
Association Between IL-28B (rs8099917) and IL-28B (rs12979860) with Predisposition to Diseases Related to the HTLV-1: Systematic Review and Meta-Analysis


Abstract
1. Introduction
2. Materials and Methods
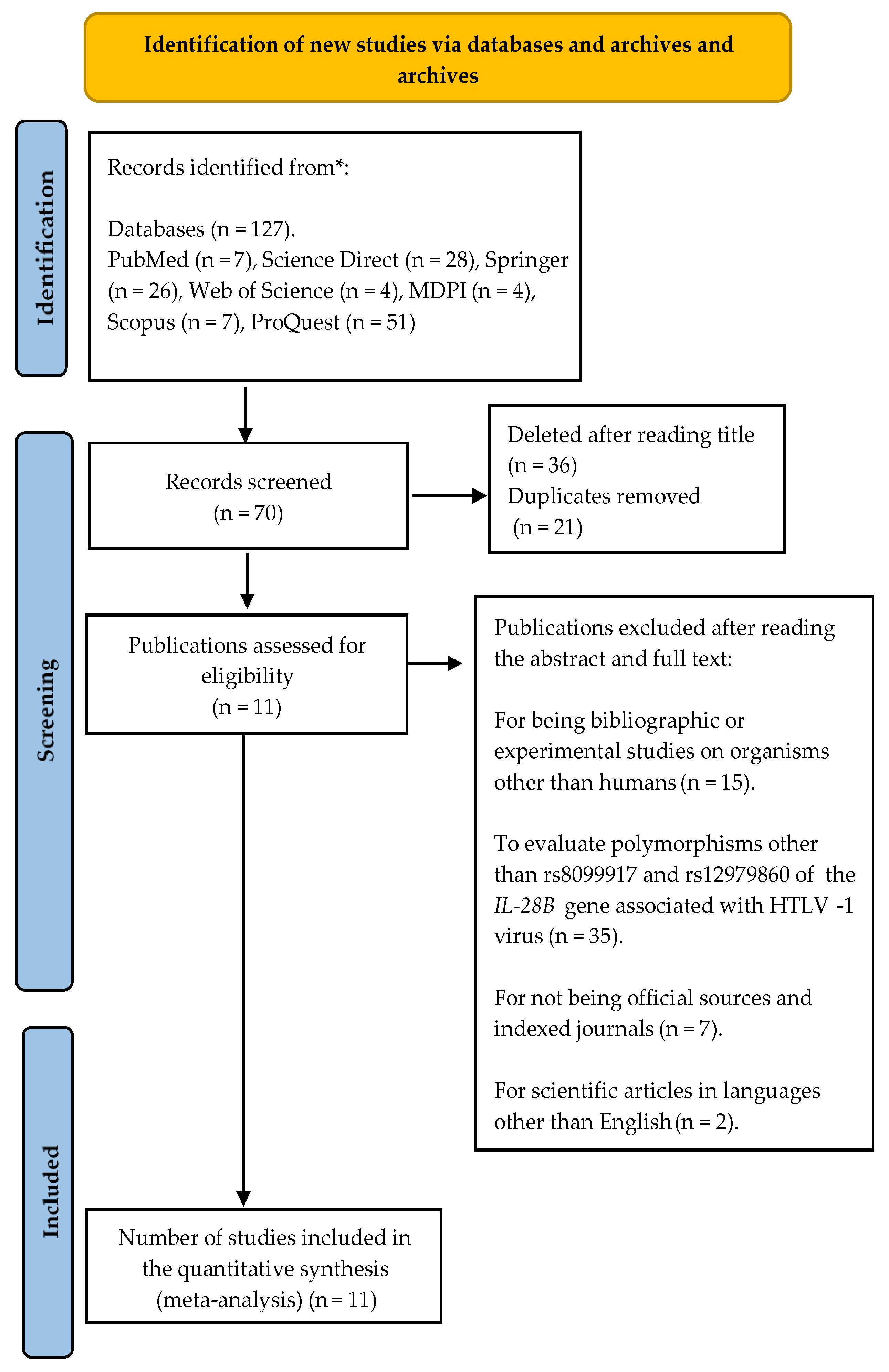
2.1. Bibliographic Search Strategy
