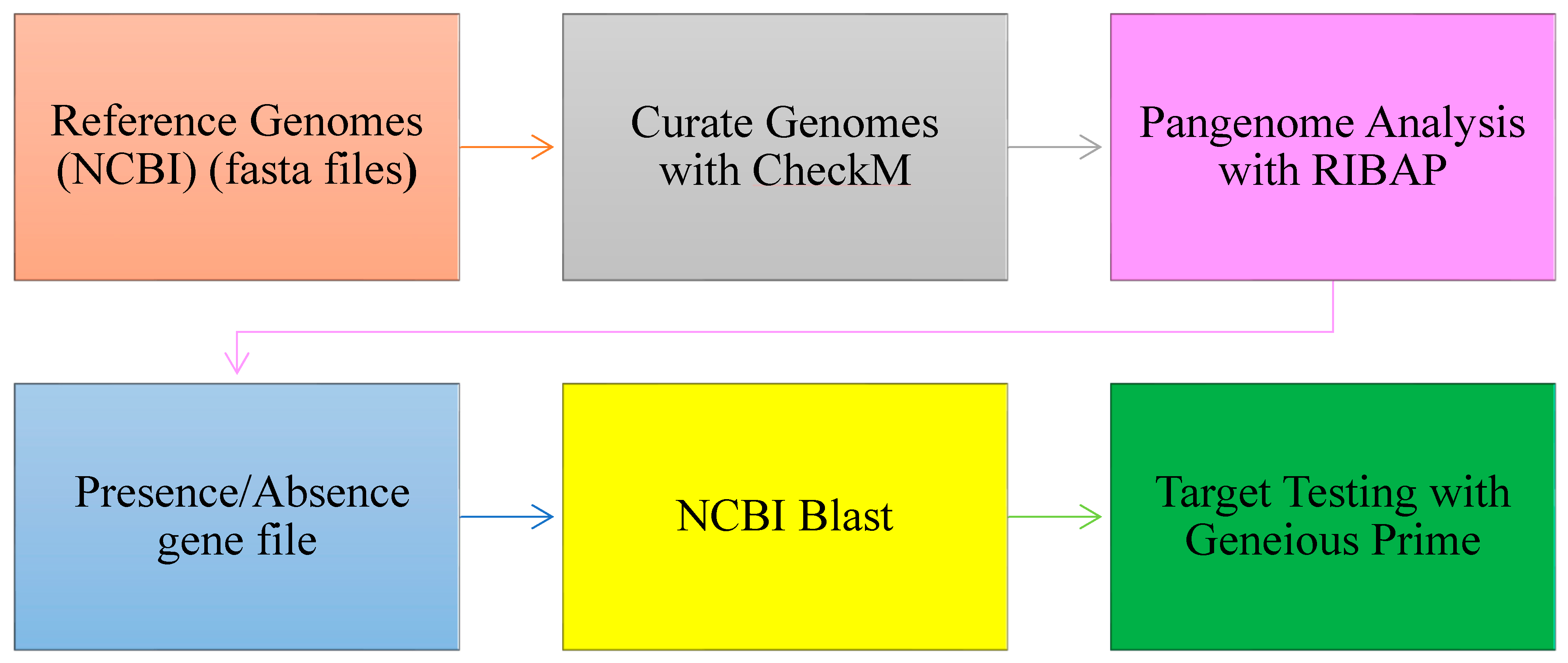
2. Materials and Methods
2.1. Data Collection
Bacterial Genomes Used for the Study
2.2. Data Analysis
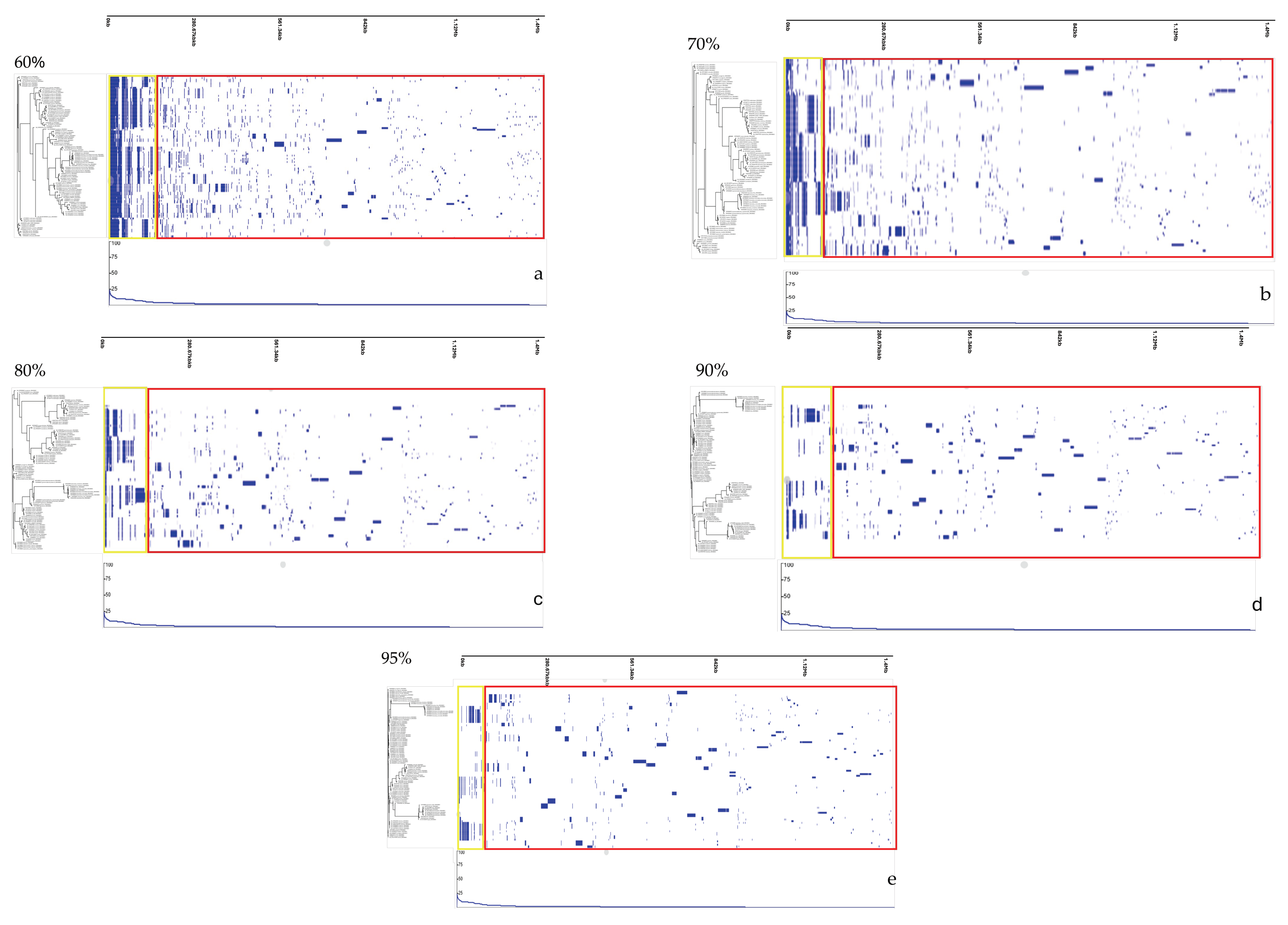
2.2.1. Pangenome Analysis to Identify Core and Accessory Genes
2.2.2. Selection of Unique Targets for Campylobacter spp. Detection
2.2.3. In Silico Validation of the Selected Targets Across Diverse Campylobacter Strains
2.2.4. Functionality of Core Genes and Identified Genetic Targets
3. Results
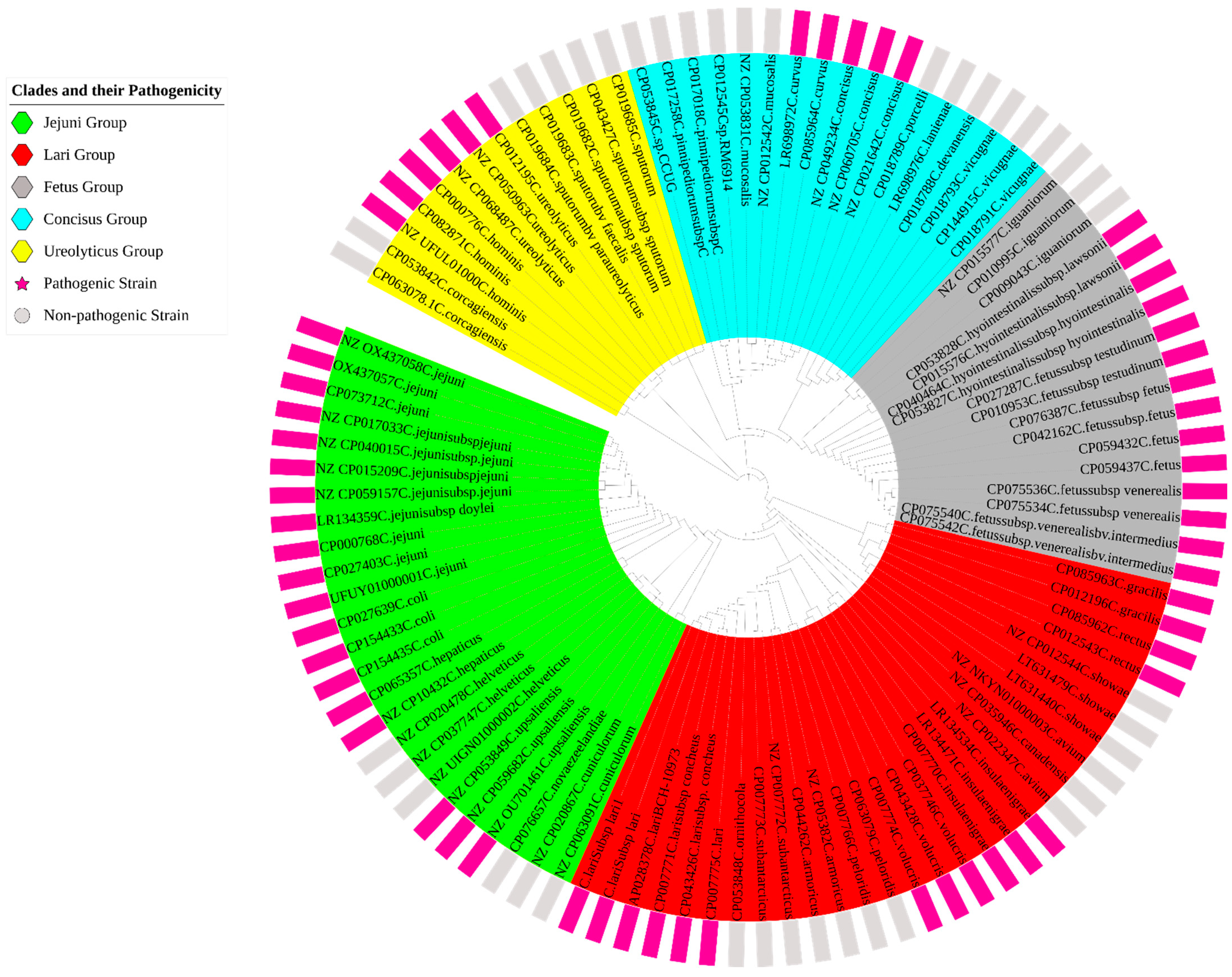
3.1. Genomic Diversity and Characteristics of Curated Strains of Campylobacter
3.2. Specific Genetic Targets and In Silico Validation (Sensitivity and Specificity)
3.3. Functionality of Core Genes and Specific Genetic Targets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laing, C.R.; Whiteside, M.D.; Gannon, V.P. Pan-genome analyses of the species Salmonella enterica, and identification of genomic markers predictive for species, subspecies, and serovar. Front. Microbiol. 2017, 8, 1345. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.; Mahon, B.; Jones, T.; Griffin, P. An assessment of the human health impact of seven leading foodborne pathogens in the United States using disability adjusted life years. Epidemiol. Infect. 2015, 143, 2795–2804. [Google Scholar] [CrossRef]
- Bergholz, T.M.; Switt, A.I.M.; Wiedmann, M. Omics approaches in food safety: Fulfilling the promise? Trends Microbiol. 2014, 22, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Tack, D.M. Preliminary incidence and trends of infections with pathogens transmitted commonly through food—Foodborne Diseases Active Surveillance Network, 10 US Sites, 2016–2019. MMWR. Morb. Mortal. Wkly. Rep. 2020, 69, 509–514. [Google Scholar] [CrossRef]
- Vidic, J.; Vizzini, P.; Manzano, M.; Kavanaugh, D.; Ramarao, N.; Zivkovic, M.; Radonic, V.; Knezevic, N.; Giouroudi, I.; Gadjanski, I. Point-of-need DNA testing for detection of foodborne pathogenic bacteria. Sensors 2019, 19, 1100. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef]
- Ford, L.; Healy, J.M.; Cui, Z.; Ahart, L.; Medalla, F.; Ray, L.C.; Reynolds, J.; Laughlin, M.E.; Vugia, D.J.; Hanna, S. Epidemiology and antimicrobial resistance of Campylobacter infections in the United States, 2005–2018. Open Forum Infect. Dis. 2023, 10, ofad378. [Google Scholar] [CrossRef]