2.2. Publication Bias
2.3. Statistical Analysis
3. Results
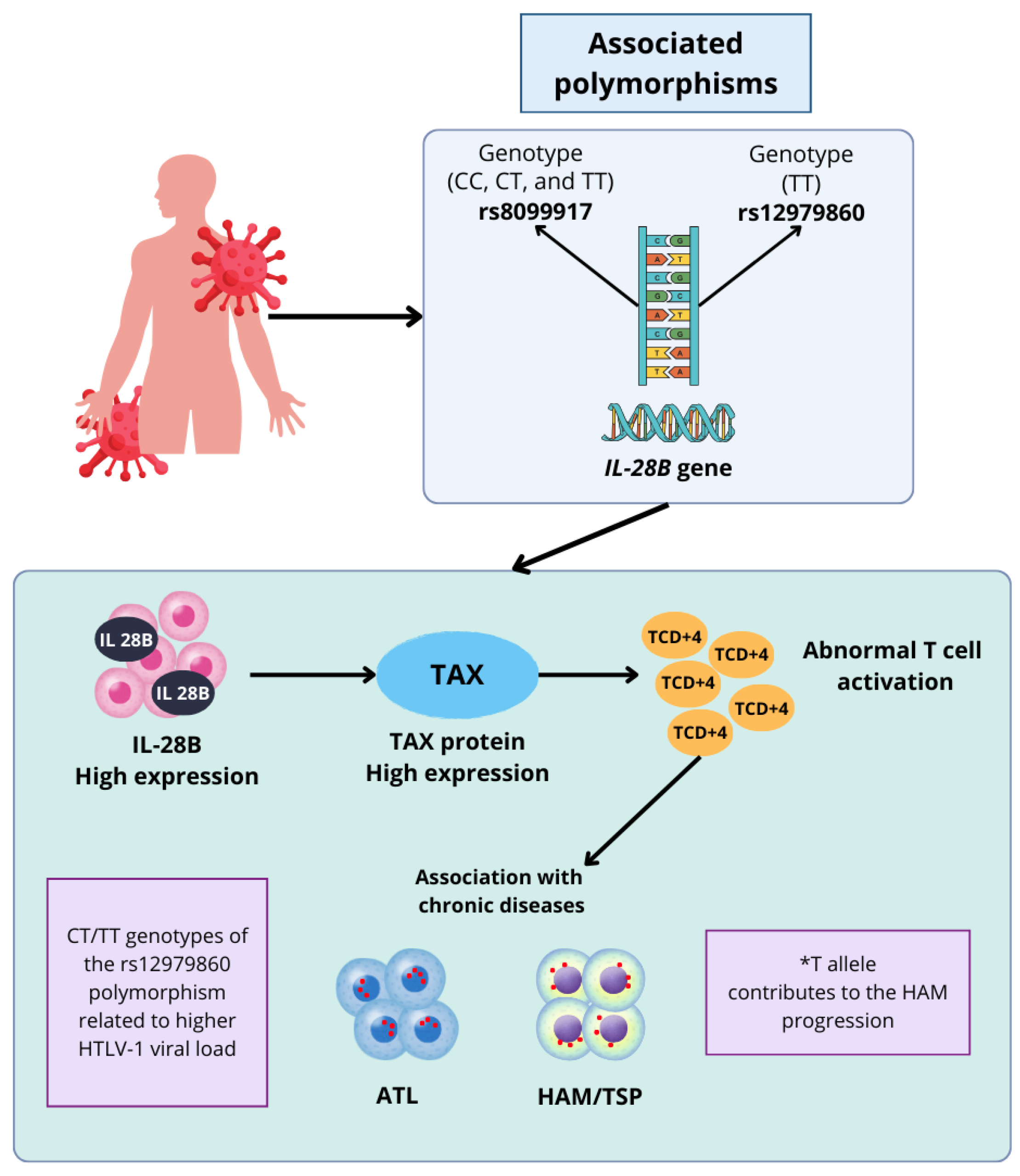
3.1. Association Between IL-28B Gene and the Pathogenesis of HTLV-1
3.2. Study Characteristics
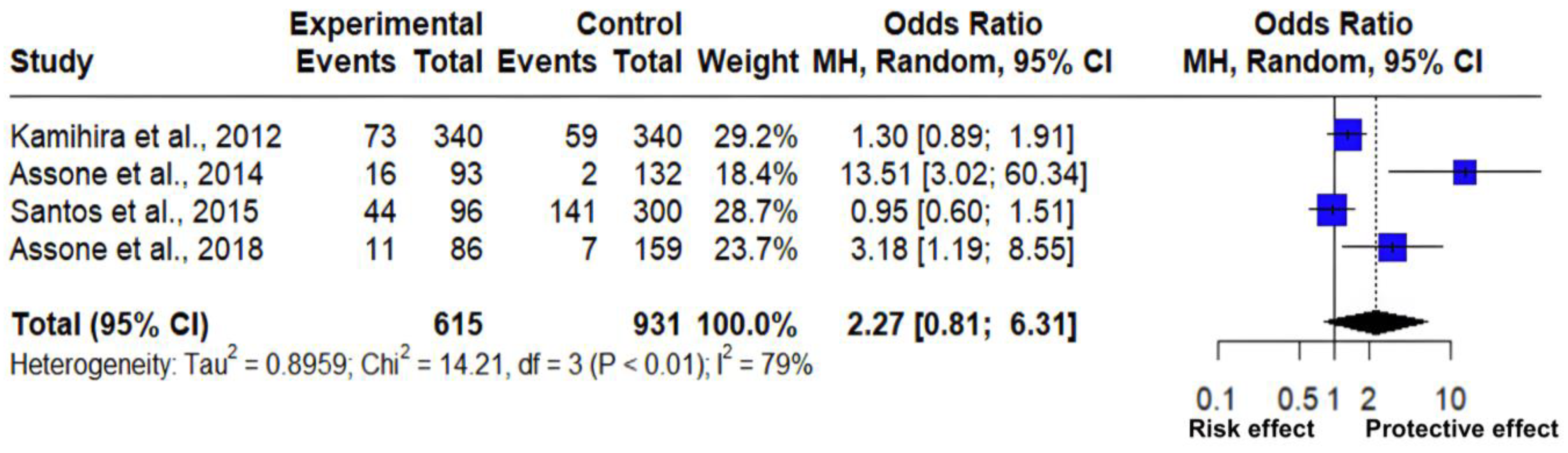
3.3. Interactions of the GG vs. TT Genotype Frequency of the IL-28B Gene rs8099917 Polymorphism
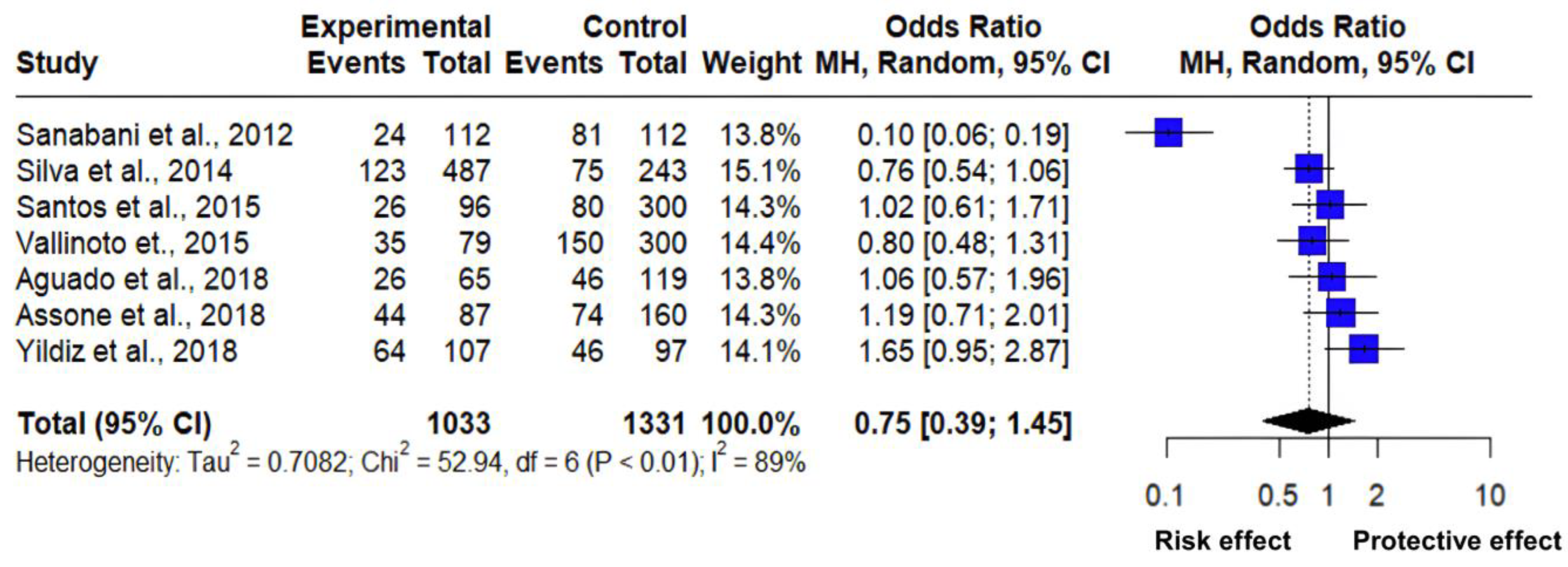
3.4. Interactions of the CC vs. CT Genotype Frequency of the IL-28B Gene rs12979860 Polymorphism
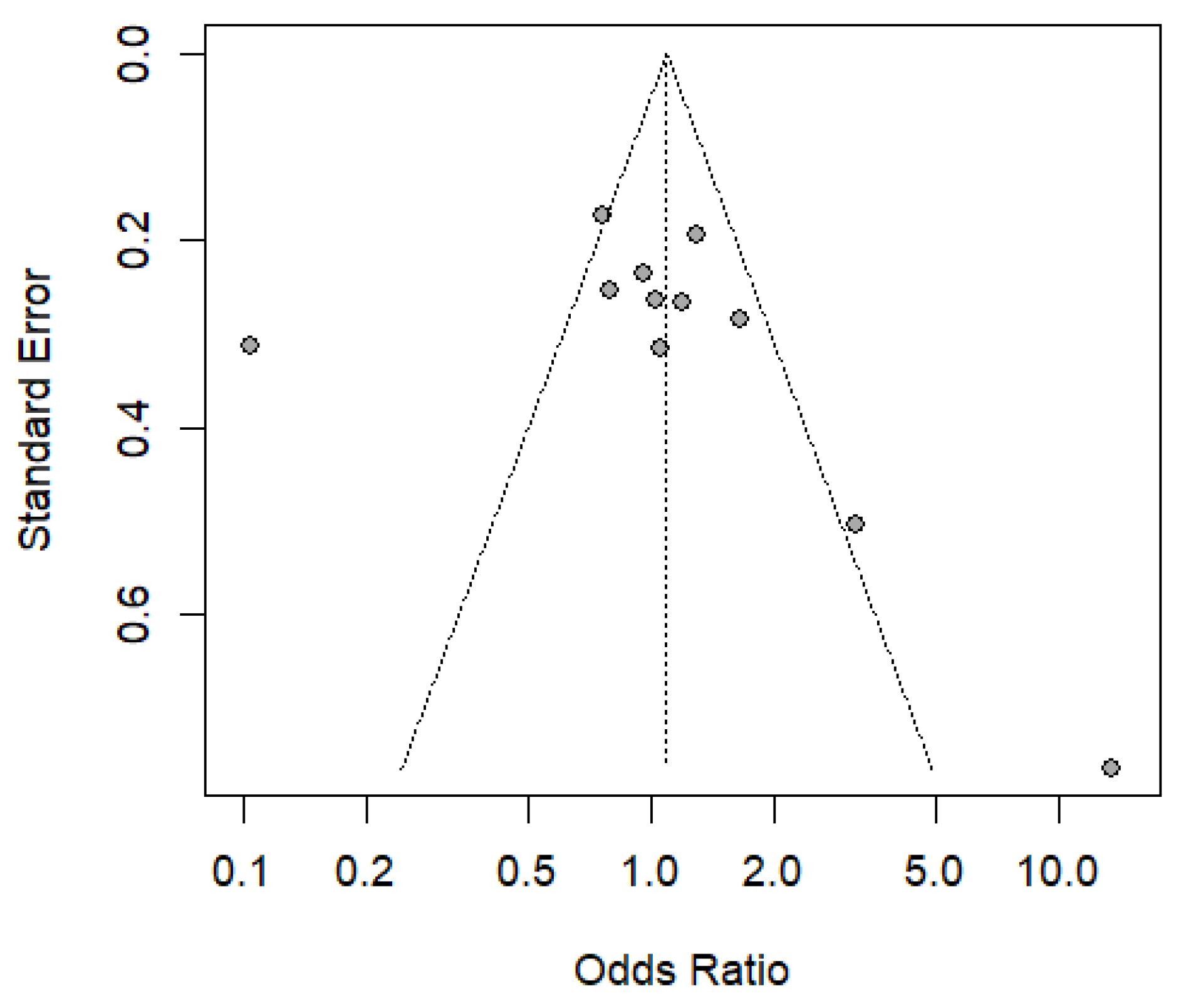
3.5. Result of the Publication Bias Analysis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Fact Sheets: Human T-Lymphotropic Virus Type 1. December 2024. Available online: https://www.who.int/news-room/fact-sheets/detail/human-t-lymphotropic-virus-type-1 (accessed on 12 March 2025).