- Tramuta, C.; Garcia-Vozmediano, A.; Group, C.C.L.; Maurella, C.; Bianchi, D.M.; Decastelli, L.; Pitti, M. Phenotypic Antimicrobial Resistance Profiles of Human Campylobacter Species Isolated in Northwest Italy, 2020–2023. Microorganisms 2024, 12, 426. [Google Scholar] [CrossRef]
- EFSA. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA J. 2017, 15, e05077. [Google Scholar]
- EFSA. The European Union One Health 2022 Zoonoses Report. EFSA J. 2023, 21, e8442. [Google Scholar]
- Christidis, T.; Pintar, K.; Butler, A.; Nesbitt, A.; Thomas, M.; Marshall, B.; Pollari, F. Campylobacter spp. prevalence and levels in raw milk: A systematic review and meta-analysis. J. Food Prot. 2016, 79, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, B.J.; Wirsing, E.; Devlin, V.; Kamhi, L.; Temple, B.; Weening, K.; Cavallo, S.; Allen, L.; Brinig, P.; Goode, B. Multistate outbreak of Campylobacter jejuni infections associated with undercooked chicken livers—Northeastern United States, 2012. Morb. Mortal. Wkly. Rep. 2013, 62, 874. [Google Scholar]
- Mohammadpour, H.; Berizi, E.; Hosseinzadeh, S.; Majlesi, M.; Zare, M. The prevalence of Campylobacter spp. in vegetables, fruits, and fresh produce: A systematic review and meta-analysis. Gut Pathog. 2018, 10, 1–12. [Google Scholar] [CrossRef]
- Verhoeff-Bakkenes, L.; Jansen, H.; In’t Veld, P.; Beumer, R.; Zwietering, M.; Van Leusden, F. Consumption of raw vegetables and fruits: A risk factor for Campylobacter infections. Int. J. Food Microbiol. 2011, 144, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Pitkänen, T. Review of Campylobacter spp. in drinking and environmental waters. J. Microbiol. Methods 2013, 95, 39–47. [Google Scholar] [CrossRef]
- Dessouky, Y.E.; Elsayed, S.W.; Abdelsalam, N.A.; Saif, N.A.; Álvarez-Ordóñez, A.; Elhadidy, M. Genomic insights into zoonotic transmission and antimicrobial resistance in Campylobacter jejuni from farm to fork: A one health perspective. Gut Pathog. 2022, 14, 44. [Google Scholar] [CrossRef]
- French, N.P.; Midwinter, A.; Holland, B.; Collins-Emerson, J.; Pattison, R.; Colles, F.; Carter, P. Molecular epidemiology of Campylobacter jejuni isolates from wild-bird fecal material in children’s playgrounds. Appl. Environ. Microbiol. 2009, 75, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Pascoe, B.; Futcher, G.; Pensar, J.; Bayliss, S.C.; Mourkas, E.; Calland, J.K.; Hitchings, M.D.; Joseph, L.A.; Lane, C.G.; Greenlee, T. Machine learning to attribute the source of Campylobacter infections in the United States: A retrospective analysis of national surveillance data. J. Infect. 2024, 89, 106265. [Google Scholar] [CrossRef]