- WHO. Public Health Impact And Implications For Future Actions: Who Global Consultation On The Human T-Lymphotropic Virus Type 1; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- WHO. Human T-Lymphotropic Virus Type 1: Technical Report February 2020; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- Organización Panamericana de la Salud. Hoja Informativa: Virus linfotrópico T humano (HTLV). 2024. Available online: https://www.paho.org/es/temas/virus-linfotropico-t-humano-htlv#info (accessed on 12 March 2025).
- Forlani, G.; Shallak, M.; Accolla, R.S.; Romanelli, M.G. Htlv-1 infection and pathogenesis: New insights from cellular and animal models. Int. J. Mol. Sci. 2021, 22, 8001. [Google Scholar] [CrossRef]
- Nakahata, S.; Enriquez-Vera, D.; Jahan, M.I.; Sugata, K.; Satou, Y. Understanding the Immunopathology of HTLV-1-Associated Adult T-Cell Leukemia/Lymphoma: A Comprehensive Review. Biomolecules 2023, 13, 1543. [Google Scholar] [CrossRef]
- Yamagishi, M.; Suzuki, Y.; Watanabe, T.; Uchimaru, K. Clonal Selection and Evolution of HTLV-1-Infected Cells Driven by Genetic and Epigenetic Alteration. Viruses 2022, 14, 587. [Google Scholar] [CrossRef]
- Yamagishi, M.; Kuze, Y.; Kobayashi, S.; Nakashima, M.; Morishima, S.; Kawamata, T.; Makiyama, J.; Suzuki, K.; Seki, M.; Abe, K.; et al. Mechanisms of action and resistance in histone methylation-targeted therapy. Nature 2024, 627, 221–228. [Google Scholar] [CrossRef]
- Toyoda, K.; Yasunaga, J.-I.; Shichijo, T.; Arima, Y.; Tsujita, K.; Tanaka, A.; Salah, T.; Zhang, W.; Hussein, O.; Sonoda, M.; et al. HTLV-1 bZIP Factor-Induced Reprogramming of Lactate Metabolism and Epigenetic Status Promote Leukemic Cell Expansion. Blood Cancer Discov. 2023, 4, 374–393. [Google Scholar] [CrossRef]
- Schwalb, A.; Pérez-Muto, V.; Cachay, R.; Tipismana, M.; Álvarez, C.; Mejía, F.; González-Lagos, E.; Gotuzzo, E. Early-onset HTLV-1-associated myelopathy/tropical spastic paraparesis. Pathogens 2020, 9, 450. [Google Scholar] [CrossRef]
- Kalinichenko, S.; Komkov, D.; Mazurov, D. HIV-1 and HTLV-1 Transmission Modes: Mechanisms and Importance for Virus Spread. Viruses 2022, 14, 152. [Google Scholar] [CrossRef]
- Yamada, A.; Yasunaga, J.; Liang, L.; Zhang, W.; Sunagawa, J.; Nakaoka, S.; Iwami, S.; Kogure, Y.; Ito, Y.; Kataoka, K.; et al. Anti-HTLV-1 immunity combined with proviral load as predictive biomarkers for adult T-cell leukemia-lymphoma. Cancer Sci. 2023, 115, 310–320. [Google Scholar] [CrossRef]
- Vallinoto, A.C.R.; Cayres-Vallinoto, I.; Queiroz, M.A.F.; Ishak, M.d.O.G.; Ishak, R. Influence of immunogenetic biomarkers in the clinical outcome of HTLV-1 infected persons. Viruses 2019, 11, 974. [Google Scholar] [CrossRef]
- Zheng, K.; Zhang, A.M. Correlations between genetic polymorphism of IFN-λ family gene and HBV infection, virus replication and clearance. Chin. Acad. Sci. 2022, 38, 893–902. [Google Scholar] [CrossRef]
- Ying, S.Y.; Hu, Y.R.; Gao, G.S.; Lou, K.H.; Huang, Z. Interleukin-28B Polymorphisms Predict the Efficacy of Peginterferon Alpha in Patients With Chronic Hepatitis B: A Meta-Analysis. Front. Med. 2021, 8, 691365. [Google Scholar] [CrossRef] [PubMed]
- Zaki, S.M.; Ahmed, H.S.; Yousif, M.M.; Awad, E.M. Interleukin 28B Polymorphism as a Predictor of Sustained Virological Response to Sofosbuvir-Based Therapy for Hepatitis C Virus Patients. Trop. Med. Infect. Dis. 2022, 7, 230. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Gao, J.; Guo, J.; Yang, D.; Li, D. Interleukin-28B dampens protease-induced lung inflammation via IL-25 and TSLP inhibition in epithelial cells. Sci. Rep. 2020, 10, 20973. [Google Scholar] [CrossRef]
- Miklos, A.C.; Li, C.; Sorrell, C.D.; Lyon, L.A.; Pielak, G.J. An upper limit for macromolecular crowding effects. BMC Biophys. 2011, 4, 13. [Google Scholar] [CrossRef]
- de Sá, K.S.G.; Santana, B.B.; Ferreira, T.C.d.S.; Sousa, R.C.M.; Caldas, C.A.M.; Azevedo, V.N.; Feitosa, R.N.M.; Machado, L.F.A.; Ishak, M.d.O.G.; Ishak, R.; et al. IL28B gene polymorphisms and Th1/Th2 cytokine levels might be associated with HTLV-associated arthropathy. Cytokine 2016, 77, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, 71. [Google Scholar] [CrossRef]
- Sos, V.J.E.; Abella, J.A.L.; Villach, L.G.; Oliver, M.B. Meta-Analysis: A Basic Way to Understand and Interpret Your Evidence. Rev. Senol. Patol. Mamar. 2021, 34, 44–51. [Google Scholar] [CrossRef]
- Bae, J.M. A suggestion for quality assessment in systematic reviews of observational studies in nutritional epidemiology. Epidemiol. Health 2016, 38, e2016014. [Google Scholar] [CrossRef]
- Nakagawa, S.; Lagisz, M.; Jennions, M.D.; Koricheva, J.; Noble, D.W.A.; Parker, T.H.; Sánchez-Tójar, A.; Yang, Y.; O’Dea, R.E. Methods for testing publication bias in ecological and evolutionary meta-analyses. Methods Ecol. Evol. 2022, 13, 4–21. [Google Scholar] [CrossRef]
- Simmonds, M. Quantifying the risk of error when interpreting funnel plots. Syst. Rev. 2015, 4, 24. [Google Scholar] [CrossRef]
- Mehar, R.; Swarnakar, S.; Lakkakula, S.; Verma, H.K.; Bhaskar, L. Interleukin-6 gene -174G>C promoter polymorphism reduces the risk of periodontitis in Brazilian populations: A meta-analysis. J. Oral Biosci. 2021, 63, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Kamihira, S.; Usui, T.; Ichikawa, T.; Uno, N.; Morinaga, Y.; Mori, S.; Nagai, K.; Sasaki, D.; Hasegawa, H.; Yanagihara, K.; et al. Paradoxical expression of IL-28B mRNA in peripheral blood in human T-cell leukemia virus Type-1 mono-infection and co-infection with hepatitis C Virus. Virol. J. 2012, 9, 40. [Google Scholar] [CrossRef]
- Assone, T.; de Souza, F.V.; Gaester, K.O.; Fonseca, L.A.M.; Luiz, O.D.C.; Malta, F.; Pinho, J.R.R.; Gonçalves, F.d.T.; Duarte, A.J.d.S.; de Oliveira, A.C.P.; et al. IL28B Gene Polymorphism SNP rs8099917 Genotype GG Is Associated with HTLV-1-Associated Myelopathy/Tropical Spastic Paraparesis (HAM/TSP) in HTLV-1 Carriers. PLoS Negl. Trop. Dis. 2014, 8, e3199. [Google Scholar] [CrossRef] [PubMed]
- Assone, T.; Malta, F.M.; Bakkour, S.; Montalvo, L.; Paiva, A.M.; Smid, J.; de Oliveira, A.C.P.; Gonçalves, F.d.T.; Luiz, O.D.C.; Fonseca, L.A.M.; et al. Polymorphisms in HLA-C and KIR alleles are not associated with HAM/TSP risk in HTLV-1-infected subjects. Virus Res. 2018, 244, 71–74. [Google Scholar] [CrossRef]
- Sanabani, S.S.; Nukui, Y.; Pereira, J.; da Costa, A.C.; de Oliveira, A.C.S.; Pessôa, R.; Leal, F.E.; Segurado, A.C.; Kallas, E.G.; Sabino, E.C. Lack of evidence to support the association of a single IL28B genotype SNP rs12979860 with the HTLV-1 clinical outcomes and proviral load. BMC Infect. Dis. 2012, 12, 374. [Google Scholar] [CrossRef]