- Wu, R.; Payne, M.; Zhang, L.; Lan, R. Uncovering the boundaries of Campylobacter species through large-scale phylogenetic and nucleotide identity analyses. Msystems 2024, 9, e01223. [Google Scholar] [CrossRef]
- Costa, D.; Iraola, G. Pathogenomics of emerging Campylobacter species. Clin. Microbiol. Rev. 2019, 32, e00072-18. [Google Scholar] [CrossRef]
- Liu, F.; Lee, S.A.; Xue, J.; Riordan, S.M.; Zhang, L. Global epidemiology of campylobacteriosis and the impact of COVID-19. Front. Cell. Infect. Microbiol. 2022, 12, 979055. [Google Scholar] [CrossRef]
- Li, T.; Wolfert, M.A.; Wei, N.; Huizinga, R.; Jacobs, B.C.; Boons, G.-J. Chemoenzymatic synthesis of Campylobacter jejuni lipo-oligosaccharide core domains to examine Guillain–Barré syndrome serum antibody specificities. J. Am. Chem. Soc. 2020, 142, 19611–19621. [Google Scholar] [CrossRef] [PubMed]
- Kalischuk, L.D.; Inglis, G.D.; Buret, A.G. Campylobacter jejuni induces transcellular translocation of commensal bacteria via lipid rafts. Gut Pathog. 2009, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, V.; Riordan, S.M.; Grimm, M.C.; Tran, T.A.T.; Major, J.; Kaakoush, N.O.; Mitchell, H.; Zhang, L. Prevalence of Campylobacter species in adult Crohn’s disease and the preferential colonization sites of Campylobacter species in the human intestine. PLoS ONE 2011, 6, e25417. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Saad, A.M.; Yang, T.; Salem, H.M.; Korma, S.A.; Ahmed, A.E.; Mosa, W.F.; Abd El-Mageed, T.A.; Selim, S.; Al Jaouni, S.K. Avian campylobacteriosis, prevalence, sources, hazards, antibiotic resistance, poultry meat contamination, and control measures: A comprehensive review. Poult. Sci. 2023, 102, 102786. [Google Scholar] [CrossRef]
- Zhong, C.; Qu, B.; Hu, G.; Ning, K. Pan-genome analysis of Campylobacter: Insights on the genomic diversity and virulence profile. Microbiol. Spectr. 2022, 10, e01022. [Google Scholar] [CrossRef]
- Sahin, O.; Yaeger, M.; Wu, Z.; Zhang, Q. Campylobacter-associated diseases in animals. Annu. Rev. Anim. Biosci. 2017, 5, 21–42. [Google Scholar] [CrossRef]