- Vasconcelos, L.R.S.; Moura, P.; Carmo, R.F.D.; Cavalcanti, M.D.S.d.M.; Aroucha, D.C.B.L.; Dutra, R.A.; Pereira, L.M.M.B. Low IL10 serum levels as key factor for predicting the sustained virological response to IFNα/ribavirin in Brazilian patients with HCV carrying IL28B CT/TT genotype. Hum. Immunol. 2014, 75, 895–900. [Google Scholar] [CrossRef]
- Vallinoto, A.C.R.; Santana, B.B.; Sá, K.S.G.; Ferreira, T.C.S.; Sousa, R.C.M.; Azevedo, V.N.; Feitosa, R.N.M.; Machado, L.F.A.; Ishak, M.O.G.; Ishak, R. HTLV-1-Associated Myelopathy/Tropical Spastic Paraparesis Is Not Associated with SNP rs12979860 of the IL-28B Gene. Mediat. Inflamm. 2015, 2015, 804167. [Google Scholar] [CrossRef]
- Aguado, R.; Páez-Vega, A.; Agüera, M.L.; Montejo, M.; Guirado, L.; Fortún, J.; Suárez-Benjumea, A.; Len, O.; Fariñas, M.C.; de Gracia, C.; et al. Lack of evidence of association between IFNG and IL28B polymorphisms and QuantiFERON-CMV test results in seropositive transplant patients. Hum. Immunol. 2018, 79, 499–505. [Google Scholar] [CrossRef]
- Aytekin, F.Y.; Barut, H.Ş.; Rüstemoğlu, A.; Atay, A.; Günal, Ö.; Duygu, F. Factors related to fatalities and clinical progression of Crimean-Congo hemorrhagic fever patients and the effects of IL 28-B gene polymorphism. Arch. Virol. 2019, 164, 547–557. [Google Scholar] [CrossRef]
- Yamauchi, J.; Araya, N.; Yagishita, N.; Sato, T.; Yamano, Y. An update on human T-cell leukemia virus type I (HTLV-1)-associated myelopathy/tropical spastic paraparesis (HAM/TSP) focusing on clinical and laboratory biomarkers. Pharmacol. Ther. 2021, 218, 107669. [Google Scholar] [CrossRef]
- Schor, D.; Porto, L.C.; Roma, E.H.; Quintana, M.d.S.B.; Fabricio-Silva, G.M.; Bonecini-Almeida, M.G.; Araújo, A.Q.-C.; Andrada-Serpa, M.J. Lack of association between single-nucleotide polymorphisms of pro- and anti-inflammatory cytokines and HTLV-1-associated myelopathy / tropical spastic paraparesis development in patients from Rio de Janeiro, Brazil. BMC Infect. Dis. 2018, 18, 593. [Google Scholar] [CrossRef] [PubMed]
- Wagatsuma, V.M.D.; Cilião-Alves, D.C.; Rocha–Junior, M.C.; Haddad, R.; Takayanagui, O.M.; A Donadi, E.; Covas, D.T.; Kashima, S. Association of interleukin—18 gene polymorphism with susceptibility or protective effect to HTLV-1 infection. Retrovirology 2011, 8, A118. [Google Scholar] [CrossRef]
- Shirdel, A.; Azarpazhooh, M.R.; Sahebari, M.; Ghanbari, M.; Mirfeizi, S.Z.; Hutchinson, I.; Ziaee, A.; Rafatpanah, H. Association of IL-10 Gene Polymorphisms and Human T Lymphotropic Virus Type I-Associated Myelopathy/tropical Spastic Paraparesis in North-East of Iran (Mashhad). Iran J. Basic Med. Sci. 2013, 16, 258–263. [Google Scholar] [PubMed] [PubMed Central]
- Takahashi, T. Interleukin 28B genetic polymorphism and hepatitis B virus infection. World J. Gastroenterol. 2014, 20, 12026–12030. [Google Scholar] [CrossRef] [PubMed]
- Moudi, B.; Heidari, Z.; Mahmoudzadeh-Sagheb, H. Impact of host gene polymorphisms on susceptibility to chronic hepatitis B virus infection. Infect. Genet. Evol. 2016, 44, 94–105. [Google Scholar] [CrossRef]
- Elsheredy, A.G.; Ghazy, A.A.-K.; Madkour, M.A.; El Sheredy, A.G. Study of Interleukin IL-28B rs12979860 and rs8099917 Polymorphisms among HBV Infected Egyptian Patients. Egypt J. Med. Microbiol. 2017, 26, 45–51. [Google Scholar]