- Pena-Fernández, N.; Ocejo, M.; van der Graaf-van Bloois, L.; Lavín, J.L.; Kortabarria, N.; Collantes-Fernández, E.; Hurtado, A.; Aduriz, G. Comparative pangenomic analysis of Campylobacter fetus isolated from Spanish bulls and other mammalian species. Sci. Rep. 2024, 14, 4347. [Google Scholar] [CrossRef]
- Sprenger, H.; Zechner, E.L.; Gorkiewicz, G. So close and yet so far—Molecular microbiology of Campylobacter fetus subspecies. Eur. J. Microbiol. Immunol. 2012, 2, 66–75. [Google Scholar] [CrossRef]
- Vizzini, P.; Braidot, M.; Vidic, J.; Manzano, M. Electrochemical and optical biosensors for the detection of Campylobacter and Listeria: An update look. Micromachines 2019, 10, 500. [Google Scholar] [CrossRef]
- Ricke, S.C.; Feye, K.M.; Chaney, W.E.; Shi, Z.; Pavlidis, H.; Yang, Y. Developments in rapid detection methods for the detection of foodborne Campylobacter in the United States. Front. Microbiol. 2019, 9, 3280. [Google Scholar] [CrossRef] [PubMed]
- Vizzini, P.; Vidic, J.; Manzano, M. Enrichment Free qPCR for Rapid Identification and Quantification of Campylobacter jejuni, C. coli, C. lari, and C. upsaliensis in Chicken Meat Samples by a New Couple of Primers. Foods 2021, 10, 2341. [Google Scholar] [CrossRef] [PubMed]
- Böhme, K.; Antelo, S.C.; Fernández-No, I.; Quintela-Baluja, M.; Barros-Velázquez, J.; Cañas, B.; Calo-Mata, P. Detection of foodborne pathogens using MALDI-TOF mass spectrometry. In Antimicrobial Food Packaging; Elsevier: Amsterdam, The Netherlands, 2025; pp. 277–291. [Google Scholar]
- Guernier-Cambert, V.; Trachsel, J.; Maki, J.; Qi, J.; Sylte, M.J.; Hanafy, Z.; Kathariou, S.; Looft, T. Natural horizontal gene transfer of antimicrobial resistance genes in Campylobacter spp. from turkeys and swine. Front. Microbiol. 2021, 12, 732969. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Lamkiewicz, K.; Barf, L.-M.; Sachse, K.; Hölzer, M. RIBAP: A comprehensive bacterial core genome annotation pipeline for pangenome calculation beyond the species level. Genome Biol. 2024, 25, 170. [Google Scholar] [CrossRef]
- Di Tommaso, P.; Chatzou, M.; Floden, E.W.; Barja, P.P.; Palumbo, E.; Notredame, C. Nextflow enables reproducible computational workflows. Nat. Biotechnol. 2017, 35, 316–319. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Hadfield, J.; Croucher, N.J.; Goater, R.J.; Abudahab, K.; Aanensen, D.M.; Harris, S.R. Phandango: An interactive viewer for bacterial population genomics. Bioinformatics 2018, 34, 292–293. [Google Scholar] [CrossRef]
- Costa, D.; Lévesque, S.; Kumar, N.; Fresia, P.; Ferrés, I.; Lawley, T.D.; Iraola, G. Pangenome analysis reveals genetic isolation in Campylobacter hyointestinalis subspecies adapted to different mammalian hosts. Sci. Rep. 2021, 11, 3431. [Google Scholar] [CrossRef]
- Gemmell, M.R.; Berry, S.; Mukhopadhya, I.; Hansen, R.; Nielsen, H.L.; Bajaj-Elliott, M.; Nielsen, H.; Hold, G.L. Comparative genomics of Campylobacter concisus: Analysis of clinical strains reveals genome diversity and pathogenic potential. Emerg. Microbes Infect. 2018, 7, 116. [Google Scholar] [CrossRef]
- Costa, S.S.; Guimarães, L.C.; Silva, A.; Soares, S.C.; Baraúna, R.A. First steps in the analysis of prokaryotic pan-genomes. Bioinform. Biol. Insights 2020, 14, 1177932220938064. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.J.; Gabriel, E.; Leatherbarrow, A.J.; Cheesbrough, J.; Gee, S.; Bolton, E.; Fox, A.; Hart, C.A.; Diggle, P.J.; Fearnhead, P. Rapid evolution and the importance of recombination to the gastroenteric pathogen Campylobacter jejuni. Mol. Biol. Evol. 2009, 26, 385–397. [Google Scholar] [CrossRef]
- Rodriguez-R, L.M.; Conrad, R.E.; Viver, T.; Feistel, D.J.; Lindner, B.G.; Venter, S.N.; Orellana, L.H.; Amann, R.; Rossello-Mora, R.; Konstantinidis, K.T. An ANI gap within bacterial species that advances the definitions of intra-species units. mBio 2024, 15, e02623. [Google Scholar] [CrossRef] [PubMed]
- Quisel, J.D.; Grossman, A.D. Control of sporulation gene expression in Bacillus subtilis by the chromosome partitioning proteins Soj (ParA) and Spo0J (ParB). J. Bacteriol. 2000, 182, 3446–3451. [Google Scholar] [CrossRef]
- Zhou, Y.; Bu, L.; Guo, M.; Zhou, C.; Wang, Y.; Chen, L.; Liu, J. Comprehensive genomic characterization of Campylobacter genus reveals some underlying mechanisms for its genomic diversification. PLoS ONE 2013, 8, e70241. [Google Scholar] [CrossRef]
- Epping, L.; Walther, B.; Piro, R.M.; Knüver, M.-T.; Huber, C.; Thürmer, A.; Flieger, A.; Fruth, A.; Janecko, N.; Wieler, L.H. Genome-wide insights into population structure and host specificity of Campylobacter jejuni. Sci. Rep. 2021, 11, 10358. [Google Scholar] [CrossRef]
- Mourkas, E.; Yahara, K.; Bayliss, S.C.; Calland, J.K.; Johansson, H.; Mageiros, L.; Muñoz-Ramirez, Z.Y.; Futcher, G.; Méric, G.; Hitchings, M.D. Host ecology regulates interspecies recombination in bacteria of the genus Campylobacter. eLife 2022, 11, e73552. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Payne, M.; Lan, R. In silico identification of serovar-specific genes for Salmonella serotyping. Front. Microbiol. 2019, 10, 835. [Google Scholar] [CrossRef]