- Jeannin, S.; Costa, J.-M.; Poveda, J.-D.; Belrose, G.; Lezin, A.; Cabie, A.; Cesaire, R.; Olindo, S. Absence of association of IFNL3/IL28B rs 12979860 and IFNL4 ss 469415590 polymorphisms with the neurological status of HTLV-1 Afro-Caribbean subjects in Martinique. Retrovirology 2015, 12, P95. [Google Scholar] [CrossRef]
- Treviño, A.; Lopez, M.; Vispo, E.; Aguilera, A.; Ramos, J.M.; Benito, R.; Roc, L.; Eiros, J.M.; de Mendoza, C.; Soriano, V.; et al. Development of tropical spastic paraparesis in human T-lymphotropic virus type 1 carriers is influenced by interleukin 28B gene polymorphisms. Clin. Infect. Dis. 2012, 55, e1–e4. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Selection | Comparability | Results | Total |
|---|---|---|---|---|
| Kamihira et al., 2012 [27] | 3 | 2 | 3 | 8 |
| Assone et al., 2014 [28] | 3 | 1 | 2 | 6 |
| Santos et al., 2015 [19] | 4 | 2 | 3 | 9 |
| Assone et al., 2018 [29] | 2 | 1 | 2 | 5 |
| Sanabani et al., 2012 [30] | 4 | 2 | 3 | 9 |
| Silva et al., 2014 [31] | 4 | 2 | 3 | 9 |
| Vallinoto et al., 2015 [32] | 4 | 2 | 3 | 9 |
| Aguado et al., 2018 [33] | 3 | 1 | 2 | 6 |
| Yildiz et al., 2018 [34] | 3 | 2 | 3 | 8 |
| Pathology | Country | Genetic Model | Cases (n) | Total (n) | Control (n) | Total (n) | Reference |
|---|---|---|---|---|---|---|---|
| HCV | Japan | TT | 73 | 340 | 59 | 340 | [27] |
| HAM/TSP | Brazil | GG | 16 | 93 | 2 | 132 | [28] |
| Arthropathy | Brazil | TT | 44 | 96 | 141 | 300 | [19] |
| HAM/TSP | Brazil | GG | 11 | 86 | 7 | 159 | [29] |
| Z | Tau | Tau2 | I2 | H | Q | p |
|---|---|---|---|---|---|---|
| 1.57 | 0.947 | 0.8959 | 79% | 2.18 | 14.209 | 0.003 |
| Pathology | Country | Genetic Model | Cases (n) | Total (n) | Control (n) | Total (n) | Reference |
|---|---|---|---|---|---|---|---|
| HAM/TSP | Brazil | CT | 24 | 112 | 81 | 112 | [30] |
| HCV | Brazil | CC | 123 | 487 | 75 | 243 | [31] |
| Arthropathy | Brazil | CC | 26 | 96 | 80 | 300 | [19] |
| HAM/TSP | Brazil | CT | 35 | 79 | 150 | 300 | [32] |
| Cytomegalovirus | Spain | CT | 26 | 65 | 46 | 119 | [33] |
| HAM/TSP | Brazil | CT | 44 | 87 | 74 | 160 | [29] |
| Crimean–Congo hemorrhagic fever | Turkey | CT | 64 | 107 | 46 | 97 | [34] |
| Z | Tau | Tau2 | I2 | H | Q | p |
|---|---|---|---|---|---|---|
| −0.843 | 0.842 | 0.7082 | 88.7% | 2.97 | 52.94 | 0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuenca, N.; Cordero, D.; López-Ulloa, B. Association Between IL-28B (rs8099917) and IL-28B (rs12979860) with Predisposition to Diseases Related to the HTLV-1: Systematic Review and Meta-Analysis. Pathogens 2025, 14, 470. https://doi.org/10.3390/pathogens14050470
Cuenca N, Cordero D, López-Ulloa B. Association Between IL-28B (rs8099917) and IL-28B (rs12979860) with Predisposition to Diseases Related to the HTLV-1: Systematic Review and Meta-Analysis. Pathogens. 2025; 14(5):470. https://doi.org/10.3390/pathogens14050470
Chicago/Turabian StyleCuenca, Naomi, Damarys Cordero, and Brenda López-Ulloa. 2025. "Association Between IL-28B (rs8099917) and IL-28B (rs12979860) with Predisposition to Diseases Related to the HTLV-1: Systematic Review and Meta-Analysis" Pathogens 14, no. 5: 470. https://doi.org/10.3390/pathogens14050470
APA StyleCuenca, N., Cordero, D., & López-Ulloa, B. (2025). Association Between IL-28B (rs8099917) and IL-28B (rs12979860) with Predisposition to Diseases Related to the HTLV-1: Systematic Review and Meta-Analysis. Pathogens, 14(5), 470. https://doi.org/10.3390/pathogens14050470