- Yang, S.-M.; Kim, E.; Kim, D.; Kim, H.-B.; Baek, J.; Ko, S.; Kim, D.; Yoon, H.; Kim, H.-Y. Rapid real-time polymerase chain reaction for Salmonella serotyping based on novel unique gene markers by pangenome analysis. Front. Microbiol. 2021, 12, 750379. [Google Scholar] [CrossRef]
- Méric, G.; Yahara, K.; Mageiros, L.; Pascoe, B.; Maiden, M.C.; Jolley, K.A.; Sheppard, S.K. A reference pan-genome approach to comparative bacterial genomics: Identification of novel epidemiological markers in pathogenic Campylobacter. PLoS ONE 2014, 9, e92798. [Google Scholar] [CrossRef]
- Iraola, G.; Perez, R.; Naya, H.; Paolicchi, F.; Pastor, E.; Valenzuela, S.; Calleros, L.; Velilla, A.; Hernández, M.; Morsella, C. Genomic evidence for the emergence and evolution of pathogenicity and niche preferences in the genus Campylobacter. Genome Biol. Evol. 2014, 6, 2392–2405. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Rodríguez, M.; Córdoba, J.J.; Andrade, M.J. Design of primers and probes for quantitative real-time PCR methods. In PCR Primer Design; Humana: New York, NY, USA, 2015; pp. 31–56. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification of Genes | 60% | 70% | 80% | 90% | 95% | |
|---|---|---|---|---|---|---|
| Core genes | 226 | 119 | 46 | 9 | 1 | |
| Soft-core genes | 77 | 49 | 23 | 3 | 1 | |
| Accessory genes | Shell genes | 2419 | 2529 | 2031 | 889 | 339 |
| Cloud genes | 22,699 | 33,788 | 43,063 | 53,319 | 69,826 | |
| Pathogenic Species/Subspecies | Protein Name (Gene) | Accession No. | Size (bp) |
|---|---|---|---|
| C. coli | Cytochrome b (PetB) ** | WP_038836333.1 | 1248 |
| C. coli | ATP-binding subunit (ClpX) ** | WP_002778039.1 | 1224 |
| C. coli | ATP-dependent Clp protease ** | WP_264378315.1 | 291 |
| C. coli | Transaldolase ** | WP_289867517 | 987 |
| C. coli | Hypothetical protein ** | WP_002779418.1 | 171 |
| C. coli | Carbamoyl-phosphate synthase large subunit (carB) ** | WP_264378315.1 | 3270 |
| C. concisus | Hypothetical protein ** | WP_072594306.1 | 432 |
| C. concisus | Hypothetical protein | WP_103643789.1 | 429 |
| C. concisus | Phosphoribosylformlyglcinsmidine cyclo-ligase | WP_107892589.1 | 984 |
| C. curvus | Hypothetical protein ** | WP_009649311.1 | 711 |
| C. curvus | Hypothetical protein ** | WP_018136234.1 | 696 |
| C. curvus | Virulence protein ** | WP_011992574.1 | 393 |
| C. curvus | Bifunctional enzyme IspD/IspF (ispDF) | WP_018136231.1 | 1116 |
| C. fetus | Hypothetical protein ** | WP_024305373.1 | 1296 |
| C. fetus | Hypothetical protein ** | WP_144685876.1 | 654 |
| C. fetus subp. testudinum | Lipopolysaccharide export system protein (lptA) | WP_023385482.1 | 468 |
| C. fetus subp. testudinum | Hypothetical protein | WP_039362567.1 | 1248 |
| C. fetus subp. testudinum | Hypothetical protein | WP_058909030.1 | 936 |
| C. fetus subsp. fetus | Type I-B CRISPR associated endonuclease Cas1 (cas1b) | WP_041738340.1 | 1002 |
| C. fetus subsp. fetus | Hypothetical protein | WP_038454040.1 | 228 |
| C. fetus subsp. veneralis | Hypothetical protein ** | WP_303297428.1 | 243 |
| C. fetus subsp. veneralis | Hypothetical protein ** | WP_303297427.1 | 297 |
| C. fetus subsp. veneralis | Hypothetical protein | WP_002850340.1 | 261 |
| C. fetus subsp. veneralis | Hypothetical protein | AIR80954.1 | 279 |
| C. gracilis | Putative oxidoreductase | WP_050346355.1 | 855 |
| C. gracilis | DNA adenine methylase ** | EEV17345.1 | 813 |
| C. gracilis | Apolipoprotein N-acyltransferase ** | WP_005872283.1 | 1182 |
| C. hepaticus | Hypothetical protein | WP_124134096.1 | 780 |
| C. hepaticus | L-asparaginase 2 (ansA) ** | MDX2324043.1 | 231 |
| C. hominis | Hypothetical protein ** | WP_012109050.1 | 711 |
| C. hominis | Hypothetical protein ** | WP_011991502.1 | 918 |
| C. hyointestinalis subsp. hyointestinalis | Arginine exporter protein (argO) | WP_232051094.1 | 603 |
| C. hyointestinalis subsp. lawsonii | Beta sliding clamp | WP_063997406.1 | 1071 |
| C. hyointestinalis subsp. lawsonii | Manganese transport system membrane protein (mtB_3) | WP_151062156.1 | 825 |
| C. hyointestinalis subsp. lawsonii | Flagellar biosynthesis protein (flhA) | WP_244948766.1 | 2160 |
| C. insulaenigrae | Altronate dehydratase (uxaA_2) ** | WP_039651237.1 | 264 |
| C. insulaenigrae | Flavin-dependent thymidylate synthase (thyX) | WP_039648794.1 | 633 |
| C. insulaenigrae | 4-hydroxy-tetrahydrodipicolinate synthase (dapA_2) | WP_039651239.1 | 909 |
| C. jejuni | Hypothetical protein ** | SUW97209.1 | 552 |
| C. jejuni | Hypothetical protein ** | MBX0540796.1 | 183 |
| C. jejuni | Methyl-accepting chemotaxis protein ** | WP_257408974.1 | 420 |
| C. jejuni subsp. doylei | Hypothetical protein | ABS44036.1 | 231 |
| C. jejuni subsp. jejuni | Hypothetical protein | ADT66919.1 | 378 |
| C. lari | Major outer membrane protein (porA_2) | WP_317728191.1 | 525 |
| C. lari subp. lari | Hypothetical protein | EFN2797389.1 | 1575 |
| C. lari subsp. concheus | Hypothetical protein ** | EAJ5700938.1 | 1353 |
| C. rectus | Hypothetical protein ** | WP_002945817.1 | 225 |
| C. upsaliensis | Spermidine export protein (mdlj) ** | WP_176318412.1 | 330 |
| C. upsaliensis | Hypothetical protein ** | WP_176318076.1 | 162 |
| C. upsaliensis | Hypothetical protein | WP_004277133.1 | 402 |
| C. upsaliensis | Hypothetical protein | EAJ0411667.1 | 666 |
| C. ureolyticus | Hypothetical protein ** | WP_202751539.1 | 891 |
| C. volucris | Peptidoglycan O-acetyltransferase (patA) | WP_149104689.1 | 1500 |
| C. volucris | Methyl-accepting chemotaxis protein (mcpA) | QEL09028.1 | 1977 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuufire, E.; Bentum, K.E.; Nyarku, R.; Osei, V.; Elrefaey, A.; James, T.; Woube, Y.; Folitse, R.; Samuel, T.; Abebe, W. Identification of Novel Gene-Specific Markers for Differentiating Various Pathogenic Campylobacter Species Using a Pangenome Analysis Approach. Pathogens 2025, 14, 477. https://doi.org/10.3390/pathogens14050477
Kuufire E, Bentum KE, Nyarku R, Osei V, Elrefaey A, James T, Woube Y, Folitse R, Samuel T, Abebe W. Identification of Novel Gene-Specific Markers for Differentiating Various Pathogenic Campylobacter Species Using a Pangenome Analysis Approach. Pathogens. 2025; 14(5):477. https://doi.org/10.3390/pathogens14050477
Chicago/Turabian StyleKuufire, Emmanuel, Kingsley E. Bentum, Rejoice Nyarku, Viona Osei, Asmaa Elrefaey, Tyric James, Yilkal Woube, Raphael Folitse, Temesgen Samuel, and Woubit Abebe. 2025. "Identification of Novel Gene-Specific Markers for Differentiating Various Pathogenic Campylobacter Species Using a Pangenome Analysis Approach" Pathogens 14, no. 5: 477. https://doi.org/10.3390/pathogens14050477
APA StyleKuufire, E., Bentum, K. E., Nyarku, R., Osei, V., Elrefaey, A., James, T., Woube, Y., Folitse, R., Samuel, T., & Abebe, W. (2025). Identification of Novel Gene-Specific Markers for Differentiating Various Pathogenic Campylobacter Species Using a Pangenome Analysis Approach. Pathogens, 14(5), 477. https://doi.org/10.3390/pathogens14050477